
Abstract
Background:
In Parkinson’s disease (PD), neurodegeneration of dopaminergic systems leads to motor and non-motor abnormalities. Sex might influence the clinical PD phenotypes and progression. Previous molecular imaging data focused only on the nigro-striato-cortical dopamine system that appeared more preserved in women. There is still a lack of evidence on gender/sex differences in the mesolimbic dopaminergic system. We aimed at assessing PD gender differences in both the dopaminergic pathways, by using a brain metabolic connectivity approach. This is based on the evidence of a significant coupling between the neurotransmission and metabolic impairments.
Methods:
We included 34 idiopathic PD patients (Female/Male: 16/18) and 34 healthy controls for comparison. The molecular architecture of both the dopaminergic networks was estimated throughout partial correlation analyses using brain metabolism data obtained by fluorine-18-fluorodeoxyglucose positron emission tomography (threshold set at p < 0.01, corrected for Bonferroni multiple comparisons).
Results:
Male patients were characterized by a widespread altered connectivity in the nigro-striato-cortical network and a sparing of the mesolimbic pathway. On the contrary, PD females showed a severe altered connectivity in the mesolimbic network and only a partial reconfiguration of the nigro-striato-cortical network.
Discussion:
Our findings add remarkable knowledge on the neurobiology of gender differences in PD, with the identification of specific neural vulnerabilities. The gender differences here revealed might be due to the combination of both biological and sociodemographic life factors. Gender differences in PD should be considered also for treatments and the targeting of modifiable risk factors.
Impact statement
Multivariate methods to fluorine-18-fluorodeoxyglucose positron emission tomography (FDG-PET) brain data enable to reconstruct brain connectivity patterns within and between neurotransmitter systems, thus providing an in vivo window to the functional architecture of these systems. We focused on the dopaminergic nigro-striato-cortical and mesolimbic networks to investigate gender differences in Parkinson's disease. We found specific neural vulnerabilities of the dopaminergic pathways in males and females by using a metabolic connectivity approach. Our study contributes to the knowledge of the biological basis of gender differences in PD, which may lead to different clinical phenotypes. Understanding how gender may influence pathology is critical for prevention and treatment strategies.
Introduction
Parkinson's disease (PD) is the second-most common neurodegenerative disorder after Alzheimer's disease (AD) (Poewe et al., 2017). It is clinically defined by the combination of rigidity, bradykinesia, and rest tremor (McCann et al., 2014). The pathologic hallmark of PD is represented by the Lewy bodies neuronal inclusion, consisting largely of α-synuclein protein aggregations (McCann et al., 2014). The inclusions lead to classical neurodegeneration prominent in the substantia nigra pars compacta (SNc), which is the main cause of dopaminergic depletion within basal ganglia leading to alteration of movements.
Moreover, postmortem studies revealed variable levels of dopaminergic neural loss simultaneously in the ventral tegmental area (VTA) (Surmeier and Sulzer, 2013). VTA dopaminergic neurons project to the nucleus accumbens, amygdala, hippocampus, and prefrontal cortex, shaping the mesolimbic dopamine system (Barth et al., 2015). A series of nonmotor features, such as depression or anxiety, often occur in PD and may precede motor disturbances. In particular, these psychopathological states can be associated with damage in the dopaminergic mesolimbic system (Castrioto et al., 2016; Gustafsson et al., 2015).
PD has been found predominantly in males than in females by an approximate ratio of 2:1. Male sex, therefore, is reported as a risk factor for PD (Baldereschi et al., 2000). The expression of clinical symptoms could be influenced by gender (Haaxma et al., 2007), whose differences have been observed in PD clinical phenotype, especially in the onset of symptoms, type of motor and nonmotor symptoms, medication use, levodopa bioavailability, and quality of life (Cerri et al., 2019). Moreover, several data reported a later disease onset and a more benign motor phenotype in PD women compared with men (Haaxma et al., 2007). However, as the disease progresses, women show higher risk in developing levodopa-related motor complications (e.g., dyskinesia) compared with men, regardless of body weight. This is likely because women have higher levodopa bioavailability and lower levodopa clearance levels compared with men (Bjornestad et al., 2016). In particular, the female sex is also positively associated with the severity and prevalence of psychiatric symptoms, namely fatigue, feeling of nervousness, pain, anxiety, and depression (Martinez-Martin et al., 2012), whereas impulse control disorders are more common in PD males (Cerri et al., 2019).
Molecular neuroimaging studies have demonstrated a more preserved dopamine transporter (DAT) uptake binding in the striatal and extrastriatal regions, more in women than in men (Eusebio et al., 2012; Kaasinen et al., 2015; Koch et al., 2014). However, there is a lack of studies regarding gender differences in DAT binding in the mesolimbic dopaminergic system, which may be related to the presence of neuropsychiatric symptoms.
Studies exploring in vivo the gender influence on neurotransmitter circuits in PD have the potential to contribute to the optimization of individual treatments and interventions. Fluorine-18-fluorodeoxyglucose positron emission tomography (FDG-PET) represents a proxy for neuronal activity and an index of synaptic function and density and it is used to measure resting-state cerebral metabolic rates of glucose (Perani et al., 2017). The application of multivariate methods to FDG-PET data can reliably evaluate crucial cerebral features, such as metabolic network connectivity and the changes in neurodegenerative diseases, including synucleinopathies (Sala and Perani 2019).
In particular, brain connectivity is affected by multiple pathological events, including altered neurotransmission (Sala and Perani 2019). Mapping changes in the neurotransmission pathways via estimation of metabolic connectivity within selected brain regions is based on the assumption that energy consumption is influenced by multiple pathological events (Iaccarino et al., 2017; Perani 2014; Sala and Perani 2019).
Previous evidence revealed a significant coupling between neurotransmission impairment and integrity of metabolic networks (Holtbernd et al., 2015; Huber et al., 2020; Ko et al., 2017; Massa et al., 2019; Niethammer et al., 2013). Specifically, in PD and dementia with Lewy bodies (DLB), FDG-PET-based metabolic network expression and dopamine availability were found to provide consistent information underlying the disease process (Huber et al., 2020; Ko et al., 2017; Massa et al., 2019; Niethammer et al., 2013). Moreover, the accurate selection of brain regions of interest (ROIs), guided by the general evidence for biochemical and histochemical studies together with anatomopathological findings, ensures a reliable estimation of metabolic connectivity alterations in the neurotransmission pathways of interest (Caminiti et al., 2017, 2017; Sala et al., 2017).
In this study, we used the brain metabolic connectivity approach to investigate gender differences in the nigro-striato-cortical and mesolimbic dopaminergic networks in a cohort of idiopathic PD patients. Following previous evidence in literature and the estrogen-induced neuroprotection, we can expect two scenarios: (i) a more severe metabolic connectivity alteration in the nigro-striato-cortical system in PD males; and (ii) greater connectivity alterations in the female mesolimbic system due to the high frequency and severity of neuropsychiatric symptoms in PD females (Martinez-Martin et al., 2012).
Materials and Methods
Participants
Idiopathic PD patients
Thirty-four PD patients (mean age in years ± standard deviation [SD]: 62.97 ± 10.48; gender [F/M]: 16/18) without cognitive impairment at baseline were retrospectively collected from the clinical and imaging database of the Neurology Unit, Department of Clinical and Experimental Sciences, at the University of Brescia, Brescia, Italy. The clinical diagnosis of PD was made according to the Movement Disorder Society Clinical Diagnostic Criteria (Postuma et al., 2015). All patients underwent magnetic resonance imaging or computed tomography (CT) to exclude cerebrovascular diseases or brain/iron accumulation. Patients with deep brain stimulation-verified genetic mutation known to cause PD, concomitant psychiatric or other neurological disorders, hallucination, psychosis or antipsychotic drug use, a background of drug or alcohol addiction, and mild cognitive impairment were excluded.
Gender groups in the PD cohorts were compared through a one-way analysis of variance (Table 1). SPSS 21.0 software for Windows (SPSS, Chicago, IL) was used to perform statistical analyses.
Demographic and Clinical Characteristics
The differences were tested using a one-way analysis of variance.
M, male; F, female; SD, standard deviation; MMSE, Mini-Mental State Examination; UPDRS, Unified Parkinson's Disease Rating Scale; LED, levodopa equivalent dose; PD, Parkinson's disease.
Healthy controls
A group of 34 healthy age-matched volunteers (mean age in years ± SD: 62.52 ± 4.52; gender [F/M]: 16/18) were retrospectively collected from the internal database of the in vivo Human Molecular and Structural Neuroimaging Unit, San Raffaele Scientific Institute for Research and Healthcare (IRCCS), Milan, Italy. The subjects presented a negative medical history for neurological or other chronic diseases and they were not taking psychoactive medication.
Standard protocol approvals, registrations, and patient consents
This study was approved by the local ethics committee and it was performed according to the Helsinki Declaration. Informed consent was obtained from all participants.
FDG-PET image acquisition and preprocessing
All subjects underwent an FDG-PET examination using a Discovery 690 or Discovery STE CT-PET scanner (3.27 mm thickness; 5.55 mm in-plane full width at half maximum [FWHM]), which where both manufactured by GE Healthcare medical systems. All FDG-PET acquisition procedures comply with the European Association of Nuclear Medicine guidelines (Varrone et al., 2009). Static emission images were acquired 45 minutes after injecting 185–250 MBq of FDG via a venous cannula, with 15-min scan duration. Attenuation correction was based on CT scans. Experts visually examined each reconstructed image to check for the artifacts. Image preprocessing was performed using SPM12 software, running in MATLAB (MathWorks, Inc., Sherborn, MA). Each FDG-PET image was spatially normalized to a specific FDG-PET template in the Montreal Neurological Institute space (Della Rosa et al., 2014; Perani et al., 2014). Then images were spatially smoothed with an isotropic three-dimensional (3D) Gaussian kernel (FWHM: 8–8–8 mm). Global mean scaling was applied to each image to account for between-subject uptake variability (Perani et al., 2014).
Network connectivity analyses
We have addressed neural networks related to dopaminergic neurotransmission systems, that is, the nigro-striato-cortical and mesolimbic pathways, according to well-validated node selection and ROI definition strategy (Caminiti et al., 2017, 2017; Sala et al., 2017; Tziortzi et al., 2013). Specifically, these previous studies assessed metabolic connectivity changes within the dopaminergic nigro-striato-cortical and mesolimbic system in both PD (Caminiti et al., 2017; Sala et al., 2017) and DLB (Caminiti et al., 2017). In the present study, we follow the same node selection and ROI definition approach according to the established related literature.
In detail, the two dopaminergic systems were assembled considering the topographical organization of ascending dopaminergic projections based on biochemical, histochemical, and anatomopathological findings (Ciliax et al., 1999; Fallon 1988; Hall et al., 1999; Meador-woodruff et al., 1996; Ungerstedt and Herrera-marschitz 1970). The dopamine neurons of the SNc, also known as the A9 group, project to the striatum, forming the nigrostriatal pathway, and the dopamine neurons of the VTA gave rise to the mesolimbic and mesocortical pathways (Nieuwenhuys et al., 2013). The topography of the projections from SNc and VTA is organized into three main planes, namely medial–lateral, anterior–posterior, and dorsal–ventral organization (Fallon 1988). The dorsal–ventral plane segregated the nigro-striato-cortical and mesocorticolimbic dopaminergic projections. Thus, the nigro-striato-cortical system consists of SNc projections to the dorsal caudate and dorsal putamen, frontal premotor, motor, executive dorsolateral frontal regions, and somatosensory cortex (Caminiti et al., 2017; Fallon 1988; Sala et al., 2017; Tziortzi et al., 2013). On the contrary, the limbic striatal system consists of VTA projections to the nucleus accumbens, the deep portion of the olfactory tubercle, and the most ventral parts of both the caudate nucleus and the putamen; it receives fibers from limbic and paralimbic cortices, as well as from the amygdala and the hippocampus (Caminiti et al., 2017; Fallon 1988; Parent and Hazrati 1995; Sala et al., 2017; Tziortzi et al., 2013).
We selected the ROIs considering the topographical organization of ascending dopaminergic projections based on biochemical, histochemical, anatomopathological findings (Ciliax et al., 1999; Hall et al., 1999; Meador-woodruff et al., 1996; Parent and Hazrati 1995; Ungerstedt and Herrera-Marschitz 1970). Moreover, we followed a multimodal imaging study that parcelled the human ventral and dorsal striatum into functional ROIs based on their subcortical–cortical connections (Tziortzi et al., 2013).
We also considered a previously validated ROI definition strategy (Caminiti et al., 2017, 2017; Sala et al., 2017). The nigro-striato-cortical network consisted of the dorsal caudate and dorsal putamen, frontal premotor, motor, executive dorsolateral frontal regions, and somatosensory cortex. The mesolimbic network included the ventral striatum, ventral and medial frontal areas, anterior and middle cingulate cortices, as well as the amygdala and the parahippocampal cortex (Supplementary Table S1).
All ROIs used for the connectivity analyses were derived from Automated Anatomical Labeling atlas, except for the bilateral dorsal putamen and caudate, for which we used ROIs derived from Harvard-Oxford subcortical Atlas available in FMRIB Software Library.
The small dopaminergic nuclei, that is, SNc and VTA, were not included in the analysis, due to the limited spatial resolution of PET method and the lack of reference atlases for these regions.
Partial correlation analysis
To assess the connectivity within the nigro-striato-cortical and the mesolimbic dopaminergic pathways in PD females and males, we created the subject-by-node/ROI matrix for each gender subgroup in PD and healthy controls (HC). The matrices contained the regional metabolic mean value extracted from the specific ROIs of each neurotransmitter network for each subject (Supplementary Table S1).
Partial correlations were computed using MATLAB software (Mathworks, Inc., Sherborn, MA). Partial correlation is a statistical approach that allows the measurement of metabolic connectivity between two nodes while factoring out the contributions of other regions (Sala and Perani 2019). This approach is based on the evidence that brain regions, whose metabolism values are significantly correlated, are functionally associated (Horwitz et al., 1984; Sala and Perani 2019). This multivariate method relies on good reproducibility and general applicability within the range of typical experimental settings of PET neuroimaging studies (Veronese et al., 2019). The resulting partial correlation matrices were set at a statistical threshold p < 0.01, uncorrected for multiple comparisons. Age was included as a nuisance covariate in the analyses.
The resulting matrices were represented in a 3D brain template using BrainNet toolbox (Xia et al., 2013).
We applied the Fisher's transformation to the partial correlation coefficients to test whether the strength of each coefficient (between nodes and dopaminergic networks) differed between groups (PD and HC). Z-test was used to test for significant changes in partial correlation coefficients. All the presented results were set at a statistical threshold p < 0.01, corrected for Bonferroni multiple comparisons.
Connectivity measures and statistical analyses
We calculated the percentage of altered metabolic connections of each network (i.e., connections linking two nodes in the same matrix derived from the comparison between patients and HC) to quantify the severity of metabolic connectivity alterations in PD males and females. Specifically, we computed the total number of connections and the number of altered connections to calculate the percentage of altered metabolic connectivity of each network. Then, we compared the percentage of altered metabolic connections between males and females for each network through the chi-squared test. Thus, chi-square tests of independence examined the relationship between gender and the altered connectivity of dopaminergic networks.
Results
The demographical and the clinical data of the whole PD cohort are reported in Table 1. Males and females did not show significant differences in demographical and clinical data, including both cognitive and motor assessment and medication state (Table 1).
Dopaminergic network analysis results
Nigro-striato-cortical network
A widespread alteration of metabolic connectivity characterized PD males within the nigro-striato-cortical dopaminergic network, with 34.69% of altered connections in comparison with HC (p-value <0.01, corrected for Bonferroni multiple comparisons) (Fig. 1). On the contrary, the connectivity reconfiguration of the same network was relatively spared in PD females (only 17.35% changes in comparison with HC) (p-value <0.01, corrected for Bonferroni multiple comparisons) (Fig. 1). The direct comparison between PD males and females showed that the metabolic connectivity alteration of nigro-striato-cortical network was significantly more severe in PD males compared with PD females (χ 2 = 7.65, p-value <0.005) (Fig. 1). Since the p-value is significant, the observed difference is not due to chance (assumption H0), but due to the gender factor.

Nigro-striato-cortical dopaminergic network analyses. 3D brain templates display
Mesolimbic network
Significant widespread alterations of connections within the mesolimbic dopaminergic network characterized PD females, reporting 55.10% of altered connections in comparison with HC (p-value <0.01, corrected for Bonferroni multiple comparisons). On the contrary, the connectivity reconfiguration of the same network was limited in PD males compared with HC (1.02%) (p-value <0.01, corrected for Bonferroni multiple comparisons) (Figs. 2 and 3). The direct comparison between PD males and females showed that the metabolic connectivity alteration of the mesolimbic network was significantly more severe in PD females compared with PD males (χ 2 = 70.99, p-value <0.00001) (Figs. 2 and 3). Since the p-value is significant, the observed difference is not due to chance (assumption H0), but due to the gender factor.
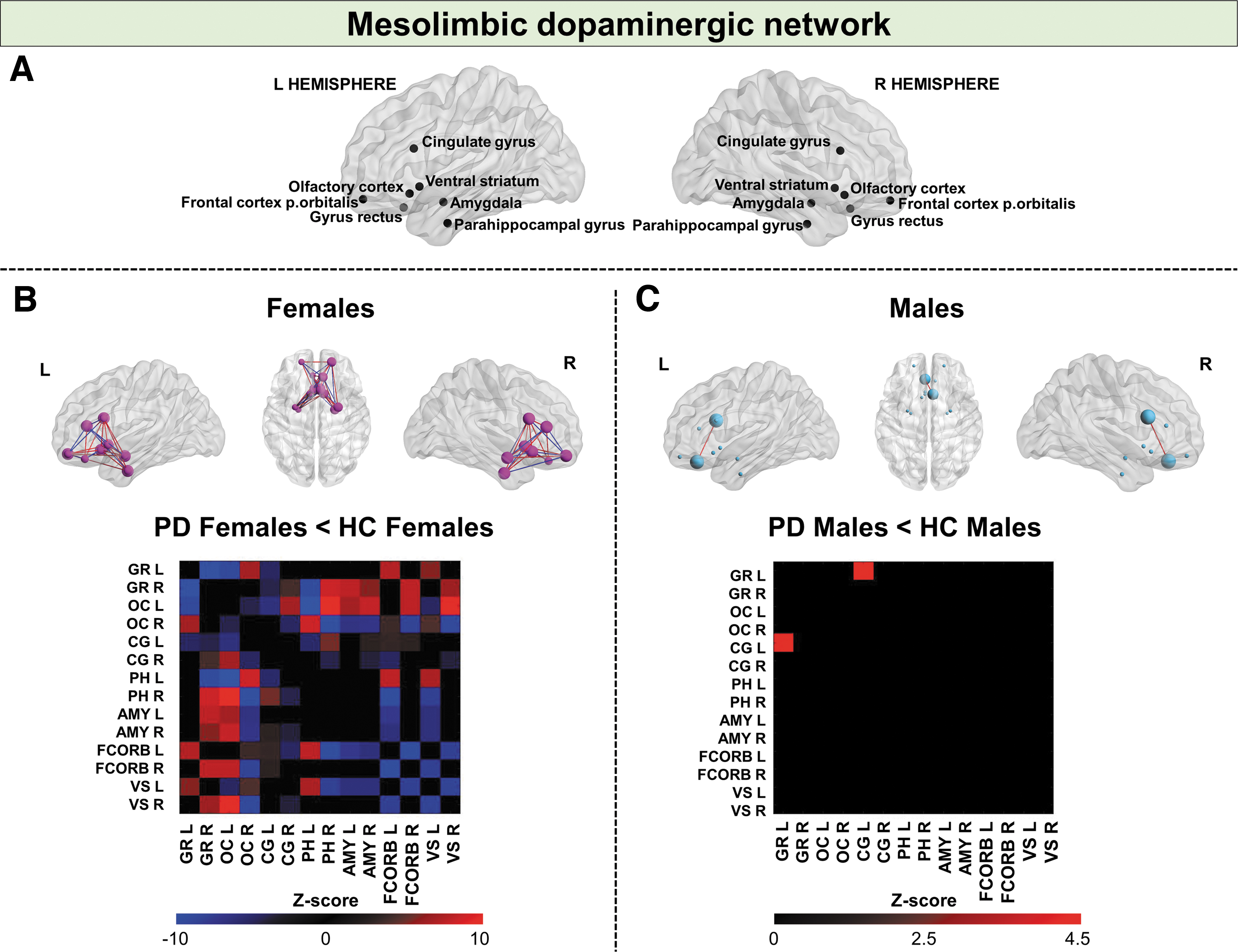
Mesolimbic dopaminergic network analyses. 3D brain templates display
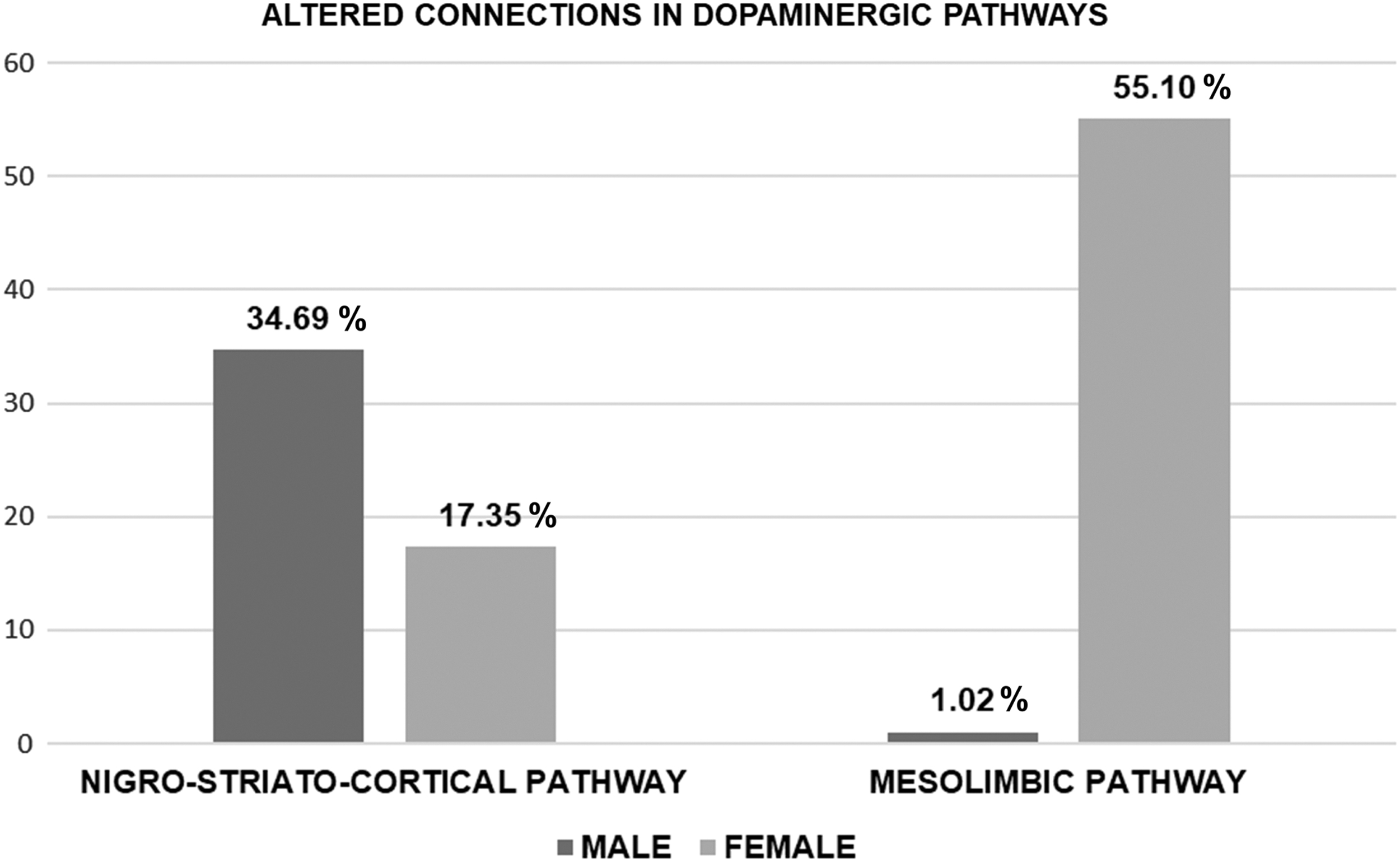
The histograms show the percentage of altered connections in the dopaminergic pathways compared with the control group: nigro-striato-cortical and mesolimbic networks in PD females (light grey) and males (dark grey).
Discussion
We report gender-specific effects on brain metabolic connectivity of dopaminergic networks in a cohort of PD patients. We considered the nigro-striato-cortical and the mesolimbic dopaminergic pathways linked to specific clinical PD features (Castrioto et al., 2016; Surmeier and Sulzer 2013). Our results show a widespread alteration of metabolic connectivity within the nigro-striato-cortical network in PD males (Figs. 1 and 3). On the contrary, the connectivity of the mesolimbic dopaminergic system was remarkably reconfigured in women (Figs. 2 and 3). On the methodological side, the present study is based on a metabolic connectivity approach that has demonstrated good reproducibility and applicability on PET-based assessments of brain connectivity (Veronese et al., 2019), encouraging its use to investigate the neurotransmission network characteristics and the changes in neurological conditions. Subsequently, with the use of this approach, previously published studies revealed metabolic connectivity alterations within the nigrostriatal dopaminergic and mesolimbic system, in PD and DLB (Caminiti et al., 2017; Sala et al., 2017).
Sex hormones play a crucial role in gender differences in PD providing disease protection in women (Gillies and McArthur 2010). The nigrostriatal dopaminergic system expresses high densities of estrogen and progesterone receptors in the human brain, determining its vulnerability to the modulation of sex hormones (Barth et al., 2015). Estrogens show neuroprotective effects on the nigrostriatal dopaminergic system. It has been proposed as a supportive effect of estradiol on DAT, leading to a greater amount of striatal dopamine availability in women (Gillies and McArthur 2010). Following this evidence, we have described a more preserved nigro-striato-cortical dopaminergic network in PD females compared with males, supporting a possible protective role of estrogens on the dopaminergic nigro-striato-cortical system. Thus, the development of symptomatic PD is delayed in women by higher physiological dopamine levels on the striatum due to the activity of estrogens. The estradiol-induced neuroprotection might be an adaptive response in the surviving neurons, restoring striatal dopaminergic functionality (Gillies and McArthur 2010).
In addition to sex hormones, a variety of broader social factors (e.g., role expectations and social attitudes) might modulate the course and outcome of neurodegenerative diseases. For example, gender differences can be noticed in the types of education and occupation due to gender roles, behaviors, and activities. A recent study highlighted gender differences in the effects of education and occupation on brain metabolism and cognitive reserve in AD (Malpetti et al., 2017). In this context, the gender differences obtained in the metabolic connectivity may also be related to environmental factors, such as individual learning experiences, culture, or gender stereotypes. Indeed, gender roles, behaviors, and social activities due to historical and cultural aspects may influence the brain organization and the vulnerability and integrity of the brain itself. All of the above suggest that different connectivity reconfigurations of dopaminergic networks that were revealed in women and men might be due to the interaction between sex-biological and social factors.
α-Synuclein plays a key role in dopaminergic neurotransmission pathways, regulating the filling and fusion of vesicles via vesicular monoamine transporter 2, the dopamine reuptake via DAT, and the postsynaptic dopamine activity (Calo et al., 2016). Previous evidence acquired using the very same metabolic approach in a PD cohort showed a major vulnerability of the nigro-striato-cortical in comparison with the mesolimbic pathway (Sala et al., 2017). Considering PD males and females separately, specific metabolic connectivity alterations emerged in the present study, suggesting that gender may play a role in the variability of PD pathological substrate (Figs. 1 and 2). Specifically, we found that the metabolic connectivity reconfiguration characterized by connectivity decreases and increases (Figs. 1 and 2). Connectivity changes are usually interpreted in terms of function: connectivity decrements may indicate functional disconnection between regions, while connectivity increments indicate increased functional coupling between regions. However, increased connectivity that affects metabolically impaired brain regions might indicate a maladaptive mechanism, such as the disrupted excitatory–inhibitory balance of damaged networks due to underlying pathological processes (Pievani et al., 2014; Sala and Perani 2019).
The present results concerning the nigro-striato-cortical pathway are in line with other data obtained in PD patients using single-photon emission computerized tomography imaging, where women show greater striatal DAT activity than men (Eusebio et al., 2012; Kaasinen et al., 2015; Koch et al., 2014). The reduced vulnerability in the female nigro-striato-cortical dopaminergic system might be explained by the estradiol-induced neuroprotection on this system, as suggested (Cerri et al., 2019; Gillies and McArthur 2010; Haaxma et al., 2007). It is well known that estradiol increases the synthesis, release, reuptake, and the turnover of dopamine (Cerri et al., 2019). Studies mostly point toward a neuroprotective effect of estrogen on dopaminergic systems and toward a consequent reduction of risk for PD. These brain differences may account for the gender-related differences in motor symptoms that are primarily caused by the degeneration of the nigrostriatal dopamine neurons (McCann et al., 2014). Coherently, greater motor impairment was found across a large PD cohort, mainly in men compared with women (Lubomski et al., 2014). Specifically, men demonstrated higher Unified Parkinson's Disease Rating Scale-III scores and required a significantly higher daily levodopa equivalent dose for therapy. Moreover, a recent study investigating the neurophysiological sex differences in early PD reported that female patients have a more favorable profile during transcranial magnetic stimulation, as suggested by the preserved physiological focal response to sensorimotor plasticity compared with men (Kolmancic et al., 2019). However, the mechanisms through which estrogens act on the dopaminergic system protect against the α-synuclein-related neurodegeneration and the consequent motor clinical manifestations are not yet fully understood (Cerri et al., 2019). Moreover, there is also a lack of data regarding whether the estrogen has a neuroprotective effect on the mesolimbic system, despite its involvement in PD clinical nonmotor symptoms.
In this study, we also considered the mesolimbic system connectivity. Our results demonstrated a major vulnerability in PD females in this system, which is relatively spared in men (Figs. 2 and 3). The mesolimbic system impairment in humans shows high intersubject variability, as demonstrated by postmortem studies (Surmeier and Sulzer 2013) and in vivo molecular data (Caminiti et al., 2017). Although the α-synuclein pathology is present in the VTA, its neurons seem relatively spared to the overexpression of human A53T mutated α-synuclein compared with SNc neurons (Maingay et al., 2006). The specific mechanisms defending VTA dopaminergic neurons from neurodegeneration are still unclear (Calo et al., 2016). In this context, our findings suggest that gender may represent a crucial source of this variability in PD patients. This conclusion is consistent with previous studies that demonstrated that several psychiatric symptoms, such as apathy, depression, anxiety, and fatigue, are more common and severe in women (Martinez-Martin et al., 2012). In PD, impairment in the mesolimbic dopaminergic system has been linked to nonmotor symptoms, in particular mood disorders, which may affect patients since or even before the initial clinical phase of PD (Castrioto et al., 2016; Gustafsson et al., 2015). These findings would also fit with the higher rate of impulse compulsive disorder that resulted from the use of dopaminergic drugs observed in males (Premi et al., 2016). Further studies will be critical for a better understanding of the neurobiological bases of the clinical different phenotypes in women and men.
The gender-related vulnerabilities here reported might be explained also by sociodemographic factors, as briefly stated above. Educational, behavioral, and lifestyle choices might affect the organization of specific brain networks (Malpetti et al., 2017). In particular, limbic structures are sensitive to sex hormones, leading to high stress vulnerability of this system in females (McLaughlin et al., 2009). Coherently, the mesolimbic dopamine system seems to be more sensitive to social defeat in females compared with males (Greenberg and Trainor 2015). As supportive evidence, the cerebral metabolism and the metabolic connectivity in aging and AD dementia are affected by occupation and education with clear gender differences (Malpetti et al., 2017). These gender brain discrepancies might be also explained by sociodemographic factors that can consequently contribute to differences between females and males in brain functions and networks. The female widespread mesolimbic connectivity reconfiguration here reported could be explained as a function of gender-related interactions between biological and social elements.
Limitations
From the methodological standpoint, it must be acknowledged that despite the high accuracy and reproducibility of covariance FDG-PET network analysis in the assessment of neurotransmitter system connectivity (Veronese et al., 2019), some cortical regions receive overlapping projections from different neurotransmitter systems. Thus, results obtained with metabolic connectivity based on FDG-PET data might lose a portion of their specificity.
We also acknowledge that our data do not address the relationship between clinical characteristics and brain metabolic connectivity changes in the dopaminergic neural networks, due to data missing for the retrospective nature of the study.
Conclusion
Our findings indicate a different gender-related vulnerability in the dopaminergic systems that are crucially involved in PD, using a metabolic connectivity approach. This multivariate approach allows to reliably reconstruct the brain connectivity patterns within and between neurotransmitter systems, providing in vivo access to the functional architecture of the brain (Sala and Perani 2019). In this way, we found gender differences in the vulnerability of the nigro-striato-cortical and the mesolimbic pathways. Our results support a specific gender vulnerability of the dopaminergic networks in PD, which might suggest different pathological substrates underlying the motor and psychiatric symptoms that characterize clinical manifestation of male and female PD (Cerri et al., 2019). In this context, our study enhances the knowledge on the biological basis of PD gender differences, which may lead to different clinical phenotypes. Further studies are necessary to investigate the direct association between gender-specific brain connectivity patterns and related clinical manifestation for an overall picture in prevention, as well as for diagnostic and treatment strategies.
Compliance with Ethical Standards
The experimental procedure was approved by the local ethics committee. The protocol conformed to the ethical standards of the Declaration of Helsinki for protection of human subjects. Informed consent was obtained from all participants included in the study.
Footnotes
Authors' Contributions
C.B.: Data curation, formal analysis, investigation, methodology, visualization, and writing—original draft. G.C.: Data curation, formal analysis, and writing—review and editing. A.P.: Resources and investigation. A.P.: Resources and investigation. D.P.: Conceptualization, funding acquisition, investigation, resources, project administration, supervision, and writing—review and editing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the CARIPLO Project “Evaluation of autonomic, genetic, imaging, and biochemical markers for Parkinson-related dementia: longitudinal assessment of a PD cohort” 2016–2020 (grant agreement no. 2014-0832)” (D.P.).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.