
Abstract
Background:
The cognitive dysmetria theory of schizophrenia proposes that communication between the cerebellum and cerebral cortex is disrupted by structural and functional abnormalities, resulting in psychotic symptoms and cognitive deficits.
Methods:
Using publicly available data, resting-state functional connectivity (rsFC) was calculated from 20 hemispheric cerebellar lobules as seed regions of interest to the rest of the brain. Group differences in rsFC between individuals with schizophrenia (SZ) and healthy controls (HCs) were computed, and relationships between rsFC and symptom severity and cognitive functioning were explored.
Results:
HCs demonstrated stronger connectivity than SZ between several cerebellar lobules and cortical regions, most robustly between motor-related cerebellar lobules (V and VIIIa/b) and temporal and parietal cortices. In addition, seven of nine lobules in which reduced cerebellocortical connectivity was observed showed diagnosis × processing speed interactions; HC showed a positive relationship between connectivity and processing speed, whereas SZ did not show this relationship. Other cognitive domains and symptom severity did not show relationships with connectivity.
Conclusions:
These findings partially support the cognitive dysmetria theory, and suggest that disrupted cerebellocortical connectivity is associated with slowed processing speed in schizophrenia.
Impact statement
We show in this work that in chronic schizophrenia, there is weaker functional connectivity between previously unstudied inferior posterior cerebellar lobules and cortical association areas. These findings align and extend previous work showing abnormal connectivity of anterior cerebellar lobules. Further, we present a novel finding that these connectivity deficits are differentially associated with processing speed in the schizophrenia versus healthy control groups. Findings provide further evidence for cerebellocortical dysconnectivity and processing speed deficits as biomarkers of schizophrenia, which may have implications for downstream effects on higher order cognitive functions, in line with the cognitive dysmetria theory.
Introduction
Although the cerebellum has traditionally been recognized as a motor modulator, it is structurally and functionally connected to most of the cerebral cortex and contributes to many cognitive domains, including language, executive function, emotion processing, and working memory (Andreasen and Pierson, 2008; Andreasen et al., 1998; Bernard et al., 2014; Bostan et al., 2013; Buckner et al., 2011; Koziol et al., 2014; Stoodley and Schmahmann, 2009). It coordinates and automates both movement and thought through internal models, based on the parts of the cerebral cortex from which it receives input (Ito, 2008; Ramnani, 2006, 2014); therefore, when the cerebellum and its connections are not functioning efficiently, disorganized and uncoordinated thoughts may occur, which may manifest as psychotic symptoms or cognitive deficits (Andreasen and Pierson, 2008; Schmahmann and Sherman, 1998).
Jeremy Schmahmann observed dysmetria of thought and described the cerebellar cognitive affective syndrome due to cerebellar damage, which includes a cluster of cognitive and behavior deficits often in the domains of executive function, language, visuospatial functioning, memory, and behavior regulation (Schmahmann and Sherman, 1998). Similarly, Nancy Andreasen's cognitive dysmetria theory proposes that disrupted connectivity between the prefrontal cortex, thalamus, and cerebellum may underlie psychotic symptoms and cognitive deficits of schizophrenia (Andreasen and Pierson, 2008; Andreasen et al., 1998).
Resting-state functional connectivity (rsFC) illustrates that the cerebellum is included in most major intrinsic networks, such as default mode network, cingulo-opercular network, and frontoparietal network (Buckner et al., 2011; Habas et al., 2009). Cerebellar functional connectivity has been shown to discriminate between controls and individuals with schizophrenia with high precision and better than cortical networks (Peters et al., 2016; Shen et al., 2010). Cerebellocortical connectivity deficits have been observed in adolescents at risk of psychosis (Anticevic et al., 2015; Bernard et al., 2014), siblings of those with schizophrenia (Collin et al., 2011; Repovs et al., 2011), early schizophrenia (Du et al., 2018), and chronic schizophrenia (Bernard et al., 2017; Chen et al., 2013; Repovs et al., 2011; Wang et al., 2014; Zhuo et al., 2017), but some studies also show stronger connectivity in the psychosis spectrum (Chen et al., 2013; Collin et al., 2011; Du et al., 2018; Walther et al., 2017; Zhuo et al., 2017).
Most studies demonstrating cerebellocortical dysconnectivity thus far were either incidental findings or investigated only a few regions of interest (ROIs), rather than connectivity of multiple cerebellar lobules (Bernard et al., 2017; Chen et al., 2013; Wang et al., 2014). Thus, we aimed to compute cerebellocortical connectivity using all hemispheric cerebellar lobules as seed ROIs providing a more comprehensive approach and including inferior lobules.
Further, we wished to investigate relationships between cerebellocortical connectivity with symptom severity and cognitive function, in line with the cognitive dysmetria theory (Andreasen et al., 1998). A meta-analysis has revealed abnormal cerebellar activation across cognitive domains in schizophrenia (Bernard and Mittal, 2015). However, relationships between cerebellocortical connectivity and specific domains of cognitive functioning have not been comprehensively explored, despite findings that functional connectivity strength and amplitude of low-frequency fluctuations in other networks predicted overall performance on a comprehensive neurocognitive battery (MATRICS Consensus Cognitive battery [MCCB]) (Sui et al., 2015; Wu et al., 2018).
Using a composite measure of cognitive domains may only be informative of global cognitive dysfunction, however, and does not specify deficits in domains such as memory, processing speed, and executive functioning (Wu et al., 2018). Specific structural and functional imaging measures have been associated with performance on specific MCCB domains in schizophrenia; however, the cerebellum was not included (Sui et al., 2015). One study did find that lower connectivity between the cerebellum and frontoparietal network predicted poor working memory in schizophrenia (Repovs et al., 2011), but no other studies have sought to specifically and comprehensively investigate relationships between cerebellocortical connectivity and MCCB cognitive domains in schizophrenia.
In addition, work connecting cerebellocortical connectivity to psychotic symptom severity has been inconsistent. Weaker cerebellar connectivity predicted presence of auditory verbal hallucinations in one study (Chang et al., 2015). Further, posterior cerebellum–prefrontal cortex dysconnectivity predicted negative symptom severity, and transcranial magnetic stimulation to the cerebellum increased connectivity while decreasing symptom severity (O'Brady et al., 2019). However, other recent work has not demonstrated cerebellocortical connectivity relationships with total positive or negative symptom severity (Bernard et al., 2017; Wu et al., 2018). Therefore, investigating more specific symptom profiles rather than broad positive and negative symptom domains may yield more information about neurobiological mechanisms of schizophrenia.
While there is some evidence that a few cerebellar lobules may demonstrate weaker cerebellocortical connectivity, no comprehensive work has investigated all hemispheric cerebellar lobules, particularly the inferior posterior cerebellum (e.g., lobules VIIIa and VIIIb), potentially leaving important cerebellar regions understudied. Thus, the first goal of our analysis was to investigate rsFC between the whole brain (cortical and subcortical) and all individual hemispheric cerebellar lobules in individuals with schizophrenia and schizoaffective disorder and healthy controls (HCs). We hypothesized that the schizophrenia group would demonstrate weaker cerebellocortical connectivity in general.
We also tested whether connectivity differences were correlated with cognitive and symptom measures, specifically investigating group × cognition interaction effects using specific domain scores (Sui et al., 2015). While the schizophrenia group is expected to demonstrate weaker cerebellocortical connectivity compared with controls, we examined whether it is general connectivity or more regionally specific connectivity deficits, which are related to poorer cognition.
Methods
Participants
Participants included HCs and individuals with schizophrenia or schizoaffective disorder (SZ), a subset from the Center of Biomedical Research Excellence (COBRE) study from the Mind Research Network (MRN), Albuquerque, NM (Aine et al., 2017; Çetin et al., 2014). The Georgia State University IRB approved obtaining and analyzing deidentified data for this study, accessible through the COINS system (Wood et al., 2014). Written informed consent was obtained from all participants at MRN.
Participants were outpatients and clinically stable on antipsychotic medications. Exclusion criteria included history of neurological disorder, severe head trauma with >5 min loss of consciousness, severe motor dysfunction, and history of substance abuse or dependence within the last 12 months (except for nicotine). Motor dysfunction and substance abuse exclusion criteria were especially important because the cerebellum is known to be vulnerable to substance abuse and highly associated with motor dysfunction. For this imaging study, participants were also excluded if their volume-to-volume in-scanner head motion was >3 mm or 3° in any direction. The final sample size included 85 HC and 73 SZ.
The Structured Clinical Interview for DSM-IV-TR Disorders (SCID-II) was used to establish diagnoses. The Positive and Negative Syndrome Scale (PANSS) was administered to the SZ group to measure symptom severity. PANSS items were summed into five empirically supported factors according to the “narrow definition” of Lehoux and colleagues' (2009) critical review: negative, positive, disorganized, excited, and anxiety/depression (Table 1). Dosages of medications were converted to olanzapine equivalents to measure medication load (Gardner et al., 2010). The Wechsler Abbreviated Intelligence Scale, Second Edition (WASI-II) was administered to estimate intelligence quotient (IQ) of all participants. Table 1 presents participant demographic details.
Participant Demographic, Clinical, and Cognitive Characteristics (N = 158)
Symptom severity was assessed by the narrow definition of the PANSS five factor solutions (Lehoux et al., 2009). Cognition was assessed with the MCCB.
HC, healthy controls; IQ, intelligence quotient; MCCB, MATRICS Consensus Cognitive battery; PANSS, Positive and Negative Syndrome Scale; SZ, individuals with schizophrenia; WASI-II, Wechsler Abbreviated Intelligence Scale, Second Edition.
The MCCB was utilized to measure neurocognitive functioning in most participants (HC n = 77–79; SZ n = 65–69, depending on number of participants who completed each measure). The MCCB includes 10 tests that assess cognitive domains of Processing Speed, Attention/Vigilance, Working Memory, Verbal Learning, Visual Learning, Reasoning and Problem Solving, and Social Cognition (Kern et al., 2008). Raw scores from cognitive tests were standardized to domain T scores following the procedure outlined in Kern and colleagues (2008). Two-sample t-tests were utilized to examine group differences in all MCCB domains.
Functional magnetic resonance imaging data acquisition and processing
Participants underwent a resting-state (T2*-weighted) functional magnetic resonance imaging (fMRI) scan, on a 3T Siemens Trio using a 12-channel head coil: TR = 2 sec, TE = 29 ms, matrix size: 64 × 64, 32 slices (ascending order), voxel size: 3 × 3 × 4 mm3 (Aine et al., 2017). During the scan, participants were instructed to stare at a fixation cross for 5 min.
Preprocessing was performed according to a standard pipeline using the Data Processing Assistant For Resting-State fMRI, Advanced edition (DPARSFA) (Yan and Zang, 2010) implemented in Statistical Parametric Mapping-8 (SPM8) and MATLAB R2013a. The first four volumes were discarded, then the following steps were performed: slice timing correction, realignment, coregistration to T1, normalization to the MNI template using SPM unified segmentation, smoothing with a 6 mm full width at half maximum Gaussian kernel, nuisance covariate regression (white matter, cerebrospinal fluid, 12 motion parameters [6 + their derivatives], linear trend), and low-pass filtering (0.01–0.08 Hz).
rsFC was calculated using DPARSFA with 20 hemispheric cerebellar lobules as seed ROIs (Supplementary Fig. S1). Masks were created utilizing the Spatially Unbiased Infratentorial Template atlas and are consistent with those used in previous work (Bernard et al., 2012, 2014; Clark et al., 2018; Diedrichsen, 2006). Connectivity was computed as Pearson's correlations between timecourses of each ROI and all voxels in the brain. This computation resulted in seed-to-voxel maps of Fisher's Z-transformed correlations across the entire brain for each of the cerebellar ROIs for each subject.
Statistical analyses
Whole-brain connectivity maps from each of the 20 lobules were individually entered into two-sample t-tests in SPM12 to assess group differences, covarying for mean framewise displacement (FD) (Power et al., 2014), age, and gender. Results were masked with a gray matter mask excluding the cerebellum to investigate connectivity between the cerebellum and the rest of the brain. The mask was created in SPM8 with the PickAtlas tool (Maldjian, et al., 2003). Results were first thresholded with an uncorrected cluster-forming threshold of p < 0.001. We also performed a Bonferroni correction for the number of lobules we tested. Significant clusters were identified using the SPM-generated cluster threshold of q FDR < 0.0025 (0.05/20 tests = 0.0025). The SPM-estimated cluster size threshold was >150 voxels for all t-tests, and >300 voxels for the majority. Masks were created of significant clusters for use in further analyses using SPM, at a more lenient threshold of q FDR < 0.05.
We then explored relationships between connectivity and clinical and cognitive measures, including MCCB performance and PANSS factors. Because the cerebellum is sensitive to the effects of chronic alcohol use, we tested for group differences in connectivity between those who have a history of alcohol abuse and dependence and those who do not; there were no significant differences. We also tested for connectivity differences between those who are moderately to severely addicted to nicotine and those who are not; no significant differences were observed.
First, multiple linear regression was used to explore the relationships between PANSS factors and cerebellocortical connectivity in the schizophrenia group alone. Three participants were not included in this analysis due to missing data. For each of the nine cerebellar lobules in which we observed significant group connectivity differences, we entered the five PANSS factors as predictors and age, gender, and FD as covariates of no interest. Results were explicitly masked by t-test results (uncorrected p < 0.001, cluster q FDR < 0.05). We also performed a Bonferroni correction for the number of lobules we tested (0.05/9 = 0.0056). Therefore, significant clusters were identified using the SPM-generated cluster threshold of q FDR < 0.0056.
Second, flexible factorial models were run from the nine ROIs that demonstrated significant group connectivity differences. Twenty-one participants missing MCCB scores were excluded from cognitive analyses (HC n = 13, SZ n = 8). For each ROI, group was entered as a factor of interest, gender was entered as a factor of no interest, age, gender, and IQ were entered as covariates of no interest, and all seven MCCB domain T scores were included as covariates of interest (Processing Speed, Attention/Vigilance, Working Memory, Verbal Learning, Visual Learning, Reasoning/Problem Solving, and Social Cognition). Group × MCCB domain interaction terms were also included in the models. We tested for main effects of all MCCB domain T scores and Group × MCCB domain interactions. Again, significant clusters were identified using the SPM-generated cluster threshold of q FDR < 0.0056 based on the number of ROIs tested.
Results
Group differences in cerebellocortical connectivity
The control group demonstrated significantly stronger cerebellocortical connectivity than the schizophrenia group from nine ROIs, which included both anterior and posterior cerebellar lobules (Table 2 and Fig. 1). Supplementary Figure S2 shows extent of cerebello-cortical connectivity in each group separately, visualized by one-sample t-tests. In line with recent work demonstrating distinct cognitive and motor cerebellar activation and motor-attentional/executive-default mode gradients in a large dataset (Guell and Schmahmann, 2020; Guell et al., 2018), we will refer to “cognitive”- and “motor”-associated lobules based on their primary functions. While lobules VIIIa/b have shown activity during cognitive tasks, we refer to them as secondary motor lobules for consistency with the majority of their topography being associated with motor function, while acknowledging that there is some overlap with attention/executive function in these lobules (Guell and Schmahmann, 2020).

Clusters demonstrating stronger cerebellocortical connectivity in the control group. Clusters resulted from two-sample t-tests comparing individuals with schizophrenia and healthy controls, controlling for age, gender, and framewise displacement. Results were masked with a gray matter mask excluding the cerebellum and thresholded at uncorrected p < 0.001 and SPM-defined cluster q FDR < 0.0025. The minimum T value was 3.14, and maximum T values are depicted on the right side of the figure. Cerebellar regions of interest are labeled on the right side of the figure and shown in coronal sections. Left side of the image represents the left side of the brain. Note, while right and left lobule VIIIa/b figures show some cerebellar activation, this was excluded from analysis with the cortical gray matter mask. The result is visualized using the xjView toolbox. Color images are available online.
Regions in Which the Control Group Demonstrated Significantly Greater Cerebellocortical Connectivity Than the Schizophrenia Group
Regions of interest from which connectivity was calculated appear above the significant cluster coordinates, statistics, and brain regions. t-Tests were performed in SPM12, including age, gender, and framewise displacement as covariates. Results were masked with a gray matter mask excluding the cerebellum and thresholded at uncorrected p < 0.001 and SPM-defined cluster q FDR < 0.0025. Regions were identified using xjView.
FDR, false discovery rate; L, left; MNI, Montreal Neurological Institute; R, right; SPM, Statistical Parametric Mapping.
Left lobule VIIb, a secondary cognition-related lobule, showed stronger connectivity in HC with right prefrontal regions, including middle and inferior frontal gyrus.
Regarding motor-associated lobules, connectivity was stronger in the control group throughout much of the cortex. Primary motor lobules I–IV and V showed stronger connectivity in HC with largely parietal and temporal regions, including bilateral precuneus, inferior parietal lobe, and somatosensory cortex; stronger connectivity was also observed in the motor cortex, supplementary motor area, and inferior frontal gyrus. Secondary motor lobules VIIIa and VIIIb also showed widespread connectivity differences between groups. Regions with which these lobules showed stronger connectivity in HCs were mainly located in temporal and parietal areas, including the temporoparietal junction, supramarginal gyrus, angular gyrus, somatosensory cortex, and precuneus; stronger connectivity was also observed in the inferior frontal gyrus and anterior cingulate.
There were no ROIs in which the SZ group demonstrated stronger connectivity than the HC group. Medication dosage and symptom severity did not significantly predict cerebellocortical connectivity for any ROIs. In addition, after controlling for age, gender, and FD, no PANSS symptom factors showed significant relationships with cerebellocortical connectivity in SZ.
Relationships between cerebellocortical connectivity and cognition
Regarding MCCB domains as predictors of cerebellocortical connectivity, diagnosis × processing speed was the only significant interaction for several lobules (Fig. 2 and Table 3). No main effects of any cognitive measure were significant for any lobules. Group × processing speed interactions were present for connectivity from motor lobules I–IV, V, VIIIa, and VIIIb. Interactions showed stronger relationships between connectivity and processing speed in the control group mostly with the supplementary motor and somatosensory areas for primary motor lobules, and with the temporoparietal junction and precuneus for secondary motor lobules. See Supplementary Figure S3 for interaction graphs.
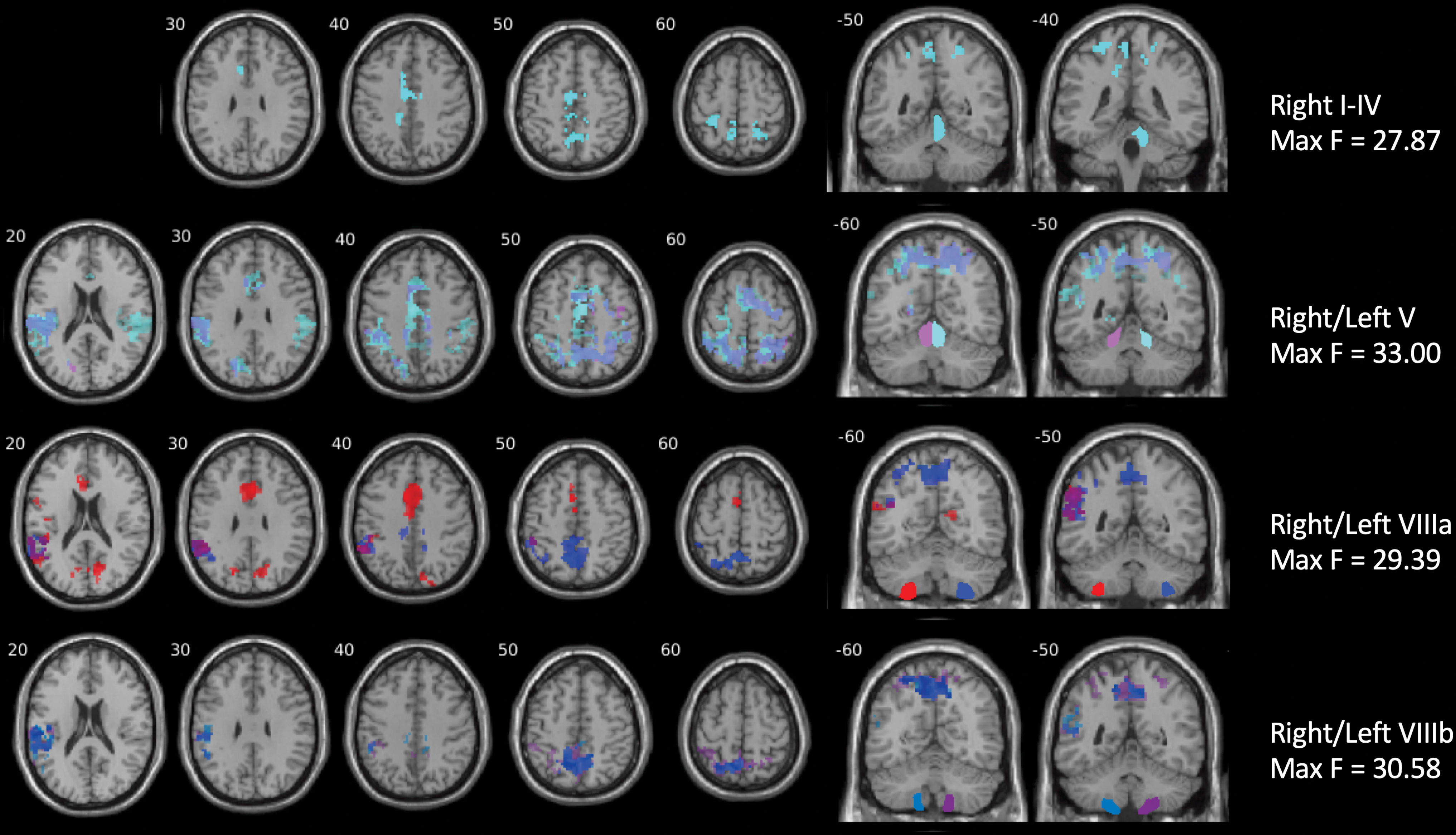
Significant clusters demonstrating Group × Processing Speed interactions in areas showing weaker cerebellocortical connectivity in schizophrenia. Regions of interest are labeled on the right side of the figure. A flexible factorial model was utilized in SPM12, with group as a factor of interest, gender as a factor of no interest, age and framewise displacement as covariates, and seven MCCB cognitive domains as predictors of interest. Group × MCCB domain interaction terms were also included. Results were masked with significant clusters resulting from t-tests and thresholded at uncorrected p < 0.001 and SPM-defined cluster q FDR < 0.0056. The minimum F value was 11.38, and maximum F values are depicted on the right side of the figure. Regions of interest are labeled on the right side of the figure and shown in coronal sections. Left side of the image represents the left side of the brain. The result is visualized using the xjView toolbox. MCCB, MATRICS Consensus Cognitive Battery. Color images are available online.
Regions in Which Group × Processing Speed Interactions Were Observed
Regions of interest from which connectivity was calculated appear above the significant cluster coordinates, statistics, and brain regions. A flexible factorial model was utilized in SPM12, with group and gender as factors, age and framewise displacement as covariates, and seven MCCB cognitive domains as predictors of interest. Group × MCCB domain interaction terms were also included. Results were masked with significant clusters resulting from t-tests and thresholded at uncorrected p < 0.001 and SPM-defined cluster q FDR < 0.0056. Regions were identified using xjView.
Discussion
We investigated rsFC of 20 individual hemispheric cerebellar lobules in schizophrenia and demonstrated widespread rsFC deficits between specific cerebellar lobules and cerebral cortex, as well as associations between connectivity and processing speed measured by the MCCB.
First, HC showed stronger connectivity between cerebellar lobules and cortical regions than individuals with schizophrenia, with the strongest results located in primary and secondary motor lobules (V and VIIIa/b). Second, several of these areas that showed significant connectivity reductions also showed diagnosis × processing speed interactions in which HCs showed positive relationships between connectivity and processing speed but SZ showed no positive relationships. No areas of stronger connectivity were observed in the schizophrenia group, though both stronger and weaker cerebellocortical connectivity compared with controls have been reported elsewhere (Bernard et al., 2017; Walther et al., 2017).
Weaker cerebellocortical rsFC in our SZ group is consistent with previous research in psychosis and typical rsFC (Anticevic et al., 2015; Bernard et al., 2017; Chang et al., 2015; Collin et al., 2011; Peters et al., 2016; Repovs et al., 2011; Shen et al., 2010; Wang et al., 2014, 2016), and we extend these findings to inferior cerebellar lobules that have not yet been investigated. Results broadly align with Friston's dysconnection hypothesis that proposes schizophrenia arises from poor functional integration of brain networks, including cortical–subcortical–cerebellar connections (Friston, 1998; Friston et al., 2016). They also support the cognitive dysmetria theory by showing weaker cerebellocortical connectivity specifically and relationships with cognition (Andreasen et al., 1998).
The most robust differences in cerebellocortical connectivity were observed in motor-associated lobules, I–IV and V, and VIIIa/b. However, the cortical regions from which the cerebellar lobules were disconnected in schizophrenia were located mostly in the precuneus and temporoparietal junction rather than motor and premotor cortex. It is noteworthy that the most extensive connectivity differences were associated with right cerebellar ROIs, but cortical regions of cerebellocortical connectivity differences were not strongly lateralizing and mainly bilateral.
Bilateral dysconnectivity from unilateral ROIs in schizophrenia may be related to previously observed lower hemispheric asymmetry in SZ resting-state connectivity and white matter structure (Agcaoglu et al., 2018; Ho et al., 2017; Hoptman et al., 2012). For instance, if individuals with schizophrenia show less lateralization or interhemispheric communication than controls, this may present as lower cerebellocortical connectivity bilaterally when it is expected that the right cerebellum would be disconnected primarily from the left cortical hemisphere. Swanson and colleagues (2011) showed less resting-state lateralization, and Hoptman and colleagues (2012) showed lower voxel-mirrored homotopic connectivity in several regions in which we observed lower connectivity in SZ (e.g., cerebellum, posterior sensory areas). Thus, our findings may broadly align with and add to this past work in schizophrenia showing weaker lateralization and interhemispheric communication in areas involved in visuospatial, attentional, and sensorimotor processing.
Functionally, lobule V contains the primary motor representation, and lobules VIIIa/b contain a secondary motor representation (Guell et al., 2018; Walther and Strik, 2012), but some studies observe activity during cognitive tasks in lobules VIIIa/b, as well (Bernard and Mittal, 2015; Keren-Happuch t al., 2014; Stoodley and Schmahmann, 2009). While one recent study demonstrated lower effective connectivity in schizophrenia between lobule V and motor cortex during finger tapping (Moussa-Tooks et al., 2018), other studies demonstrated weaker rsFC between anterior cerebellum and nonmotor cortical regions (Bernard et al., 2017; Collin et al., 2011).
A dynamic functional connectivity study also showed weaker connectivity between inferior cerebellum, likely including lobule VIII, and temporal cortex in individuals with early schizophrenia (Du et al., 2018). In contrast, Dong and colleagues (2020) observed compression of the motor-cognitive gradient within the cerebellum in schizophrenia, accompanied by hyperconnectivity between cerebellar sensorimotor and cerebral cognitive areas. While their methods are novel and differ from traditional ROI analyses, their results indicate abnormalities in both cerebellar and cerebellocortical organization and connectivity. Thus, our findings are in keeping with the recent literature observing dysfunctional connectivity in schizophrenia between motor cerebellar lobules and association cortex, suggesting a motor-cognition connection.
Connectivity differences were also observed in one cognitive-associated lobule, left VIIb (Guell et al., 2018). Our findings of lower connectivity in schizophrenia between left lobule VIIb and right prefrontal cortex generally aligned with expectations based on contralateral anatomical connections. In healthy individuals, lobule VIIb is functionally connected to executive networks such as the frontoparietal and cingulo-opercular networks (Buckner et al., 2011; Sang et al., 2012), and is associated with executive functions and working memory (Keren-Happuch et al., 2014; Stoodley and Schmahmann, 2009). However, we did not find that lobule VIIb connectivity differences were associated with the expected cognitive measures.
While we observed widespread lower cerebellocortical connectivity in schizophrenia, connectivity differences were not correlated with psychosis symptoms, consistent with recent studies using the same dataset that did not find cerebellar connectivity to predict symptom severity (Bernard et al., 2017; Wu et al., 2018). We used a different approach to others by investigating PANSS factors rather than the traditional positive and negative subscales (Lehoux et al., 2009); however, these factors do not appear to explain more variance than the positive and negative subscales. Past work by Moberget and colleagues also showed that cerebellar morphology was a strong predictor of psychotic symptoms in a large sample of youth (Moberget et al., 2019), but not in adults with chronic schizophrenia (Moberget et al., 2018), suggesting a possible influence of duration of illness or medication. Overall, it appears that cerebellocortical connectivity does not influence symptoms directly in this chronically ill sample that is stabilized on medication.
With regard to cognitive functioning, individuals with schizophrenia demonstrated severe deficits in all MCCB domains compared with controls, as expected (August et al., 2012; Green et al., 2014; Nuechterlein et al., 2008). Moreover, many regions in which cerebellocortical connectivity was weaker in the SZ group showed diagnosis × processing speed interactions, supporting findings that specific cognitive domains may be related to specific neural substrates (Sui et al., 2015). Specifically, whereas HC showed a positive relationship between connectivity and processing speed within regions of connectivity differences, SZ showed no relationship or a slightly negative relationship.
This interaction suggests that these networks may not be functioning as expected in schizophrenia, which likely influences an individual's ability to quickly process stimuli from the environment, as well as ability to execute a quick response, and may lead to a further cascade of cognitive deficits (Dong et al., 2020). We did not observe relationships with any other cognitive measures, indicating that processing speed is specifically related to these regions of weaker cerebellocortical connectivity in schizophrenia. This finding may be due to the fact that significant connectivity differences were primarily observed between motor-associated lobules and association cortex, highlighting the cerebellum's function in both motor and cognitive efficiency.
Processing speed was assessed with three different tasks: a symbol-digit coding task, Trail Making Test part A, and category fluency, all of which require multiple cognitive and motor processes for good performance (Nuechterlein et al., 2008). These MCCB tasks likely tap into the cerebellum's roles in timing, sequencing, motor coordination, attention, and verbal fluency, which have been localized to anterior and inferior posterior lobules (Caligiore et al., 2017; Fuentes and Bastian, 2007; Guell et al., 2018; Hoche et al., 2018; Koziol et al., 2014; Molinari and Leggio, 2007; Molinari et al., 1997; Schmahmann and Sherman, 1998).
Consistent with our findings in lobules I–V and VIIIa/b, and the nature of these tasks, Bernard and Mittal's (2015) meta-analysis revealed that controls had greater lobule VIII activation during language and motor tasks than individuals with schizophrenia. In addition, individuals with cerebellar cognitive affective syndrome (Schmahmann and Sherman, 1998) show consistent deficits in both psychomotor processing speed and verbal fluency (Hoche et al., 2018). Thus, our findings align with and extend previous work by linking these cognitive deficits to specific regions of lower cerebellocortical connectivity in schizophrenia.
Motor efficiency has been clearly associated with cerebellar function (Koziol et al., 2014; Wolpert and Kawato, 1998), and individuals with schizophrenia have deficits in both timing and motor coordination (Carroll et al., 2009a,b; Walther and Strik, 2012). Further, Fuentes and Bastian (2007) proposed a role for the cerebellum in “motor cognition,” which refers to predicting movement outcomes and understanding the meaning of one's movements. This idea is consistent with the proposed internal models that the cerebellum utilizes to predict and control behavior (Ito, 2008; Ramnani, 2014). For example, individuals with schizophrenia have difficulty distinguishing between self-produced and other-produced tactile stimulation (Blakemore et al., 2000; Shergill et al., 2014), which may be related to deficits in prediction of movement outcomes. The same deficits may be related to impaired processing speed on tasks in which one needs to quickly sequence or predict the correct response.
Cortical areas with which cerebellar lobules showed weaker connectivity in schizophrenia and diagnosis × processing speed interactions were mainly localized to parietal association areas, precuneus, temporoparietal junction, supplementary motor area, and cingulate gyrus. The precuneus and temporoparietal junction are heteromodal association areas responsible for attending to and integrating somatosensory information (Cavanna and Trimble, 2006; Donaldson et al., 2015; Wenderoth et al., 2005). The precuneus has also been associated with visuospatial attention and learning, motor coordination, episodic memory, and self-reflection (Cavanna and Trimble, 2006; Wenderoth et al., 2005). The supplementary motor area is essential for planning movements, consistent with predicting the correct responses on these tasks (Goldberg, 1985). In addition, the cingulate gyrus is part of the cingulo-opercular network, which is important for directing and sustaining attention, as well as detecting errors (Dosenbach et al., 2006, 2008).
Thus, in healthy individuals, the cerebellum may work with these cortical regions in attending to stimuli, coordinating sensorimotor information and motor responses, and predicting or sequencing responses while an individual is performing complex psychomotor tasks (Ito, 2008; Molinari and Leggio, 2007). These processes are likely impaired in schizophrenia. Further, results are consistent with studies of schizophrenia that linked lower cerebellum and temporoparietal activity during visual attention and spatial working memory tasks (Kang et al., 2011; Laurens et al., 2005), and a lack of cerebellar activation during impaired procedural learning performance (Kumari et al., 2002; Siegert et al., 2008), suggesting that the cerebellum is not facilitating these processes efficiently in schizophrenia.
It is well established that individuals with schizophrenia have impaired processing speed; in fact, it is one of the most severely impaired cognitive domains (Heinrichs and Zakzanis, 1998; Kern et al., 2011; Mesholam-Gately et al., 2009). Interestingly, the MCCB standardization study revealed that processing speed was one measure that most accurately distinguished patients from controls, and also distinguished those patients who were employed from those who were not (Kern et al., 2011). Thus, weak cerebellocortical rsFC and associated processing speed deficits may be a clinically meaningful phenotype (Osborne et al., 2020).
Notably, it is important to consider the effect of neuroleptic medications when measuring processing speed; however, processing speed is impaired in first-episode psychosis and may even represent a core cognitive deficit that accounts for much of the variance in other cognitive deficits (Mesholam-Gately et al., 2009; Rodríguez-Sánchez et al., 2007). Osborne and colleagues (2020) also posited that psychomotor slowing is an endophenotype and biomarker of schizophrenia, and our neuroimaging findings add to this literature.
There are notable differences between our study and other recent rsFC studies using the same publicly available dataset (Bernard et al., 2017; Wang et al., 2014; Wu et al., 2018). Most notably, we computed rsFC from seeds in 20 hemispheric cerebellar lobules, whereas the other studies utilized only two to three ROIs. Bernard and colleagues (2017) focused on right lobule V and Crus I, Wang and colleagues (2014) used spherical seeds placed in Crus I and lobule IX to measure default mode connectivity, while Wu and colleagues (2018) used independent components analysis, resulting in two cerebellar components (bilateral anterior and posterior) that did not correlate with cortical components. Thus, our study is both more exploratory and more complete, in that we investigated lobules that had not been used as ROIs in rsFC previously.
Our results specifically differ from Bernard and colleagues (2017), in that we found connectivity differences in bilateral lobule V, and differences were between lobule V and mainly temporoparietal regions. Thus, our sample slightly differs from Bernard, and we processed our data differently, and our results do not align exactly, but we also see dysconnectivity in schizophrenia between motor lobules and a broad sensorimotor network. Similar to Bernard, cortical regions with which our cerebellar ROIs demonstrated lower connectivity were not located in the primary motor cortex, suggesting that the extended “motor network” dysfunction involves other motor-related processes such as attention, visuospatial integration, and motor prediction/planning.
This study therefore adds to Bernard's evidence for motor network dysfunction in schizophrenia by implicating dysconnectivity between the secondary cerebellar motor (lobules VIIIa/b) and cognitive (lobule VIIb) representations and association cortex, though our results were primarily located in temporoparietal areas responsible for attentional and visuospatial processing rather than prefrontal cortex (Bernard et al., 2017; Cavanna and Trimble, 2006; Krall et al., 2015; Seghier, 2013; Trés and Brucki, 2014). Our findings also suggest that these patterns of dysconnectivity may be specifically related to deficits in processing speed.
In addition, consistent with Bernard's and Wu's work (Bernard et al., 2017; Wu et al., 2018), we did not find correlations between cerebellar rsFC and symptom severity. We also did not observe relationships between the MCCB composite score and cerebellar connectivity, in line with Wu and colleagues' (2018) findings. Therefore, it appears that our approach of investigating MCCB domains may be more sensitive to specific group differences in lobular rsFC and specific cognitive deficits (Sui et al., 2015). Further, the MCCB total scores are highly correlated with IQ, so group IQ differences may obscure any possible findings.
This study is limited by some characteristics of the sample. All patients included in this study had been ill for an average of 14 years and were stable on antipsychotic medication, which can impact brain structure and function, including the cerebellum (Loeber et al., 2002; Smieskova et al., 2009). However, we did not see correlations between medication dosage and connectivity in this sample. Our patient sample also demonstrated greater head motion than controls, which impacts connectivity measures, though we used recommended measures to control for head motion (Power et al., 2014), and included it as a covariate in all analyses.
Neuroimaging of the cerebellum, particularly the inferior lobules, is challenging, as well, and therefore it is possible that our findings are impacted by noise (Schlerf et al., 2014). It is encouraging, though, that our inferior cerebellar findings are supported by group differences in anterior motor lobules, and that they align with the past studies. It is also important to note that we utilized anatomical boundaries for our ROIs, and functional networks may span multiple lobules (Bernard et al., 2012; Buckner et al., 2011; King et al., 2019). This approach may have affected the extent of cerebellocortical connectivity differences we observed; however, we hope that by including all hemispheric lobules we did cover the majority of correlations between cerebellum and cortex. Our analysis is in keeping with previous seed-to-voxel cerebellar connectivity studies (Bernard et al., 2017), but it will be important to use a more data-driven approach to define cerebellar clusters in future work, especially for regions such as Crus I/II that correlate with multiple cognitive domains.
Further, as this study was largely exploratory and we performed many statistical analyses, it is somewhat underpowered, though we corrected for multiple comparisons conservatively. Finally, differences between our study and others using the same data highlight the issues with neuroimaging replicability and reproducibility; however, using large publicly available datasets, such as this one, and standardizing processing procedures, can help ameliorate some of these difficulties.
In sum, we observed general cerebellocortical connectivity reductions in schizophrenia, primarily between primary and secondary motor cerebellar lobules and parietal and temporal cortex and between left lobule VIIb and right prefrontal cortex. Our results point to lobules primarily associated with motor function as having the strongest relationship with diagnosis. In addition, stronger connectivity between these regions was associated with better processing speed performance in the control group, but these relationships were not present in the schizophrenia group.
Our results align with previous work that has also implicated dysfunctional cerebellar motor and somatosensory networks in schizophrenia (Bernard and Mittal, 2015; Bernard et al., 2017; Collin et al., 2011; Moussa-Tooks et al., 2018), and partially support the cognitive dysmetria theory (Andreasen and Pierson, 2008; Andreasen et al., 1998). While this study cannot determine if cerebellocortical connectivity is causing processing speed deficits, we speculate that dysfunction of cerebellocortical circuits in schizophrenia may contribute to attention, procedural learning, timing, prediction, and/or motor coordination deficits that may then impair performance on processing speed tasks. These results suggest potentially fruitful avenues for future research. Many neuroimaging studies do not fully image lobules VIIIa/b, and none have specifically investigated them in schizophrenia, but it may be an important area to consider in schizophrenia pathogenesis.
Footnotes
Authors' Contributions
S.V.C. performed data analysis and prepared the article. A.T. assisted with data analysis and prepared the article. V.D.C. is the principal investigator on the original COBRE grants and edited the article. J.A.B. created the masks used in analysis and edited the article. J.B. collected the data and edited the article. J.A.T. advised S.V.C. and oversaw data analysis and article preparation.
Acknowledgments
Data were downloaded from the COllaborative Informatics and Neuroimaging Suite Data Exchange tool, and data collection was performed at the Mind Research Network, and funded by a Center of Biomedical Research Excellence (COBRE) grant 5P20RR021938/P20GM103472 from the NIH to Dr. Vince Calhoun. Data used in preparation of this article were obtained from the SchizConnect database. As such, the investigators within SchizConnect contributed to the design and implementation of SchizConnect and/or provided data but did not participate in analysis or writing of this report. Data collection and sharing for this project was funded by NIMH cooperative agreement 1U01MH097435.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Data collection was funded by the Center of Biomedical Research Excellence (COBRE) grant 5P20RR021938/P20GM103472 from the NIH to Dr. Vince Calhoun. Data collection and sharing for this project was funded by NIMH cooperative agreement 1U01MH097435. This project was partially funded by Georgia State University Neurogenomics and Brains and Behavior Fellowships to Sarah V. Clark.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.