
Abstract
Aim:
Structural connectivity in the reorganizing spinal cord after injury dictates functional connectivity and hence the neurological outcome. As magnetic resonance imaging (MRI)-based structural parameters are mostly accessible across spinal cord injury (SCI) patients, we studied MRI-based spinal morphological changes and their relationship to neurological outcome in the rat model of cervical SCI.
Introduction:
Functional connectivity assessments on patients with SCI rely heavily on MRI-based approaches to investigate the complete neural axis (both spinal cord and brain). Hence, underlying MRI-based structural and morphometric changes in the reorganizing spinal cord and their relationship to neurological outcomes is crucial for meaningful interpretation of functional connectivity changes across the neural axis.
Methods:
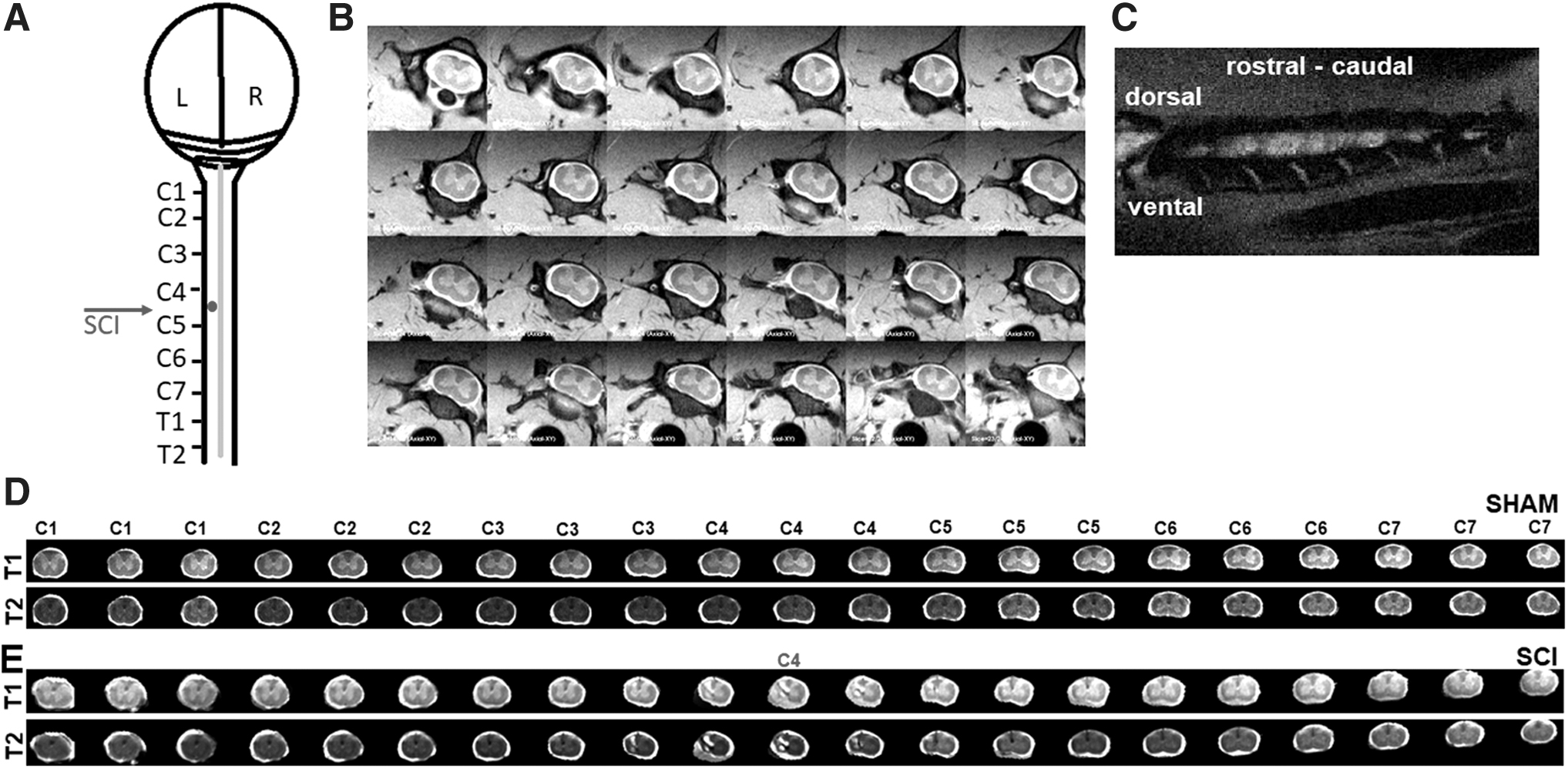
Young adult rats, aged 1.5 months, underwent a precise mechanical impact hemicontusion incomplete cervical SCI at the C4/C5 level, after which sensorimotor behavioral assessments were tracked during the reorganization period of 1–6 weeks, followed by MRI of the cervical spinal cord at 8 weeks after SCI.
Results:
A significant ipsilesional forelimb motor debilitation was observed from 1 to 6 weeks after injury. Heat sensitivity testing (Hargreaves) showed ipsilesional forelimb hypersensitivity at 5 and 6 weeks after SCI. MRI of the cervical spine showed ipsilateral T1- and T2-weighted lesions across all SCI rats compared with no significant lesions in sham rats. Morphometric assessments of the lesional and nonlesional changes showed the diverse nature of their interindividual variability in the SCI receiving rats. While the various T1 and T2 MRI lesional volumes associated weakly or moderately with neurological outcome, the nonlesional spinal morphometric changes associated much more strongly. The results have important implications for interpreting functional MRI-based functional connectivity after SCI by providing vital underlying structural changes and their relative neurological impact.
Impact statement
Functional connectivity assessments on patients with SCI relies heavily upon MRI based approaches. Hence, underlying MRI based structural and morphometric changes in the reorganizing spinal cord and its relationship to neurological outcomes is vital for meaningful interpretation of functional connectivity changes across the complete neural axis (both spinal cord and the brain).
Introduction
Magnetic Resonance Imaging (MRI) visualization in spinal cord injury (SCI) trauma victims has been helpful in improving patient outcomes (Freund et al., 2019). Secondary injury is the result of molecular and cellular mechanisms that are activated by the primary traumatic event and which lead to neuronal, axonal, and glial dysfunction and loss. Secondary injury mechanisms, which in humans are initiated hours following the initial mechanical damage and continue for months, include inflammation, glial activation, glutamate-induced excitotoxicity, oxidative stress, and mitochondrial damage (Ahuja et al., 2017; Alizadeh and Karimi-Abdolrezaee, 2016; Lipinski et al., 2015; Tator, 1995). Secondary injuries due to spinal contusions may sometimes manifest as lesions, which are generally visualized by conventional T1- and T2-weighted anatomical MRI (Endo et al., 2007; Freund et al., 2019; Lammertse et al., 2007). In the acute phase after SCI (few hours to day 1 or day 2 in humans), hemorrhages appear as T1 or T2 hypointensities, whereas tissue edema appears as T1 or T2 hyperintensities. However, during the chronic phase (weeks to months in humans), atrophic tissue appears as T1 or T2 hypointensity and neural/neurovascular dysfunctions as T1 or T2 hyperintensity (Sundberg et al., 2010; Weirich et al., 1990). Most diffuse injuries at the microstructural levels, representing axonal damage/dysfunction and decreased myelin densities, often do not show up as overt lesions in conventional T1- or T2-weighted MRI and require specialized diffusion tensor imaging (DTI) to be quantified (Deo et al., 2006; Seif et al., 2020; Sundberg et al., 2010). Diffuse injuries, remote from the SCI epicenter, tracked by DTI studies have been valuable predictors of clinical neurological outcomes in cervical SCI patients (Azzarito et al., 2020) and pre-clinical animal models (Deo et al., 2006; Sundberg et al., 2010; Wilkins et al., 2020). The nonlesional (diffuse) injuries, which evolve as parenchymal volume loss in conventional MRI, have not received much attention (Azzarito et al., 2020).
Electrophysiologic and blood oxygen level-dependent functional MRI (fMRI)-based functional connectivity mapping (i.e., mapping of neural circuits either spontaneously active in the absence of any stimulus or synchronously responding to an external task or stimulus) has been widely used to study neuroplasticity in the brain (Athanasiou et al., 2017; Endo et al., 2008; Hofstetter et al., 2003; Matsubayashi et al., 2018) and spinal cord (Eippert et al., 2017; Powers et al., 2018) in both humans and animal models. While functional connectivity studies have been impactful in translating the neuroplastic changes during reorganization after an SCI, the underlying MRI structural studies in the spinal cord mostly focus on lesions. Hence, there is a current gap in knowledge on the relationship between the different MRI-based spinal lesion volumes, spinal parenchymal volume loss, and neurological outcomes, a vital component to integrate the specialized DTI-based microstructural and fMRI-based functional connectivity changes across the spinal cord and their association with the overall neurological outcome.
The clinical relevance of this study was to inform the chronic lesional and nonlesional morphometric changes in the spinal cord that associate with the neurological debilitations presented by the SCI patient during rehabilitation. Using a rat hemicontusion model of cervical SCI, we tested the associations between nonlesional spinal parenchymal volumetric changes with T1- and T2-weighted MRI lesions and their predictive value on the neurological outcomes. Rats, aged 1.5 months, underwent a cervical SCI at the C4–C5 level, after which motor behavioral assessments were made after 1–6 weeks and sensory behavioral assessments were performed 5 and 6 weeks after SCI. After the behavioral evaluations, rats were imaged postmortem at 8 weeks after SCI/sham surgeries to obtain the T1- and T2-weighted MRI of the cervical spinal cord. Multiple morphometric parameters such as T1- and T2-based hypo- and hyperintensities, ipsilateral, contralateral, total parenchymal volumes, and T2-based ipsilateral–contralateral central axis lengths at the injury epicenter (i/c ratio at the epicenter) were determined and tested for their associations with the sensory and motor behavioral outcomes.
Methods
Surgery and SCI induction
All animal procedures were approved by the Rutgers Biomedical and Health Sciences-Newark and Yale University Institutional Animal Care and Use Committees (IACUCs). These procedures are consistent with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Food (standard chow) and tap water were provided ad libitum during the experiments. Female Sprague Dawley rats, at 1.5 months of age, were randomly assigned to sham (n = 7) and SCI (n = 11) groups. Animals were anesthetized with ketamine/xylazine (100/10 mg/kg, intraperitoneal [i.p.]) and laminectomized unilaterally on the left side at the C4–C5 level (Fig. 1A). Using aseptic techniques, a midline incision was made from the base of the skull to the scapula exposing the dorsal elements of the C2–C5 vertebrae. Using a scalpel, an incision was made through three muscle layers over the spinal column and two cotton-tipped applicators were used in a twisting motion to separate the paraspinal muscles. To obtain a good surgical field for the laminectomy, muscles were retracted with four homemade retractors (Lepore, 2011) and pulled away using a rongeur. Hemilaminectomies of the left C4 and C5 levels were performed using rongeurs. Subsequently, the animal was mounted on the Infinite Horizon (IH) impactor device (Precision Systems and Instrumentation, LLC, Lexington, KY). The impactor tip (2.5 mm in diameter) was lowered and aimed using the vertical adjustment knob and the two horizontal adjustment knobs on the IH impactor until the center of the impactor tip hovered 1 mm to the left of the apex of the C4–C5 midline spinous processes. The vertical adjustment knob was turned 3.5 turns to raise the tip 7 mm above the dura, and a 150 kDyne force setting was utilized to perform a moderate mechanical-impact contusion injury. Sham animals underwent identical hemilaminectomies without the impact injury. After injury, muscle layers were sutured using absorbable sutures (5-0) and the skin incisions were closed using wound clips. Animals were provided 2 mL saline i.p. to prevent dehydration and Buprenorphine SR (0.03 mg/kg, subcutaneous) for analgesia immediately after the surgery. Animals were monitored twice a day during the first 48 h after the surgery and daily thereafter until 6 weeks. All surgeries and the immediate postoperative care, including all behavioral testing, were performed at Rutgers Biomedical and Health Sciences. Animals were transferred to Yale University at the end of 6 weeks after SCI. After a quarantine of 2 weeks to ascertain the health of the animals and fulfilling institutional quarantine procedures, MRI experiments were performed 8 weeks after the index surgeries.
Sensorimotor behavior
Forelimb utilization test
The forelimb usage test was a modified version of the cylinder test, which used a transparent plastic cubical container of 10 inch sides and height, providing a manner to evaluate spontaneous forelimb usage behavior (Chitturi et al., 2018; Gensel et al., 2006). The forelimb utilization test is a valid measure for the current hemicontusion SCI model, exhibiting behavioral deficits across the forelimbs, which is also asymmetric due to the injury to one hemisphere of the spinal cord. Each animal was observed for 5 min of duration in a test cage to establish a baseline 5 days before the index sham or SCI surgeries and at 1, 2, 4, and 6 weeks after the surgical procedures. Active exploration of the vertical surfaces by rearing up on their hindlimbs and wall surface exploration with their forelimbs were observed and scored. The number of independent wall placements observed for contralateral, ipsilateral, or both forelimbs was scored. The contralateral, ipsilateral, or both forelimb usage as a percentage of all forelimb usage within the 5 min of observation duration was determined.
Plantar heat sensitivity test (Hargreaves test)
The Hargreaves test was carried out at 5 and 6 weeks after SCI, respectively. Tests were performed at the same time of the day for all testing sessions. Animal subjects were randomly altered within and across the testing sessions. Rats were habituated for 15 min to restrainers placed on a glass surface (IITC Life Science, Woodland Hills, CA). A beam of light with intensity of 30 was delivered to the plantar surface of the forepaws and hindpaws, separately, with modifications from our earlier procedure used in mice (Khariv et al., 2013, 2017) and optimized for rats based on earlier SCI studies in the current rat model (Detloff et al., 2013; Putatunda et al., 2014). The paw withdrawal latency was recorded three times for each forepaw and hindpaw with the final value being the mean of the respective paws. The cutoff time for observation was set to 20 sec to avoid tissue damage.
Magnetic resonance imaging
MRI of the spinal cord was performed using a 9.4T Bruker horizontal bore spectrometer with a home-built 1H surface coil radiofrequency probe (5 × 3 cm diameter). To minimize motion artifacts during MRI, animals were euthanized with 5% isoflurane until respiration ceased and they were then positioned within the radiofrequency coil. No significant changes in conventional T1 and T2 images in MRI have been observed between live and ex vivo spinal cord samples in pre-clinical animal model studies (Scholtes et al., 2008). To minimize motion artifacts and preserve MRI outcomes that are comparable to live animals, the present protocol acquired the MRI acquisitions immediately after euthanasia in the intact animal. Rapid acquisition relaxation-enhanced (RARE) and fast spin echo-RARE (TurboRARE) pulse sequences were used for the acquisition of both T1- and T2-weighted images, respectively. Axial spin echo images of the spinal cord were acquired with Bruker ParaVision 6.1 software using both T1- and T2-weighted protocols. RARE T1-weighted sequence parameters are as follows: repetition time/echo time (TR/TE) = 4000/5.8 msec, image matrix = 256 × 256, field of view (FOV) = 28 mm, and number of excitations (NEX) = 2. For the T2 TurboRARE, TR/TE = 6000/33 msec, image matrix = 256 × 256, FOV = 28 mm, number of echoes = 8, and NEX = 2 were used. Twenty-four axial MRI slices of 1 mm thickness, from the brainstem rostral to the C1 to the T1 vertebral level, were covered by the slices (Fig. 1B).
Image processing, data analysis, and statistics
Acquisition of 24 slices in the MRI acquisition protocol assured that the cervical spinal areas of interest for the current study (i.e., C1 to C7 levels) were always covered in every animal and not lost due to animal positioning differences within the coil. Using the vertebral anatomy as identifiers, 21 axial MRI slices, from the C1 to C7 vertebral levels, were identified across each animal and considered for further analysis. After manually masking the spinal parenchymal and subarachnoid spaces in the 21 images (covering C1–C7) from all animal subjects, 7 shams and 11 SCI, they were linearly registered to a typical T1-weighted spinal cord from a single sham animal using Analysis of Functional Neuroimages (AFNI) software (Cox, 1996). Choosing a specific region of interest from the gray and white matter, the T1- and T2-weighted intensities were inspected in every sham and SCI animal and normalized to the values of the sham animal chosen for the spatial registration. The intensity threshold value larger than the white matter level was used to automatically mask the hyperintense lesions, and the intensity threshold value smaller than the gray matter level was used to automatically mask the hypointense regions in every animal. Using the automatically generated hyper- and hypointensity masks from each animal as a template, the lesions were further inspected and fine-tuned by manual contouring by one of the authors (an MRI researcher trained in digital image processing strategies in both structural and functional brain image analyses in humans and animal models with over 19 years of experience). The manual contouring step was required to eliminate certain false-positive clusters (e.g., the midline arteries) in the automatically generated intensity threshold masks. Contralateral and ipsilateral masks were manually drawn to estimate the spinal cord parenchymal volumes. The injury epicenter was considered at the central MRI slice caudal to the C4 vertebral level, where the hemilaminectomy and the injuries were performed, and verified in each individual SCI subject (Supplementary Fig. S1). Statistical analysis was performed using analysis of variance (ANOVA) with repeated measures or two-tailed t-test with p < 0.05 required for significance. Association within the MRI morphometric variables and associations between morphometric and behavioral variables were determined by linear regression and correlation. The Pearson correlation coefficient value (R) was used as an estimate of the association strength, and the significance of the correlation was tested by ANOVA. Correlations with significance p < 0.05 were deemed strong, p > 0.05 but <0.25 as moderate, and p > 0.25 as low.
Results
General behavioral observation
Animals showed no bladder dysfunction or any respiratory distress at any time after injury. SCI led to immobility with bilateral forelimb and hindlimb debilitation during the first 72 h after SCI. Thereafter, partial mobility using both hindlimbs was observed with a relatively greater use of the contralateral hindlimb. By 96 h after SCI, the rats gained little locomotion across all limbs, except the ipsilateral forelimb. All injured animals demonstrated a clenched ipsilateral forepaw during locomotion until 5 weeks after injury, and by 5–6 weeks after injury, animals started intermittently to unclench the paw. These early behavioral indicators were in agreement with prior studies in this hemicontusion model at the cervical C4–C5 level (Ferguson et al., 2013; Khaing et al., 2012).
Sensorimotor behavior
Preoperatively, all animals used both forelimbs simultaneously 80–90% of the time to explore the cylinder walls (Fig. 2A). There was no specific overall preference to use either limb independently (left/ipsi ∼8%; right/contra ∼6%) (Fig. 2B, C). The limb preference of sham animals did not differ from baseline: used both forelimbs simultaneously 80–90% before and after the surgery (Fig. 2A) and no significant overall preference to use either limb independently (Fig. 2B, C). However, injured animals had a significant preference for the contralateral limb compared with sham animals at all times after SCI (Fig. 2C). By 6 weeks, injured animals significantly improved, depending less on the contralateral forelimb for vertical exploration (p < 0.0009; for week 1 vs. week 6). The improvement or reduction in contralateral limb usage was completely attributable to improved simultaneous use of both forelimbs (Fig. 2A) as opposed to increased use of the ipsilateral forelimb alone. Injured animals never used ipsilateral forelimb alone, to explore the cage walls at all times after injury (Fig. 2B). However, the engagement of both forelimbs rather than the contralateral limb to explore the cylinder walls suggested that the ipsilateral limb regained some degree of motor function at 5–6 weeks.
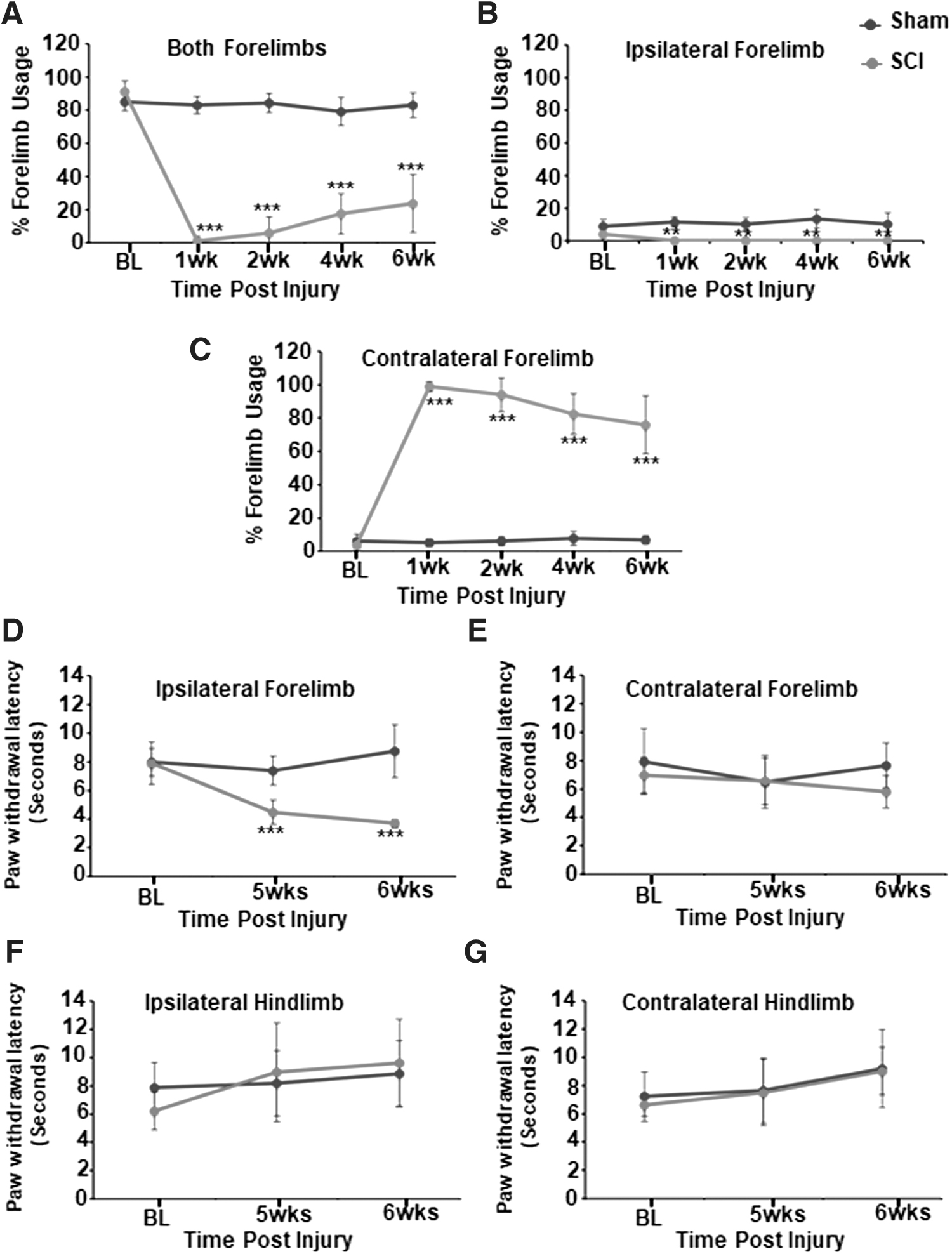
Percentage forelimb use behavior in sham (n = 5) and SCI (n = 7) animals.
The Hargreaves heat sensitivity test was performed at 5 and 6 weeks after injury, by which time, animals start intermittently to unclench the paw, so that the laser beam can be placed on the plantar surface of the forepaw. In the Hargreaves heat sensitivity test, no significant differences were observed between the presurgical baseline and post-sham procedure forelimb or hindlimb behaviors measured at 5 and 6 weeks (Fig. 2D–G). No significant differences in heat sensitivity were observed across the ipsilateral and contralateral hindlimbs (Fig. 2F, G) and contralateral forelimb (Fig. 2E), between sham and SCI. However, heat sensitivity across the ipsilateral forelimb increased significantly at 5 and 6 weeks after SCI, suggesting manifestation of neuropathic pain (Fig. 2D).
Spinal MRI morphometry
A highly accurate and controlled impact was produced in the current SCI procedure (n = 11 animals; impactor displacement = 1.43 ± 0.16 mm; mean ± standard deviation) leading to MRI lesions predominantly localized to the ipsilateral spinal cord (Fig. 1D, E), which was consistent across all SCI animal subjects (Supplementary Fig. S1). From the unique shapes of the laminae at each vertebral level (Fig. 1B), 21 axial slices exactly covering the C1–C7 vertebral levels were identified form all animal subjects and considered for further analyses. Typical T1- and T2-weighted images of the spinal cord from the C1–C7 vertebral levels from a sham animal and an SCI animal are shown in Figure 1D and E, respectively.
Interrelationship between MRI morphometric parameters
Across both T1- and T2-weighted images, lesions were clearly lateralized to the ipsilateral spinal cord (Fig. 3A, B), extending rostrally to C3 and caudally to C5 from the epicenter located at the C4 level (Fig. 3A, B). A similar pattern of ipsilaterally localized lesions was observed across all 11 SCI animal subjects, whereas no lesions were discernable in any of the 7 sham subjects (Supplementary Fig. S1). Multiple morphometric parameters such as T1 hypointensity, T1 hyperintensity, T2 hypointensity, T2 hyperintensity, T1-based ipsilateral, contralateral, and total spinal parenchymal volume (covering the complete cervical spine, C1–C7) and T2-based ipsilateral–contralateral horizontal central axis length ratio at the epicenter (i/c ratio at the epicenter) were measured (Fig. 3C, T2 image). Lesional volume had a between-subject coefficient of variation (CV) of 30% and ipsilateral volume loss had a CV of 35%, which indicated a comparable subject-wise MRI outcome variability. Although the hypointense lesional volumes from T1- or T2-weighted MRI were not highly variable, hyperintense lesional volumes showed larger variability across the SCI animals (Fig. 3D). There were no significant differences between sham ipsilateral and contralateral parenchymal volumes, whereas a significant ipsilateral volume and total parenchymal volume loss was observed across the SCI animals (Fig. 3E). Furthermore, no significant contralateral parenchymal volume difference was observed between sham and SCI animals (Fig. 3E).
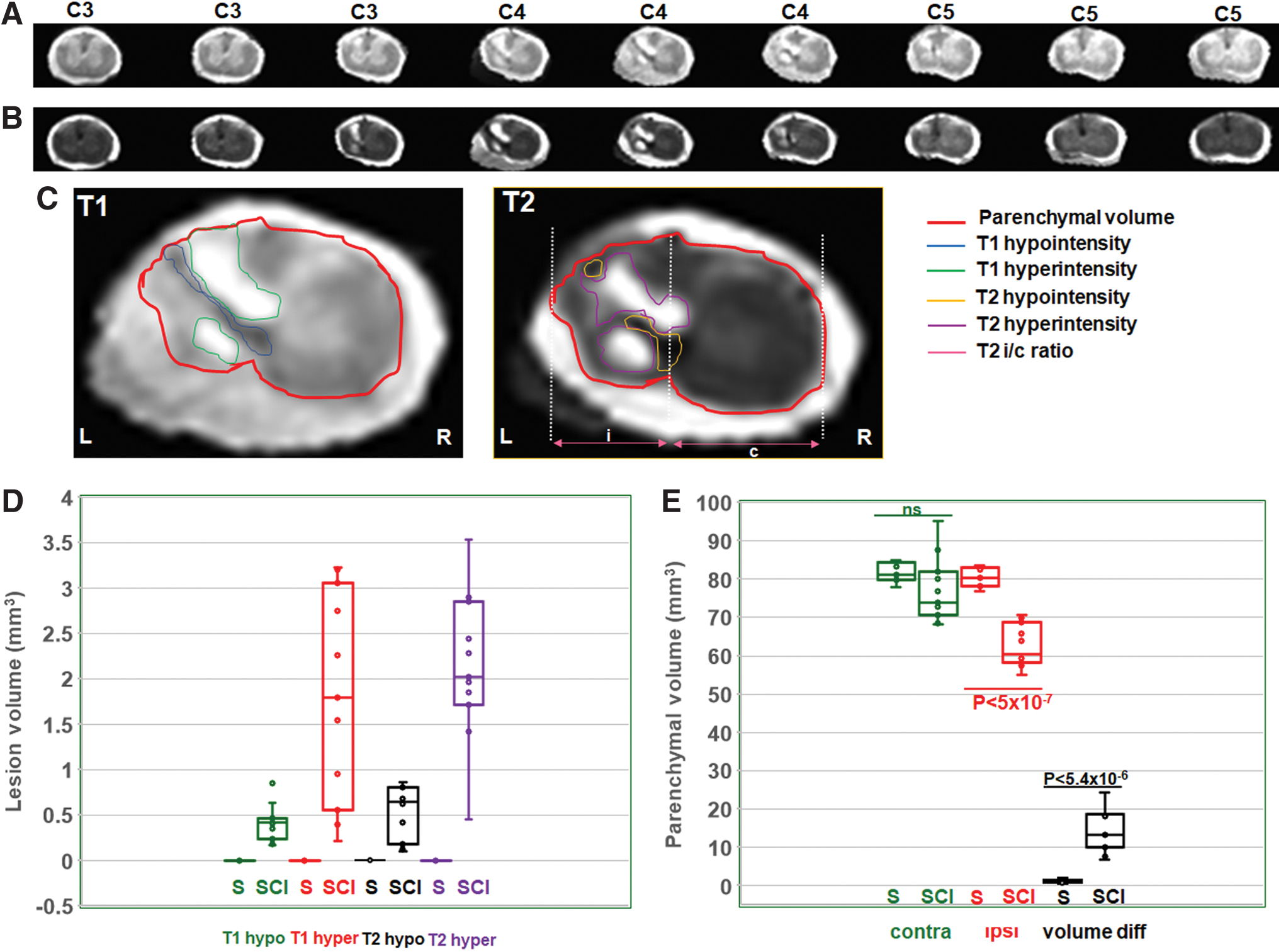
Typical T1- and T2-weighted MRIs along the C3–C5 vertebral columns with the injury epicenter at C4.
Strong correlations were observed between the T2 hypointensity versus the i/c ratio at the epicenter (Fig. 4B; R = 0.65; p < 0.03) and the i/c ratio at the epicenter versus contralateral–ipsilateral volume difference (Fig. 4C; R = 0.62; p < 0.04). Correlations were moderate between T1 hyperintensity versus T1 hypointensity (Fig. 4A; R = 0.51; p < 0.1) and total lesional volume versus contralateral–ipsilateral volume difference (Fig. 4C; R = 0.50; p < 0.11). Moderate correlations were also observed between T2 hypointensity versus contralateral–ipsilateral volume difference (Fig. 4C; R = 0.42; p < 0.2) and total lesional volume versus the i/c ratio at the epicenter (Fig. 4C; R = 0.40; p < 0.22). Low correlations were observed between the other morphometric variable combinations (Fig. 4C). A low correlation between any two morphometric variables indicated unique underlying neuropathological processes, and conversely, a stronger correlation between any two morphometric variables signified an overlap of similar underlying neuropathological processes.

Relationship between various T1- and T2-weighted morphometric parameters across the SCI animals (n = 11).
Association between MRI morphometric parameters and sensorimotor behavior
While the T1- and T2-weighted lesional volumes showed only low or moderate correlations with forelimb motor behavior (Supplementary Table S1), the i/c ratio at the epicenter (Fig. 5A–D) and the contralateral–ipsilateral volume differences (Fig. 5E–H) showed high correlations with forelimb motor behavior from 1 to 6 weeks after SCI. Across the sensory paradigm tested by the heat sensitivity (Hargreaves) test, only the i/c ratio at the epicenter and the contralateral–ipsilateral volume difference showed a moderate correlation with the forepaw withdrawal latency at 5 weeks (Fig. 6A) and 6 weeks after SCI, respectively (Fig. 6C). Other MRI variables did not show any strong association with the forelimb sensory responses except the T1-weighted hyper- and hypointensities, which showed a moderate correlation with paw withdrawal latency at 6 weeks after SCI (Fig. 6D).
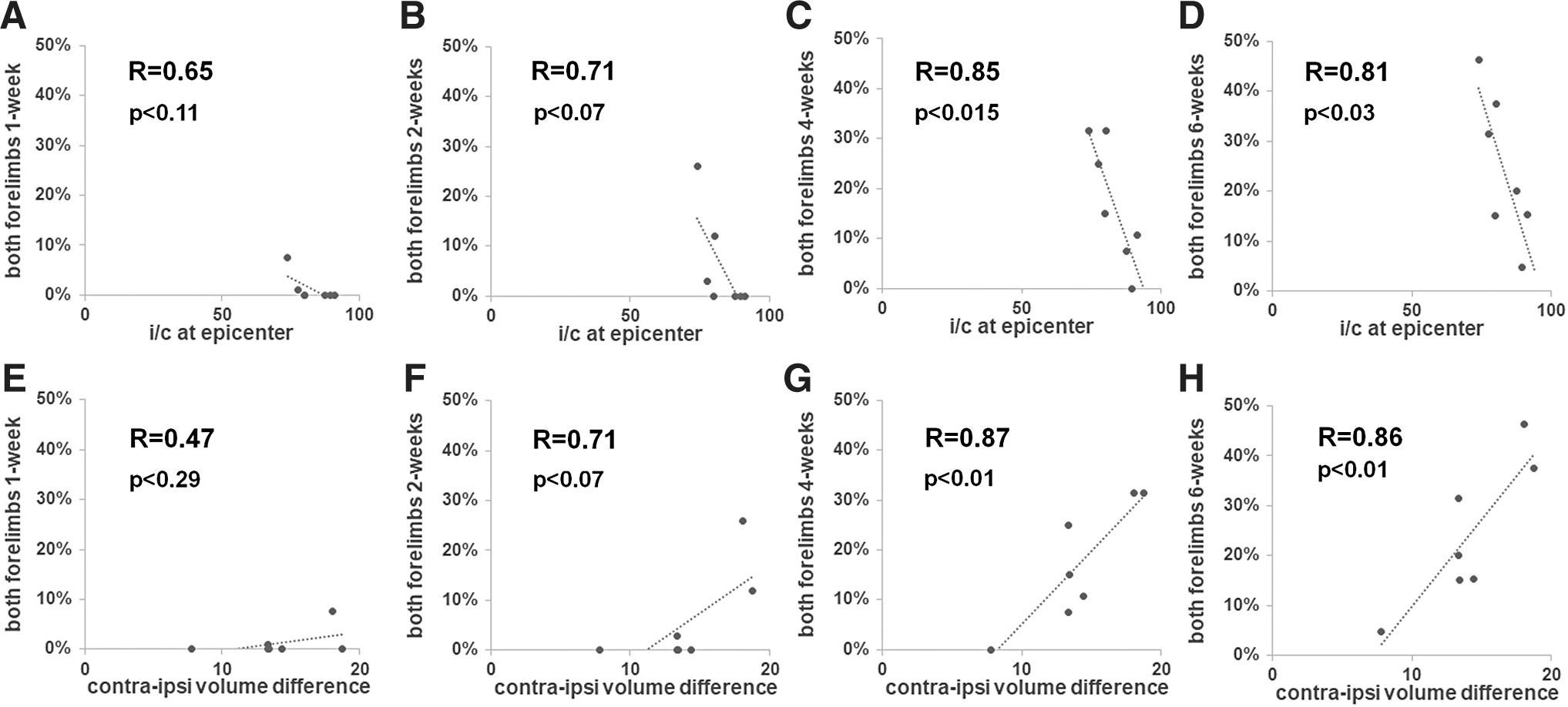
Relationship between the i/c ratio, contralateral–ipsilateral volume difference, and behavioral outcomes across seven SCI animal subjects.
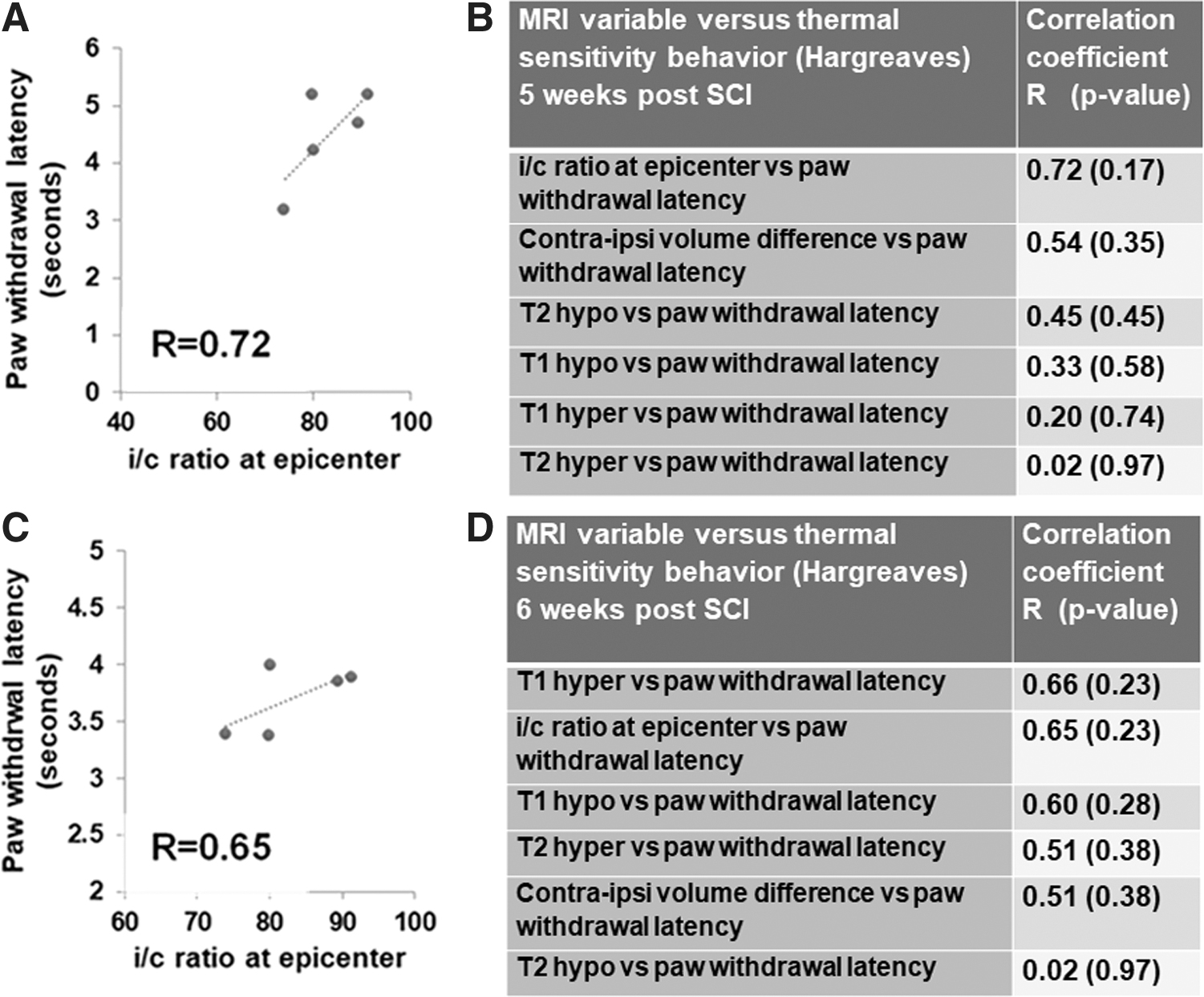
Relationship between MRI parameters and heat sensitivity behavior (Hargreaves) across five SCI animal subjects.
Discussion
Cervical spinal injuries are the most common in humans leading to sensorimotor deficits in the upper extremity, hence pre-clinical rodent models of incomplete cervical SCI at the middle of the cervical spine, producing forelimb sensorimotor deficits, have the most clinical relevance (Detloff et al., 2013; Putatunda et al., 2014). Animal model studies are required to evaluate therapies that target upper extremity functions to improve the quality of life for individuals with cervical SCI (Hilton et al., 2013; Onifer et al., 2005). Producing unilateral lesional damage at the dorsolateral funiculus and rubrospinal areas as determined by MRI, our injury model at the C4–C5 level demonstrated sustained forelimb motor dysfunction along with spontaneous neuropathic pain. Large-scale MRI-based spinal parenchymal changes occurred, revealing lesions and nonlesional spinal parenchymal volume loss, which related differently to the sensorimotor neurological outcomes.
Spinal pathophysiology underlying lesional and nonlesional morphometric parameters is coupled
Lesional volume had a CV of 30% and ipsilateral volume loss (nonlesional) had a CV of 35%, indicating a comparable subject-wise MRI outcome variability between lesional and nonlesional parameters. During the chronic phase, atrophic tissue is detected as T1 or T2 hypointense lesions and neuropil (i.e., neuronal, vascular, and axonal) dysfunctions are detected as T1 or T2 hyperintense lesions (Sundberg et al., 2010). T2 hypointensity correlated strongly with the i/c ratio at the epicenter (Fig. 4B; R = 0.65; p < 0.03) but moderately correlated with the contralateral–ipsilateral volume differences (R = 0.42; p < 0.20). These findings indicated that spinal cord atrophy was a major contributor to the spinal cord cross-sectional area changes at the injury epicenter more so than the overall ipsilateral volume loss across the entire cervical spinal cord. Additionally, the total lesional volume (T1 hypo+T1 hyper+T2 hypo+T2 hyper) correlated moderately with the total spinal cord volume loss across the SCI group (R = 0.50; p < 0.11), indicating that the underlying pathophysiological processes leading to spinal cord lesions or nonlesional volume changes are coupled to a certain extent.
Nonlesional spinal cord changes after SCI associate better with motor behavioral outcomes
Parenchymal volume change and the i/c ratio at the epicenter after SCI correlated strongly with individual motor behavioral outcomes both during the subacute and chronic phases (Fig. 5), indicating that spinal cord volume change could accurately predict the intensity of the trauma-induced motor debilitation in each subject. Although the different T1- and T2-based lesions showed low or moderate correlation between themselves (Fig. 4C), suggesting distinct underlying pathological processes represented by them, most of these lesional volumes correlated low with the forelimb motor behavioral outcomes (Supplementary Table S1). Correlation strengths did not differ between the nonlesional parenchymal volumetric variables and forelimb motor behavior when lesional volumes were either included (Fig. 5) or excluded from the estimated parenchymal volumes (data not shown). Hence, lesional volumes, by themselves, did not seem to have a reliable predictive value on motor outcomes when compared with spinal volumetric changes after SCI. Our MRI results on lesional volumes were similar to a recent diffusion MRI study of contusion SCI at the lumbar level, where pool size ratio measurements derived from quantitative magnetization transfer, indicative of myelination, progressively decreased at the injury epicenter and adjoining areas within 2 weeks after SCI (Wu et al. 2020). However, the pool size ratio changes had no association with sensory and motor behavioral outcomes (Wu et al., 2020). These results indicate that the nonlesional spinal cord may harbor the bulk of the reorganization related to motor circuits during the subacute and chronic phases after SCI.
Both lesional and nonlesional spinal cord changes after SCI associate with sensory behavioral outcomes
Ipsilesional forelimb sensory behavioral outcomes showed heat hypersensitivity at 5 weeks after SCI with low association to lesional volumes and moderate association to nonlesional spinal volumetric parameters (Fig. 6). However, ipsilesional forelimb sensory behavioral outcomes, which showed heat hypersensitivity at 6 weeks after SCI, associated moderately with T1 hyperintensity and the i/c ratio at the epicenter, where only the i/c ratio at the epicenter sustained its moderate association with the sensory behavioral outcomes at both 5 and 6 weeks after SCI (Fig. 6). The results indicate that both lesional and nonlesional areas of the spinal cord maybe dynamically involved in the reorganization of sensory circuits during the subacute and chronic phases after SCI. Maladaptive reorganization of sensory circuits after cervical SCI has been considered as a cause of neuropathic pain, which manifests at different injury stages in most cervical SCI patients. Neuropathic pain is notoriously difficult to treat as large disparities between pre-clinical and clinical studies have been reported (Kramer et al., 2017). From the current results, it can be hypothesized that sensory reorganization associates stronger with spinal lesional burden when compared with motor reorganization in SCI. While further studies are required to systematically test this hypothesis, current clinical evidence relates spinal lesions to chronic pain in specific spinal pathologies (Tackley et al., 2017). Glial activation, blood–brain barrier breakdown, and local inflammation have been shown to associate with neuropathic pain in the pre-clinical rat SCI model (Yamazaki et al., 2020). These neuropathological processes generally show up as central nervous system lesions in conventional T1 and T2 MRI sequences (Maranzano et al., 2017).
Nonlesional spinal volume changes and clinical similarities with multiple sclerosis
There have been some similarities between SCI and multiple sclerosis (MS) lesions in human studies (Pravata et al., 2020; Rocca et al., 2020). In a recent study, across 1249 individuals with MS, spinal cord volume, including both focal and diffuse pathological lesions, and their association with MS-related behavioral disabilities were analyzed (Andelova et al., 2019). Spinal cord volume had a stronger association with disability when compared with focal and diffuse pathological lesions (Andelova et al., 2019). Although focal lesions by themselves only moderately associate with neurological outcomes, they could be additive with diffuse microstructural changes in the spinal cord and contribute to MS-induced disability. Future simultaneous conventional MRI and DTI assessments may help determine whether the lesional and nonlesional spinal cord changes have any additive effects on neurological disability.
Translation to humans
The most important information from the current MRI study, not possible from a histological approach, is the accurate estimation of in situ volumetric changes in the entire cervical spinal cord after injury and its important relationship to the neurological outcome. Although cystic cavitation in the inured spinal cord is well known from previous histological investigations (Putatunda et al., 2014; Zareen et al., 2017), its exact association with behavioral outcomes is not known. The current study is the first to use MRI volumetry as used in humans to address structural changes in the pre-clinical rat model of cervical SCI. Volumetric changes from histology do not reflect the in situ values due to tissue fixation and further processing such as cryosection and mounting that are required. Additionally, information obtained from T2-MRI-based hyperintensity lesions specifically denotes functional deficits in surviving neural components, which again cannot be determined by conventional histological studies. Finally, using clinically relevant MRI biomarkers, the lesional and nonlesional associations with neurological behavioral outcomes obtained from this study can be seamlessly translated to humans using MRI, not possible with histological approaches due to their invasive nature.
One of the limitations of the study was the lower study sample numbers in the Hargreaves behavioral tests, which was performed on five SCI animals due to ipsilateral paw clench deficit not abating even after 5 weeks after SCI in two animals. Additionally, the MRI measurements lacked a subacute time point at 1 week after injury due to the terminal design of the MRI experiment, which may have provided information on the progression of the lesions with time, if performed longitudinally. Future investigations with larger samples along with longitudinal MRI studies with respiration-induced motion artifact mitigation can overcome these limitations.
Conclusions
The present study demonstrates that lesional and nonlesional spinal cord changes in T1- or T2-based conventional MRI have different associations with sensory and motor neurological outcomes after incomplete cervical SCI. The results have important ramifications for the utilization of conventional MRI-derived lesional and nonlesional parameters in recovering SCI patients, testing for specific spinal morphological associations with functional connectivity, sensory and motor neurological recovery with and without specific treatment interventions.
Footnotes
Authors' Contributions
S.S.K. conceived and designed the study. J.C., B.G.S., P.H., and S.S.K. performed the experiments. J.C. performed formal analysis of the behavioral data. S.S.K. performed the formal analysis of the MRI data. R.H., S.E., L.N., and F.H. contributed to significant laboratory resources and expertise for the generation of the rat model of cervical SCI and MRI experiments. S.S.K. wrote the first draft of the article, and all the authors edited and approved the submitted version of the article.
Author Disclosure Statement
All authors disclose that no competing financial interests exist.
Funding Information
This study was funded by the New Jersey Commission for Spinal Cord Research CSCR18ERG018 (S.S.K.) and R01 MH067528 (F.H.).
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.