
Abstract
Aim:
To determine whether Neanderthal-derived genetic variation relates to functional connectivity patterns in the brains of living modern humans.
Introduction:
Nearly 50,000 years ago, Neanderthals interbred with ancestors of modern humans, imparting a genetic legacy that lives on today. The vestiges of this Neanderthal-derived genetic variation have been previously shown to be enriched in genes coding for neurogenesis and myelination and to alter skull shape and brain structure in living people.
Materials and Methods:
Using two independent cohorts totaling 553 healthy individuals, we employed multivariate distance matrix regression (MDMR) to determine whether any brain areas exhibited whole-brain functional connectivity patterns that significantly related to the degree of Neanderthal introgression. Identified clusters were then used as regions of interest in follow-up seed-based functional connectivity analyses to determine the connectivity patterns driving the relationships.
Results:
The MDMR analysis revealed that the percentage of Neanderthal-originating polymorphisms was significantly associated with the functional connectivity patterns of an area of the intraparietal sulcus (IPS) that was nearly identical in both cohorts. Using these IPS clusters as regions of interest in seed-based connectivity analyses, we found, again in both cohorts, that individuals with a higher proportion of Neanderthal-derived genetic variation showed increased IPS functional connectivity with visual processing regions, but decreased IPS connectivity with regions underlying social cognition.
Conclusions:
These findings demonstrate that the remnants of Neanderthal admixture continue to influence human brain function today, in ways that are consistent with anthropological conceptualizations of Neanderthal phenotypes, including the possibility that Neanderthals may have depended upon visual processing capabilities at the expense of social cognition, and this may have contributed to the extinction of this species through reduced cultural maintenance and inability to cope with fluctuating resources. This and other studies capitalizing on the emerging science surrounding ancient DNA provide a window through which to view an ancient lineage long past.
Impact statement
Few biological enigmas have captivated the contemporary popular mind as much as our distant evolutionary relatives and the nature of the brain, the organ from which all our experiences and volitions arise. The results reported here indicate that gene flow from Neanderthals to modern humans is not just of evolutionary interest, but also influences the function of the living Homo sapiens brain in ways that are consistent with anthropological conceptualizations of Neanderthal culture. These findings, thus, provide a lens through which to view our distant evolutionary relatives who continue to live on as a flicker in our genetic make-up.
Introduction
Approximately 40,000
Some of this genetic inheritance may have been beneficial to modern humans, for example, affecting genes involved in keratin filaments, which may have modified skin and hair to adapt to non-African climates (Sankararaman et al., 2014; Vernot and Akey, 2014). However, other facets of these archaic genotypes have also been linked to risk for disease, including autoimmune disorders such as lupus, biliary cirrhosis, and Crohn's disease (Sankararaman et al., 2014). Perhaps most surprising, however, is that Neanderthal-derived genetic variation is enriched for associations with neuropsychiatric phenotypes and with gene expression in the cerebellum and cerebral cortex, suggesting that there are traces of this ancient DNA that shape the modern human brain (Simonti et al., 2016).
Our prior study supports the notion that Neanderthal admixture affects the brains of living humans, showing that individuals harboring a greater proportion of Neanderthal-derived variants have skull shapes that more resemble Neanderthal fossil remains, and that underlying brain morphology is altered as well (Gregory et al., 2017). Importantly, the metric used to define the proportion of Neanderthal-derived variants in this previous study (the same metric used here) was related to morphological features of the skulls of living humans in ways that closely recapitulated known features of fossilized Neanderthal skulls, thus providing crucial validation for this measure and enabling further examinations with other phenotypes for which anthropological data are not available. The structural brain changes previously found to be associated with Neanderthal introgression were located in brain regions that are important for visual processing, including a more developed intraparietal sulcus (IPS), suggesting that Neanderthal brains might have been preferentially wired for visual functions and that the vestiges of this admixture contribute to brain development in modern humans. These findings have been subsequently supported by computational modeling from fossil remains, showing putatively expanded occipital regions in Neanderthal brains (Kochiyama et al., 2018), and are highly consistent with anthropology literature examining skull shapes (Ponce de Leon and Zollikofer, 2001). In addition, introgressed Neanderthal variation has been associated with reduced endocranial globularity, and the lead single nucleotide polymorphisms (SNPs) driving these changes affect neural expression of genes important for neurogenesis and myelination (Gunz et al., 2019). Further, Neanderthal, but not Denisovan admixture, is enriched in genes associated with neurogenesis in Papauans, a population in whom both archaic genotypes are represented (Akkuratov et al., 2018). Those data offer a potential neurogenetic basis for the Neanderthal-related brain structural findings cited above and suggest that there also could be repercussions of the residua of this ancient genetic signature reflected in the characteristics of how modern human brains function. Despite the impetus collectively provided by these studies, there has been no direct search for such neurofunctional sequelae of this archaic inheritance in living persons. In this study, in two separate healthy adult cohorts of European ancestry (total n = 553), we carried out a whole-brain search for regions in which cross-regional cooperativity (i.e., “functional connectivity”) patterns were significantly related to the degree of Neanderthal admixture.
Materials and Methods
Participants
Two independent samples were used to study functional connectivity patterns. In the National Institute of Mental Health (NIMH) Discovery sample, 304 participants (mean age = 30.9 ± 9.45 years, 157 women) were identified from a pool of healthy control volunteers recruited as part of the NIMH Sibling Study—a multidisciplinary initiative aimed at examining the neurobiology of schizophrenia risk—who had completed both neuroimaging with 3-Tesla magnetic resonance imaging (MRI) and genome-wide SNP genotyping. Participants included in this study were all adults of self-reported European descent and were screened to be healthy and free of any neurological or psychiatric diseases based on clinician-obtained history and physical examination, semistructured diagnostic interview (Structured Clinical Interview for DSM-IV), clinical MRI of the brain, and routine laboratory testing, including urine toxicology. All procedures were carried out in accordance with National Institutes of Health (NIH) guidelines and were approved by the NIH CNS Institutional Review Board, and all participants provided written informed consent for the study procedures. Data were collected under protocols 00M0085/NCT00004571 and 95M0150/NCT00001486.
A replication sample was taken from the publicly available Human Connectome Project (HCP) S1200 Release (Van Essen et al., 2012). Although this release contains 1113 data sets, we only included one participant per family with high-quality imaging and genotyping. After this filtering, 249 unrelated individuals of European ancestry (mean age = 28.7 ± 3.67 years, 118 women) were included.
Genetic analyses
For the NIMH Discovery sample, details of genotyping, imputation (Gregory et al., 2019a), and calculation of a “NeanderScore” metric estimating the variation in Neanderthal-derived genetic variation across participants were described previously (Gregory et al., 2017). In brief, SNP-based genetic information was carried out on blood lymphocytes with Illumina genome-wide SNP chips (550K-2.5M SNPs). Data were phased using Shapeit and imputed using Impute2 software. For the HCP Replication sample, the imputed genotype data were obtained from dbgap (accession no. phs001364.v1.p1). For both the discovery and replication samples, only unrelated individuals of European descent were used in further analyses. Ancestry-related components were calculated from the SNP data using the “pca” function in Plink (version 1.9) for each sample separately. To estimate the percentage of Neanderthal-derived genetic variation in each person, an enhanced D statistic was calculated for each individual (termed “NeanderScore”), which is defined as:
where ∑(n ABBA ) is the count of SNP locations at which the genotyped individual shares an allele with the Neanderthal sequence but differs from all Yoruba and primate sequences, and ∑(n AABA ) is the count of locations at which the genotyped individual shares an allele with Yoruba and primate sequences but differs from the Neanderthal sequence (Fig. 1).
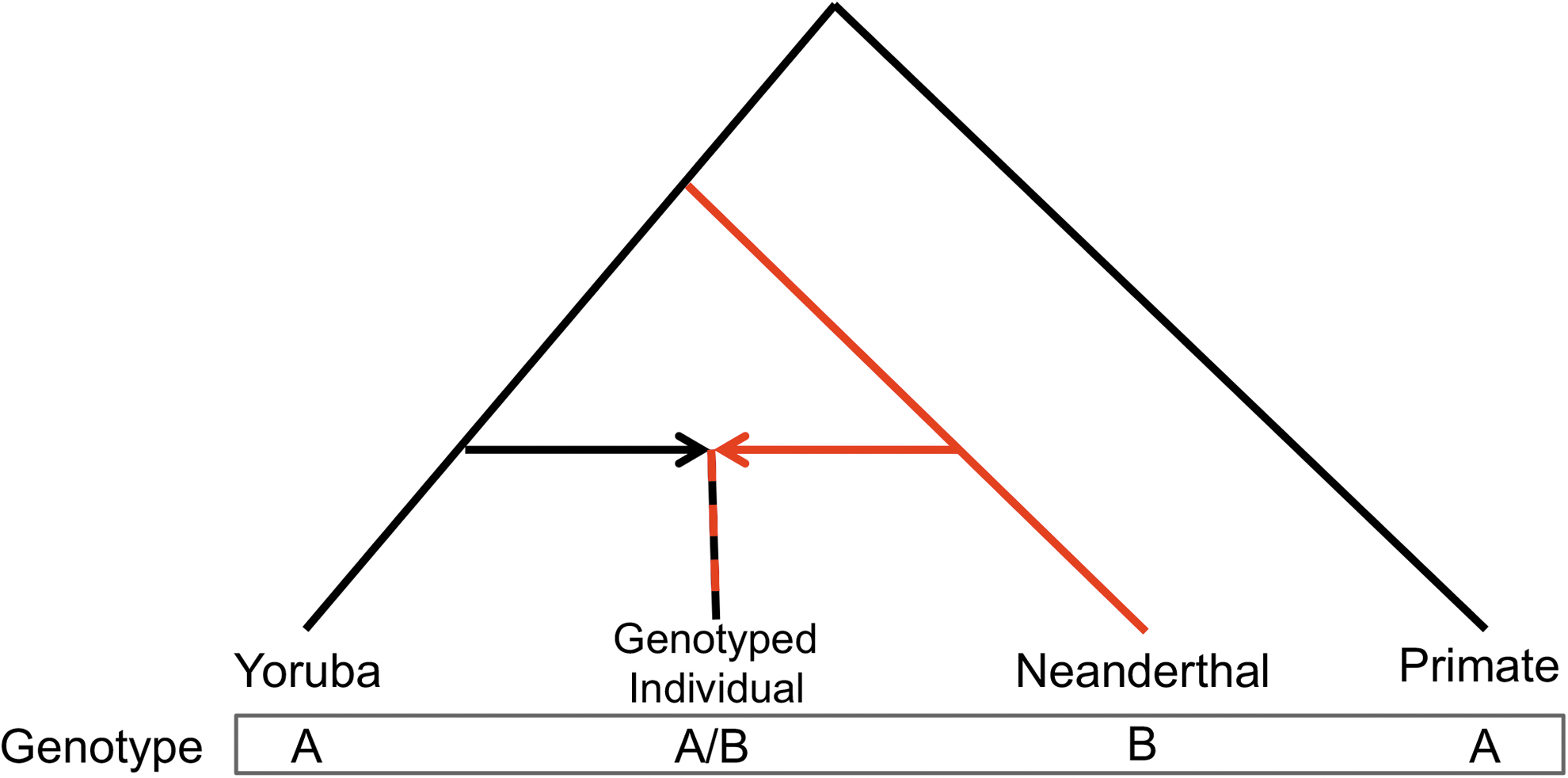
Derivation of the NeanderScore metric. Phylogenetic tree demonstrating the relationship between Yorubans, genotyped individuals of European ancestry, Neanderthals, and primates. For each SNP used in the NeanderScore metric, sites were identified where Yorubans and primates share the ancestral allele (A) and only the derived allele (B) is found in the Neanderthal genome. For each individual genotyped in this study, either the ancestral allele shared by Yorubans and primates (A) or the Neanderthal allele derived through admixture (B) might be found. This SNP could be either AABA or ABBA (reading the bottom genotype line left to right) for a given individual. The percentage of Neanderthal-derived genetic variation, an enhanced D statistic, was calculated for each individual as per the equation in the text. SNP, single nucleotide polymorphism.
Image acquisition and preprocessing
For the NIMH Discovery cohort, functional MRI (fMRI) data were collected on all participants while they performed four cognitive tasks. Acquisition parameters for all tasks were identical (repetition time = 2.0 sec, echo time = 28 msec, flip angle 90°, 3.75 × 3.75 × 6 mm voxels; 26 min, 8 sec total minutes of scanning). Available tasks with these imaging parameters included a flanker task (Sambataro et al., 2013), a facial emotion matching task (Hariri et al., 2002), and encoding and retrieval of neutral or aversive scenes (Murty et al., 2009). A T1-based structural image was acquired for coregistration purposes. “Pseudo-resting” time series were created for each participant by concatenating all task scans and regressing out the relevant task designs, an analytic procedure that has been demonstrated to produce time series comparable with resting-state scans (Cole et al., 2014; Sheffield et al., 2015). Images for each task were coregistered to each individual's structural image, motion corrected, and normalized to Montreal Neurological Institute standardized space separately. Artifact detection tool (ART) was used to exclude time points with a global signal >3 standard deviations from the mean across all time points, or an absolute movement between consecutive images >0.5 mm (Whitfield-Gabrieli and Nieto-Castanon, 2012). Using the 3dTproject function of AFNI, data were concatenated, bandpass filtered (0.008–0.1 Hz), and spatially smoothed [6 mm full width at half max (FWHM)], and effects of no interest were regressed out (including the six-direction motion parameters, the first five anatomic CompCor components from a principle components analysis of an eroded white matter/cerebrospinal fluid mask, and task design) in a single step, resulting in a pseudo-resting time series for each participant. Of note, anatomic CompCor (Behzadi et al., 2007) has been shown to appropriately remove nuisance signals from fMRI data (Caballero-Gaudes and Reynolds, 2017) and attenuates the relationship between motion and MR signal, improving connectivity metrics (Muschelli et al., 2014; Goto et al., 2016).
For the HCP Replication data set, the “minimally processed” resting-state data were used for functional processing and a T1-based structural image was used for coregistration purposes. Preprocessing was similar to that applied to the NIMH Discovery sample, where ART was used to exclude corrupted time points and data were concatenated, bandpass filtered (0.008–0.1 Hz), spatially smoothed (6 mm FWHM), and effects of no interest were regressed out (including those related to motion and anatomic CompCor components) in a single step.
Imaging analyses
A connectome-wide association study was performed using the “connectir” package for R (Shehzad et al., 2014) in each data set separately. This method identifies brain regions where whole-brain functional connectivity patterns are significantly related to a phenotype of interest (in this case, NeanderScore). In brief, this method first assesses subject-level functional connectivity by comparing the time course of each voxel with all other voxels in a gray matter mask. Then, for each voxel in the gray matter mask, the connectivity maps were compared across participants for each pair-wise comparison, creating a distance matrix of participant's connectivity. Then, multivariate distance matrix regression (MDMR) was used to test whether, for each voxel, individual variation in whole-brain connectivity patterns significantly related to NeanderScore, controlling for age, gender, and the first five ancestry-related genetics principal components from genetic information. MDMR calculates a pseudo-F statistic for each voxel to explain whether the between-subject distances of whole-brain connectivity maps are related to a variable of interest. This is analogous to a standard analysis of variance (ANOVA) model but does not have an F-distribution, therefore, p-values and Z-scores are determined by randomly permuting the subject indices and recomputing the pseudo-F statistic for each voxel. MDMR analyses were performed on the Biowulf High Performance Supercomputing Cluster at NIH (
As the MDMR measure provides a statistical relationship of a voxel's connectivity patterns to the NeanderScore metric, it does not characterize the directionality or regional specificity driving the result. Following prior study that has used post hoc seed-based functional connectivity analyses as post hoc characterizations of the results (Sharma et al., 2017; Satterthwaite et al., 2015; Satterthwaite et al., 2016), clusters identified in the MDMR analysis were carried forward as seed regions in a typical functional connectivity analysis. For this analysis, using AFNI, functional connectivity Z-score maps were created for each participant to determine the strength of the correlation between the signal time course of the significant cluster with the time course of each voxel in the brain. Voxel-wise linear regression then tested for associations across participants with NeanderScore, controlling for age, gender and the first five ancestry-related components in each sample separately. Statistical results were converted to Z-scores and combined in a meta-analytic manner using Stouffer's Z-score method (Stouffer, 1949). Results were thresholded at p < 0.05, family-wise error corrected for multiple comparisons, using 3dClustSim to compute a cluster threshold based on 100,000 Monte Carlo simulations of synthesized white Gaussian noise, taking into account the smoothing and resampling parameters of the functional connectivity analyses using the auto-correlation function method in AFNI (Cox et al., 2017).
Results
Whole-brain functional connectivity patterns that are related to NeanderScore
In the NIMH Discovery sample, we collected genetic information along with 26 min and 8 sec of fMRI neuroimaging data, which were combined into “pseudoresting” time series (see methods) (Cole et al., 2014; Sheffield et al., 2015) for 304 individuals (mean age = 30.9 ± 9.45 years, 157 women). After genetic imputation, we calculated a measure of the percentage of Neanderthal-derived genetic variation (henceforth termed “NeanderScore”) for each individual. MDMR of functional connectivity patterns was performed on fMRI data using the “connectir” R package (Shehzad et al., 2014) to determine voxels where whole-brain functional connectivity patterns significantly related to NeanderScore, controlling for age, gender, and ancestry-related components. This analysis identified one significant cluster in the left IPS (Fig. 2a; peak MNIx,y,z = −30,−85,29; Z = 3.1; p = 9 × 10−4).

Brain regions where NeanderScore was related to whole-brain functional connectivity. Posterior view of the brain highlighted in magenta the IPS voxels with whole-brain functional connectivity patterns related to NeanderScore in two independent samples:
In a replication sample, the same analysis was performed using data from the publicly available HCP S1200 release (Van Essen et al., 2012). In this study, 28 min and 48 sec of resting-state fMRI data from 249 unrelated adults (mean age = 28.7 ± 3.67 years, 118 women) were analyzed. In this confirmatory analysis, MDMR also identified a cluster of voxels in the left IPS in which functional connectivity with the rest of the brain was significantly related to NeanderScore, also controlling for age, gender, and ancestry-related components (Fig. 2b; peak MNIx,y,z = −29,−78,36; Z = 4.1; p = 2 × 10−5). Importantly, this cluster's anatomical locale was nearly identical to that identified in the discovery sample.
IPS functional connectivity patterns that drive the association with NeanderScore
Although the MDMR analytic method provides a single voxel-wise measure of the robustness of each voxel's whole-brain functional connectivity pattern with NeanderScore, this metric reveals neither the directionality nor the regional distribution of the IPS functional connectivity driving this result. Therefore, to further characterize this finding, we next used the identified IPS clusters as seed regions in follow-up functional connectivity analyses using NeanderScore as a regressor of interest, also controlling for age, gender, and ancestry-related components, separately in each sample. Statistical maps for both samples were converted to Z-scores and combined using Stouffer's Z-score method (Stouffer, 1949) for reporting purposes (Fig. 3). In both the discovery and replication samples, these follow-up connectivity analyses showed that IPS functional connectivity with known visual processing regions, including occipital cortex and fusiform gyrus, as well as with bilateral insulae, was significantly positively related to NeanderScore. In contrast, IPS connectivity with regions known to be part of the default network, involved in social processing (Spreng and Andrews-Hanna, 2015), including posterior cingulate cortex/precuneus, temporoparietal junction, prefrontal cortex, and superior frontal gyrus, was found to be significantly negatively related to NeanderScore (Table 1). Statistical maps for each cohort separately are included as Supplementary Figure S1.
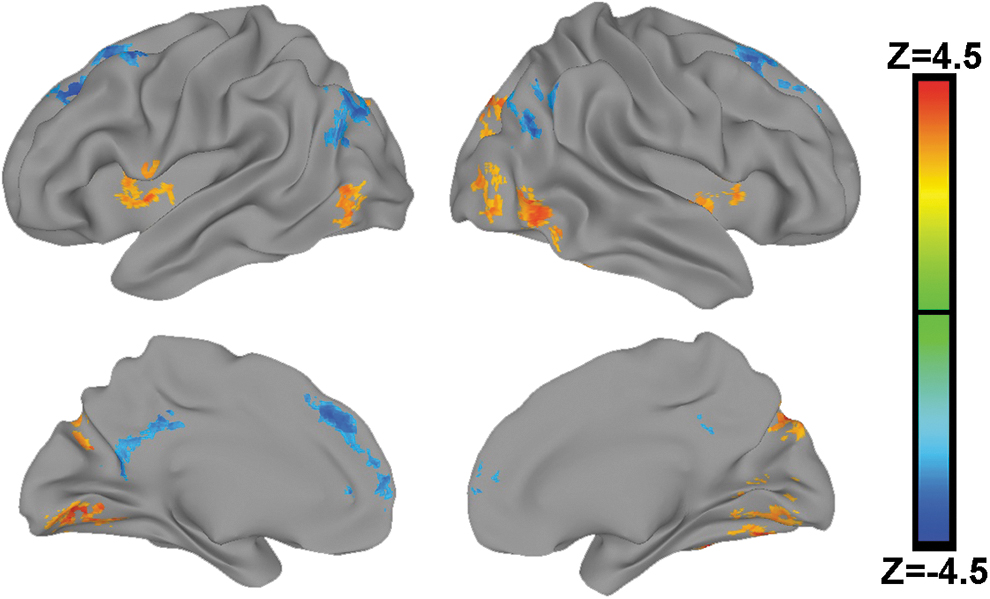
Brain regions where NeanderScore was associated with IPS functional connectivity using the IPS clusters identified in the MDMR analysis (shown in Fig. 2) as seed regions. Meta-analysis showing combined statistics from both the NIMH Discovery sample and HCP Replication sample, thresholded at p < 0.05, FWE corrected and projected onto the brain surface. Regions in orange exhibit a positive relationship wherein increasing NeanderScore was associated with greater IPS functional connectivity and regions in blue exhibit a negative relationship. Note that regions with significantly negative functional connectivity with the IPS predominantly include areas associated within the brain's default network, involved in mentalizing and social processes, whereas the orange regions showing significantly positive functional connectivity with the IPS predominantly include areas involved in visual processing. FEW, family-wise-error; MDMR, multivariate distance matrix regression.
Brain Regions Where NeanderScore Was Associated with Intraparietal Sulcus Functional Connectivity in the Combined Sample
MNI, Montreal Neurological Institute standardized space.
Discussion
Although we and others have previously shown that Neanderthal-derived genetic variation affects the shape of skull and brain, no prior studies have examined the neurofunctional effects of this inheritance. In this study, we show that the degree of Neanderthal admixture significantly relates to the functional connectivity patterns of the IPS in the human brain in vivo. This finding was specific to the IPS, was not observed in other brain regions, and was replicated in an independent sample. Further, the relationship was driven by increased IPS connectivity to visual networks and decreased connectivity to social networks as NeanderScore increased.
The IPS region identified here is remarkably similar to the cortical area previously found to have altered brain shape using the same NeanderScore metric (Gregory et al., 2017), although contralaterally. Moreover, this region directly underlies the area where skull morphology was also significantly related to Neanderthal introgression (Gregory et al., 2017), a result supported by fossil remains (Ponce de Leon and Zollikofer, 2001). The anatomical consistency of the findings across multiple neuroimaging modalities and brain phenotypes reinforces their validity. The localization of the findings to the IPS is also of special interest from both evolutionary and neurofunctional perspectives. Not only is this region highly gyrified across the primate order (Zilles et al., 2013), it is also theorized to have been significantly expanded through hominid evolution (Vanduffel et al., 2002). In addition, the IPS is a well-documented neural hub for the processing of visual information and is a critical component of the dorsal visual stream (commonly known as the “where” pathway), which deciphers spatial information from visual stimuli (Ungerleider and Mishkin, 1982). In addition to its role in visual processing, the IPS also plays a central role in tool manipulation (Vingerhoets, 2014), and both Neanderthals and early modern humans were known to fabricate tools more complicated than those used by other species (Ruebens, 2013). In sum, this information supports the idea that evolution and Neanderthal introgression have played an active role in the refinement of modern human IPS structure and function.
The connectivity patterns driving the relationship between IPS functional connectivity and Neanderthal-derived genetic variation deserve particular mention. Our seed-based functional connectivity analyses revealed that the primary IPS–NeanderScore association was driven by increased IPS functional connectivity with regions subserving visual processing, but decreased connectivity with regions implicated in the neurobiology of social processing. Anthropological evidence has sparked the theory that Neanderthals may have depended upon visual processing capabilities at the expense of social cognition, and that the latter of these characteristics may have contributed to the extinction of this species through reduced cultural maintenance and inability to cope with fluctuating resources (Pearce et al., 2013). Our findings of increased IPS functional connectivity with visual networks but decreased connectivity with social networks may provide a neurobiological framework for understanding this theory by suggesting that divergent evolutionary pressures and trajectories resulted in a modern human brain more biased toward making existing Homo sapiens the social beings that we are. The concept of such trade-offs between neural systems is further supported by our recent report that in Williams syndrome, a genetic condition in which increased social drive together with impaired visuospatial abilities are the clinical hallmarks, IPS functional connectivity is weighted oppositely to that associated with NeanderScore (i.e., in Williams syndrome, increased IPS connectivity with socially relevant networks, but decreased IPS connectivity with visually important regions) (Gregory et al., 2019b). While the findings identified here use both resting-state and pseudoresting, “baseline” fMRI data do not necessarily speak of whether Neanderthal introgression impacts the activation of specific brain regions in response to stimuli or specific tasks, they provide a perspective for important future investigations of whether, and, if so, how, task-related activation may be related to Neanderthal introgression.
Conclusion
Taken together, these data offer evidence that Neanderthal-derived genetic variation affects neural system-level cooperative function in the human brain. Although we do not identify specific genetic mechanisms responsible for these changes, and instead take a more polygenic approach, future study may elucidate particular contributions conveyed to us across time from the genes of early lineages. The approaches here, and others that capitalize on the emerging science surrounding ancient DNA, will continue to provide a lens through which to view the physiology and neurobiology of our distant evolutionary relatives who continue to live on as a flicker in our genetic make-up.
Footnotes
Authors' Contributions
M.D.G. and K.F.B. conceived of the project. M.D.G., D.P.E., J.H.C., J.S.K., P.K., B.K., and K.F.B. conducted the experiments. M.D.G. wrote the initial draft and all authors reviewed, edited, and approved the final draft.
Acknowledgments
The data for the NIMH Sample were obtained under protocol 00M0085/NCT00004571 and protocol 95M0150/NCT00001486. Some of this study utilized the computational resources of the NIH HPC Biowulf cluster (
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Intramural Research Program (ZIAMH002942), NIMH, NIH, Bethesda, MD. HCP data were funded, in part, by the HCP (1U54MH091657) from the 16 NIH Institutes and Centers that support the NIH Blueprint for Neuroscience Research, and by the McDonnell Center for Systems Neuroscience at Washington University.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.