
Abstract
Background:
The thalamus, as a key relay of neuronal information flow between subcortical structures and cortical networks, has been implicated in focal limbic seizures propagation, awareness maintenance, and seizure-related cognitive deficits. However, the specific functional alterations between different thalamic nuclei and subcortical-cortical systems in temporal lobe epilepsy (TLE) remain largely unknown.
Methods:
We examined thalamic functional connectivity (FC) in 26 TLE patients and 30 healthy controls matched for sex, age, and education. The anterior (ANT), ventral posterior medial, and central lateral nuclei of thalamus were employed to establish whole-brain seed-to-voxel thalamic FC maps. Secondary Pearson's correlation analysis was conducted to assess associations between the abnormal thalamic FC and the memory performance in TLE.
Results:
Seed-based FC analyses revealed typical distinct FC patterns within each thalamic nuclei in both controls and TLE patients. The TLE showed significantly decreased FC between different thalamic nuclei and subcortical-cortical networks, including the limbic structures, midbrain, sensorimotor network, medial prefrontal cortex, temporal-occipital fusiform gyrus, and cerebellum. Verification analyses yielded similar patterns of thalamic FC changes in TLE. Importantly, the decreased FC between the ANT and hippocampal pathway was correlated with the poorer memory performance of TLE.
Conclusion:
These findings suggest that the distinct thalamocortical FC patterns are damaged to some extent in TLE patients. Importantly, the specific pathology of the ANT–hippocampal pathway in TLE may be a potential factor that contributes to memory deficits. Our study may pave the way for improved treatments and cognitive function by directly targeting different thalamocortical circuits for TLE.
Impact statement
This study demonstrates prominent decreased functional connectivity (FC) between different thalamic nuclei and subcortical–cortical networks in patients with temporal lobe epilepsy (TLE). Moreover, the specific pathology of the anterior nuclei of the thalamus and hippocampal pathway was found to be related to the memory deficits in TLE. These findings suggest the divergent thalamocortical FC is damaged to some extent in TLE patients, which may have the potential to greatly guide future clinical treatments by targeting different thalamocortical networks.
Introduction
Temporal lobe epilepsy (TLE) is the most common adulthood form of focal epilepsy, and is characterized by seizures that often originate from limbic structures including the hippocampus (Lanerolle et al., 1989; Williamson et al., 1993). However, this type of focal seizures usually demonstrates remote changes, including those in the cortical and subcortical regions, as evidenced by electrophysiological, functional magnetic resonance imaging (fMRI), and metabolic data (Benedek et al., 2004; Blumenfeld, 2014; Englot et al., 2017). Previous studies suggest that the thalamus, as a key relay or processing hub of neuronal information flow between subcortical–cortical structures, is implicated in focal limbic seizures with impaired awareness, cognitive deficits, and seizure propagation (Feng et al., 2017; Motelow et al., 2015; Vertes et al., 2001).
The thalamus consists of a group of nuclei with reciprocal and topographically organized connectivity with the cortex, limbic system, and subcortical regions (Jones, 2012; Morel et al., 1997; Poulet et al., 2012). In particular, the anterior nuclei of the thalamus (ANT) serves as a crucial bridge between the prefrontal and limbic systems and has become an attractive target for deep brain stimulation (DBS) to treat medically refractory epilepsy because of its possible role in seizure propagation (Herrman et al., 2019; Li and Cook, 2018). Cases with anterior lesions of the thalamus show a pure deficit of memory, thus illustrating the specificity of the anterior nuclei for declarative memory processes (Aggleton et al., 2010; Jankowski et al., 2013; Van der Werf et al., 2000). The central lateral nuclei (CL) of the thalamus, which forms part of the intralaminar thalamic group, demonstrates reduced overall neuronal activity during seizures, which may contribute to depressed cortical function and loss of consciousness during TLE (Feng et al., 2017; Van der Werf et al., 2002). The ventral posteromedial nucleus (VPM) of the thalamus mediates somatosensory information from the face (trigeminal) to the primary somatosensory cortex (Oda et al., 2004), which has been found to show sleep spindles during focal seizures (Feng et al., 2017). Researchers consistently agree that heterogeneous thalamic nuclei may be involved in different arousal, motor, and memory circuits during focal seizures in animal models (Englot et al., 2009). However, the specific functional projections of these different thalamic nuclei have not been fully investigated in patients with TLE. Further understanding of distinct thalamocortical circuits in TLE may lead to improved treatments directly targeting different modes of impaired brain function.
Resting-state fMRI provides a powerful tool to investigate intrinsic functional connectivity (FC) and seizure-related functional circuit changes in epilepsy (Englot et al., 2015; Li et al., 2016, 2018). Recent work has highlighted distinct connectivity patterns within thalamic structures, showing the intrinsic connectivity based on mapping of the thalamocortical relationships (Kim et al., 2013; Yuan et al., 2016). Parcellation of the thalamus based on different thalamocortical functional connections has been efficiently investigated in healthy subjects (Dongyang et al., 2008). In addition, Xiaosong et al. (2015) has performed thalamic parcellation based on FC between five nonoverlapping cortical seeds and thalamus and reported reduced thalamocortical connectivity in patients with TLE. However, these and previous studies have only either generally focused on overall thalamocortical connectivity changes or investigated relations to individual cortical lobes and regions. The overall FC patterns of different thalamic nuclei and its alterations in TLE remain largely unknown. Our recent work from the rodent model of focal limbic seizures showed divergent neuronal activity patterns in three different thalamic regions, respectively contributing to seizure propagation, cortical suppression, and information transportation (Feng et al., 2017). We found that population firing of neurons in CL decreased whereas ANT showed a trend toward increased neuronal firing. Meanwhile, VPM exhibited a remarkable increase in sleep spindles during focal seizures. These very different activity patterns within the thalamus suggested that the thalamus might play diverse roles in cortical–subcortical brain activity patterns, which call for further confirmation in human studies. However, the specific functional projections of these different thalamic nuclei have not been fully investigated in patients with TLE.
Here, we performed resting-state FC analysis of multiple thalamus nuclei based on the anatomical thalamic segmentation templates to investigate distinct thalamic FC patterns in TLE. Three thalamus seeds including ANT, VPM, and CL were specifically selected based on their important roles in seizure propagation and behavioral manifestations. We hypothesized that spatially distinguishable FC patterns would be identified across these three typical thalamic nuclei. We further explored the relationship between these abnormal thalamic FCs and the neuropsychological function of TLE. Given that the ANT is generally assumed to be a key constituent of the neural circuits responsible for memory processes, we predicted that abnormal ANT connectivity would correlate with memory performance in TLE.
Materials and Methods
Subjects
The subjects in the present work came from our previous study (Li et al., 2019), and all of the data were collected between January 2012 and October 2014. Twenty-six TLE patients diagnosed with complex-partial seizures were consecutively and prospectively recruited from the Department of Neurology, Xiangya Hospital of Central South University. TLE was diagnosed through comprehensive evaluation, including seizure history and semiology, neurological examination, diagnostic MRI, and EEG records. The inclusion criteria for patients were: (1) a TLE diagnosis in accordance with the International League Against Epilepsy classification (Berg et al., 2010); (2) complete clinical data and more than two epileptic episodes a year; (3) complex-partial seizures in which the patient has impaired responsiveness to verbal questions or commands or amnesia; (4) aged 14–50 years; and (5) with an education level of primary school and above and right-handedness. The exclusion criteria were: (1) a diagnosis of serious medical or mental disorders based on health screening and psychiatric measures; (2) identifiable brain abnormality on structural MRI other than hippocampal sclerosis identified by conventional diagnostic MRI; and (3) history of long-term use of drugs other than antiepileptic drugs (AEDs). Among the 16 patients treated with medication, 12 received monotherapy, 4 received polytherapy, 9 were treated with valproic acid, and 3 were treated with carbamazepine. All the TLE patients avoided other psychotropic medications except for usual treatment (AEDs) on the day of fMRI scanning. The detailed clinical information for the patients is provided in Supplementary Data.
In addition, a total of 30 healthy controls matched for sex, age, and education were randomly recruited in the current study. Healthy subjects were free of neurological or systemic disorders and psychiatric illnesses and had no gross abnormalities on brain magnetic resonance images. This study received ethical approval by the Ethics Committee of Xiangya Hospital of Central South University, and all subjects provided written informed consent before fMRI scanning (see Supplementary Fig. S1 for procedure of inclusion/exclusion of the participants).
Neuropsychological evaluation
The TLE patients and healthy controls underwent neuropsychological testing of logical memory, which was conducted by two independent neurologists. The logical memory test, a subtest of the Wechsler Memory Scale-Revised (WMS-R) (Wechsler, 1987), assesses the immediate and delayed recall performance in verbal memory by asking the subject to recall two stories immediately after an oral presentation (Part I) and again after a 30 min delay (Part II). For each subject, all the neuropsychological evaluation and fMRI scanning was performed on the same day.
Imaging acquisition
All participants were scanned on a 3.0 Tesla MRI scanner (Siemens Magnetom Trio, A Tim System, German 3.0T). All fMRI images were collected by using a gradient-echo echo-planar imaging sequence. A 12-channel prototype quadrature birdcage head coil fitted with foam padding was used to minimize head movement. The acquisition parameters for functional imaging were as follows: repetition time/echo time = 3000 msec/30 msec, flip angle = 90°, field of view = 240 × 240 mm2, matrix = 64 × 64, 36 transverse slices with slice thickness = 3 mm, and interslice gap = 1 mm. Participants lay on a foam pad to comfortably stabilize the head and were instructed to simply rest with their eyes closed, keep their body still, and try not to think or fall asleep. A total of 140 frames were acquired for each participant, lasting 266 sec. High-resolution 3D T1-weighted anatomical images were also acquired in the sagittal orientation by using a magnetization-prepared rapid acquisition gradient-echo sequence (repetition time/echo time = 1900 msec/3.41 msec, field of view = 100 × 100 mm2, matrix = 256 × 256, slice thickness = 1 mm, and interslice gap = 1 mm).
Data preprocessing
Functional data preprocessing was conducted by using the DPARSF-A-toolbox. The initial five functional images of each subject were discarded for magnetic saturation, resulting in 135 frames. Subsequently, we performed slice timing correction to adjust for variable acquisition times over slices in a volume, with the middle slice in every volume used as a reference. Next, six-parameter variance cost function rigid-body affine registration was used to realign all images within a session to the first volume. We required that transient movement during scanning be no more than 2 mm of translation and 2 degrees of rotation. Each 3D T1-weighted structural image was then coregistered with its functional image and segmented into gray matter, white matter (WM), and cerebrospinal fluid (CSF). The functional images were spatially normalized to the Montreal Neurological Institute (MNI) template at a resolution of 2 × 2 × 2 mm3. Then, the images were spatially smoothed with an full width half maximum of 6 mm. The resulting data were further linear detrended and band-pass filtered (0.01–0.08 Hz) to reduce the effects of low-frequency drifts and high-frequency physiological noise. Finally, several sources of nuisance signals (i.e., 24 head motion parameters, averaged signals from the CSF, WM, and global signals) were regressed out by using multiple linear regression analysis.
Additional artifact removal was performed before further data analysis. Signal-to-noise ratio was calculated over time for each voxel [20 × log (mean blood oxygen level dependent [BOLD] signal/standard deviation of the BOLD signal)] and then averaged over the whole brain. An arbitrary cut-off of 30 dB was set for exclusion; however, because all of the fMRI runs of the TLE patients and healthy controls exceeded the threshold, no run was deleted in this step. Previous studies have identified the possible influence of motion on FC results in resting-state fMRI investigations (Power et al., 2012). Thus, the mean framewise displacement (FD) was calculated as the sum of the absolute values of changes in the six parameters for transitional and rotational displacements between volumes.
Definition of regions of interest
The goal of the current study is to demonstrate distinct FC patterns in different thalamic nuclei in TLE. The Morel stereotactic atlas was used to identify thalamic nuclei (Morel et al., 1997). This atlas was constructed based on human postmortem histology and built in the MNI space, including 40 small thalamic nuclei. We selectively employed three thalamic regions of interest (ROIs) per hemisphere based on their important roles in limbic seizure networks: ANT (which consists of three subnuclei, including anteroventral, anterodorsal, and anteromedial nuclei), VPM, and CL. The seeds volume of the left ANT was 45 voxels; the right ANT was 39 voxels; the left CL was 118 voxels, and the right CL was 108 voxels; for the left and right VPM, it was 23 and 26 voxels, respectively. We resampled the atlas from native space to the 2 mm isotropic space and employed the MRIcroN software for visual inspection of these three thalamic nuclei locations (Supplementary Fig. S2).
FC of thalamic nuclei
We analyzed FC by using resting-state fMRI data after preprocessing to explore the role of different thalamic nuclei in TLE and verify our hypothesis. For each subject, the mean BOLD signal time course was extracted from each of the six thalamic ROIs. Pearson's correlation coefficients were computed, respectively, between the average time course of all voxels in the six thalamic ROIs and for all other voxels within the whole brain. The resulted r values were then converted into z scores by using Fisher's r-to-z transformation to improve the normality of their distribution. Individual FC maps were entered into group-level analyses. First, one-sample t-tests were performed on within-group thalamic nuclei FC maps to identify brain regions showing significant connectivity with seed regions. Here, the significance level was set to p < 0.05 with an extent threshold of 10 contiguous voxels by using the false discovery rate (FDR) correction. For the reason that negative connectivity mechanisms in the context of network physiology are less understood and have been a subject of debate in terms of its origin and interpretation (Murphy et al., 2009), we set negative connections to zero to avoid uncertainty and merely extracted the positive FC connections from the one-sample t-tests of TLE patients and healthy controls, respectively. The resulted positive FC maps of each seed in two groups were combined as the templates for the second-level group comparisons. First, to examine the overall thalamic connectivity changes in TLE, we performed two-sample t-tests comparing the mean strength of positive connections between TLE and controls under these templates. Then, seed-to-voxel two-sample t-tests were performed to further examine the significant brain regions showing thalamic FC changes in TLE. The FC analysis was performed by using the REST V1.8 package, and statistical analysis was conducted by using the Statistical Parametric Mapping software. Gaussian random field (GRF) theory was employed to carry out cluster-level correction for multiple comparisons (minimum z > 2.3 and the cluster significance was set to p < 0.05).
Neuropsychological correlation analysis
We hypothesized that abnormal ANT connectivity would contribute to neuropsychological dysfunction, such as logical memory deficits. Previous investigations have emphasized the multiple hippocampal–anterior thalamic interconnections in memory processes (Aggleton et al., 2010; Jankowski et al., 2013; Mair et al., 2003; Van der Werf et al., 2000). Here, we explored the potential relationship between ANT and hippocampal and parahippocampal connectivity and logical memory scores to investigate the neuropsychological relevance of altered FC in TLE. Four patients without available neuropsychological data were excluded in this analysis. Pearson's correlation analysis was utilized to assess associations between the FC and the immediate and delayed recall memory scores, controlling for age, gender, and mean FD as confounding variables. First, we created ROIs by intersecting corrected brain clusters showing significantly FC differences in ANT-based FC group comparisons with the corresponding hippocampal and parahippocampal masks from Automated Anatomical Labeling templates. Then, we extracted the mean z values of each individual ROI and calculated the Pearson's correlations between the z values and memory scores of TLE patients. Bonferroni significance level of p < 0.05 dividing by the number of comparisons was used for correction. To test whether relationships between ANT–hippocampal FC and memory scores were specific or correlations would be seen in other nuclei, we also investigated the potential correlations between memory performance and FC in other thalamocortical circuits.
Verification analyses
We performed additional analyses to verify the stability of our main findings. First, given that the functional changes in new-onset patients and long-term patients may be different, we assessed the effects of disease on the thalamic FC by, respectively, comparing new-onset (n = 7) and long-term (n = 19) patients with controls. Second, several studies have shown a significant influence of AEDs treatment on functional properties of brain (Hamed, 2009); we provided additional analysis to assess the potential treatment effects on the thalamic FC. The analysis was repeated by separately comparing group differences of thalamic FC between healthy controls and drug-naive (n = 10) and medicated (n = 16) patients. Finally, we investigated whether the seizure lateralization of TLE would change the thalamic FC patterns. The FC maps of 16 left TLEs and 10 right TLEs were compared with controls, respectively. The detailed information for the verification analyses is provided in the Supplementary Data.
Results
Demographic and clinical data
None of the participants exceeded the maximal threshold of translation and maximal rotation, and the mean FD of all the subjects was less than 0.3 mm. Patients and controls did not differ significantly in terms of sex ratio, age, education level, or mean FD. The TLE patients had slightly lower WMS-R immediate and delayed recall memory performance scores compared with healthy controls, but no statistical difference was found between groups (p = 0.19 and p = 0.17, respectively). Details regarding patients' epilepsy history and seizure semiology, including epilepsy duration, frequency, seizure type, onset age, and side of seizure, were obtained from comprehensive clinical assessments by epileptologists. The clinical and demographic information of all participants are summarized in Table 1.
Demographic and Clinical Characteristics
Data are presented as mean ± standard deviation.
Two-sample t-test.
Chi-square test.
FD, framewise displacement; HC, healthy controls; L, left; R, right; TLE, temporal lobe epilepsy.
Distinct spatial patterns of thalamic FC
We first detected the FC patterns of different thalamic nuclei within healthy control and patient groups. The FC spatial distributions for each of the three thalamic seeds per hemisphere in the control group and TLE patients are shown in Figures 1 and 2, respectively (p < 0.05, FDR corrected). One-sample t-tests revealed the typical distinct spatial FC patterns of different thalamic seeds for both controls and TLE groups. Consistent with prior studies emphasizing the involvement of ANT in limbic associations (Dalrymple-Alford et al., 2015; Jankowski et al., 2013; Van der Werf et al., 2003), ANT was mainly projected toward the limbic structures, including the hippocampal formation and a part of parahippocampus, temporal areas, much of the anterior cingulate gyrus, insula, basal ganglia, and the remaining thalamic nuclei; it also displayed connectivity with cortical areas, including the medial and lateral prefrontal cortex, inferior parietal lobule, and angular gyrus. Unlike the anterior seeds, VPM was more highly correlated with central regions that were implicated in somatosensory processes, such as the precentral gyrus, postcentral gyrus, supplementary motor area, rolandic cortex, and cerebellum, as well as insula, basal ganglia areas, and other thalamic nuclei. The CL strongly correlated with the brainstem in the midbrain and pons and extended to the limbic parahippocampus and hippocampus, basal ganglia, basal forebrain and other thalamic nuclei, as well as the middle frontal gyrus and visual cortex. More region details are shown in Supplementary Table S1 and S2.
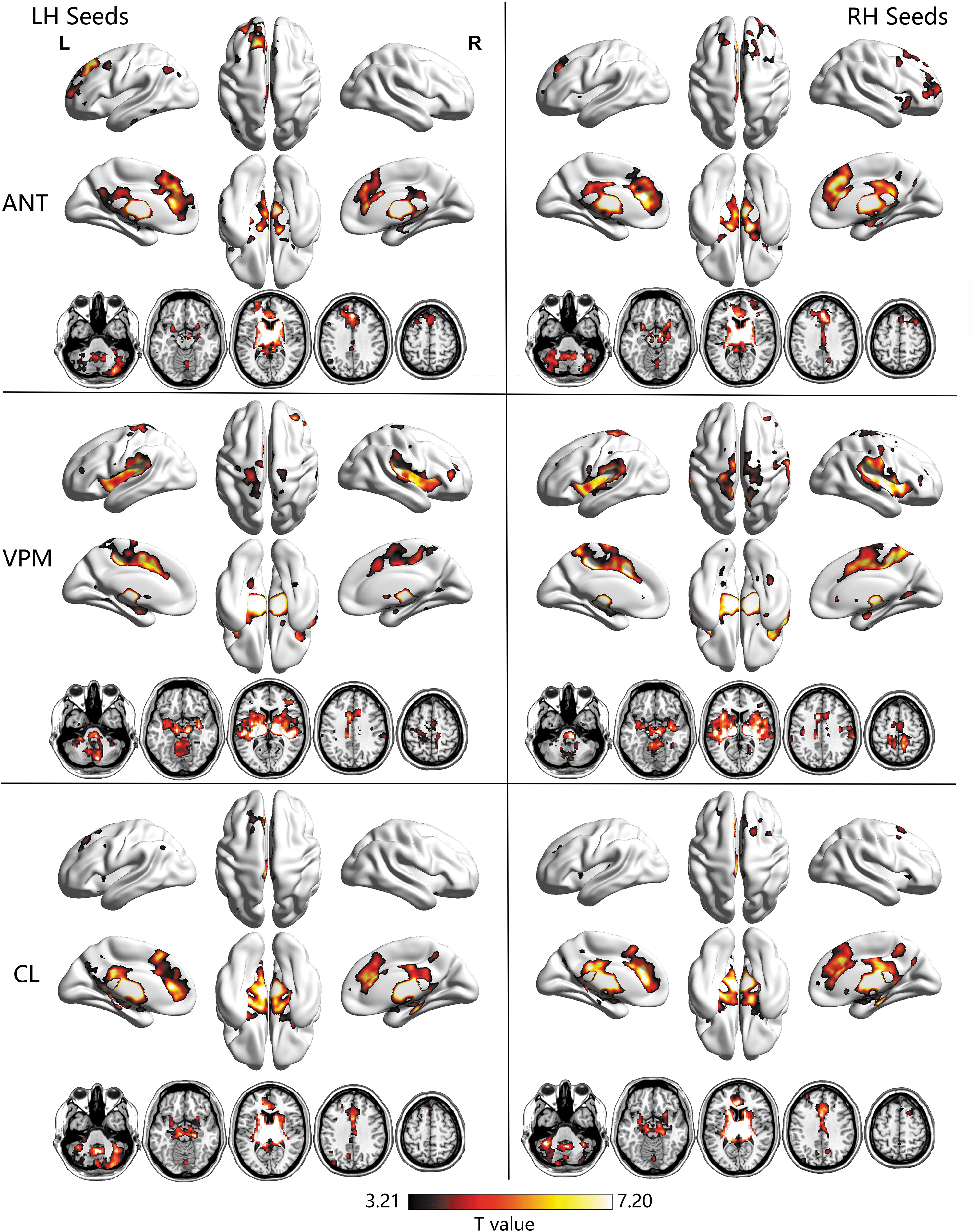
Brain regions showing statistically significant functional connectivity, with each thalamic seed in healthy controls. Within-group results were obtained by one-sample t-test using FDR correction with p < 0.05. ANT, anterior nuclei of thalamus; CL, central lateral nuclei; FDR, false discovery rate; LH/RH, left/right hemisphere; VPM, ventral posterior medial nuclei. Color images are available online.
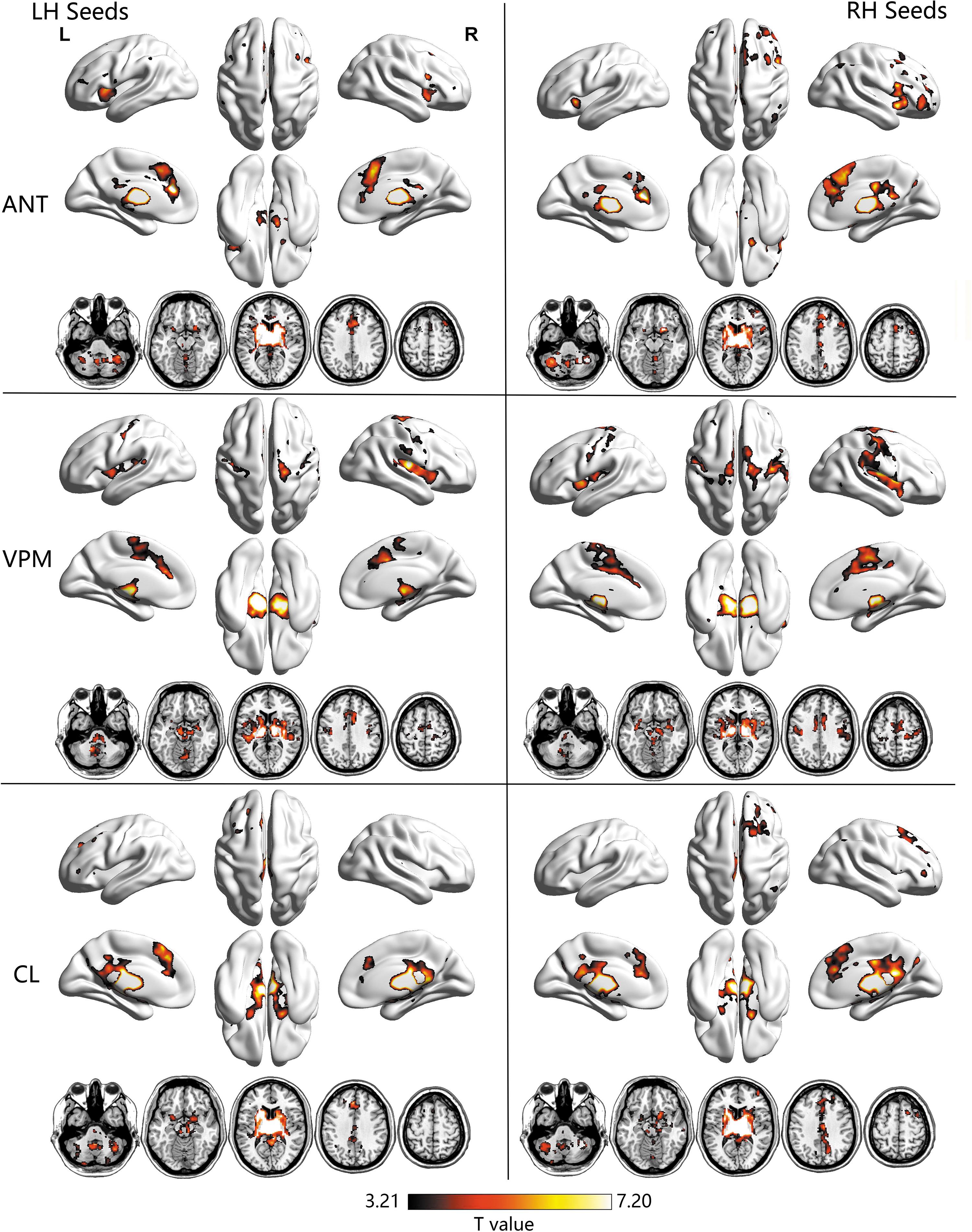
Brain regions showing statistically significant functional connectivity with each thalamic seed in TLE patients. Within-group results were obtained by one-sample t-test using FDR correction with p < 0.05. TLE, temporal lobe epilepsy. Color images are available online.
Abnormal thalamic connectivity in TLE
The comparisons of overall mean strength of connections between TLE and patients did not reveal any significant group difference. Seed-to-voxel two-sample t-test showed significantly abnormal FC between the six ROIs and a number of subcortical and cortical regions in patients with TLE relative to healthy controls. Table 2 and Figure 3 show the results of between-group FC comparisons with the thalamic seeds (GRF corrected, minimum z > 2.3, and cluster significance p < 0.05). Specifically, TLE patients showed significantly decreased FC between bilateral ANT and brain clusters of the hippocampus, parahippocampus, and anterior cingulate cortex extending to the medial prefrontal cortex but increased FC in the superior parietal gyrus and dorsal lateral prefrontal cortex. The left VPM revealed significantly decreased FC in the postcentral gyrus and cerebellum posterior lobe, whereas the right VPM indicated decreased FC in the rolandic operculum, insula cortex, postcentral gyrus, and cerebellum. Bilateral CL seeds indicated decreased FC in the hippocampus and parahippocampus clusters, including those in the midbrain, temporal-occipital fusiform gyrus, and cerebellum anterior lobe.
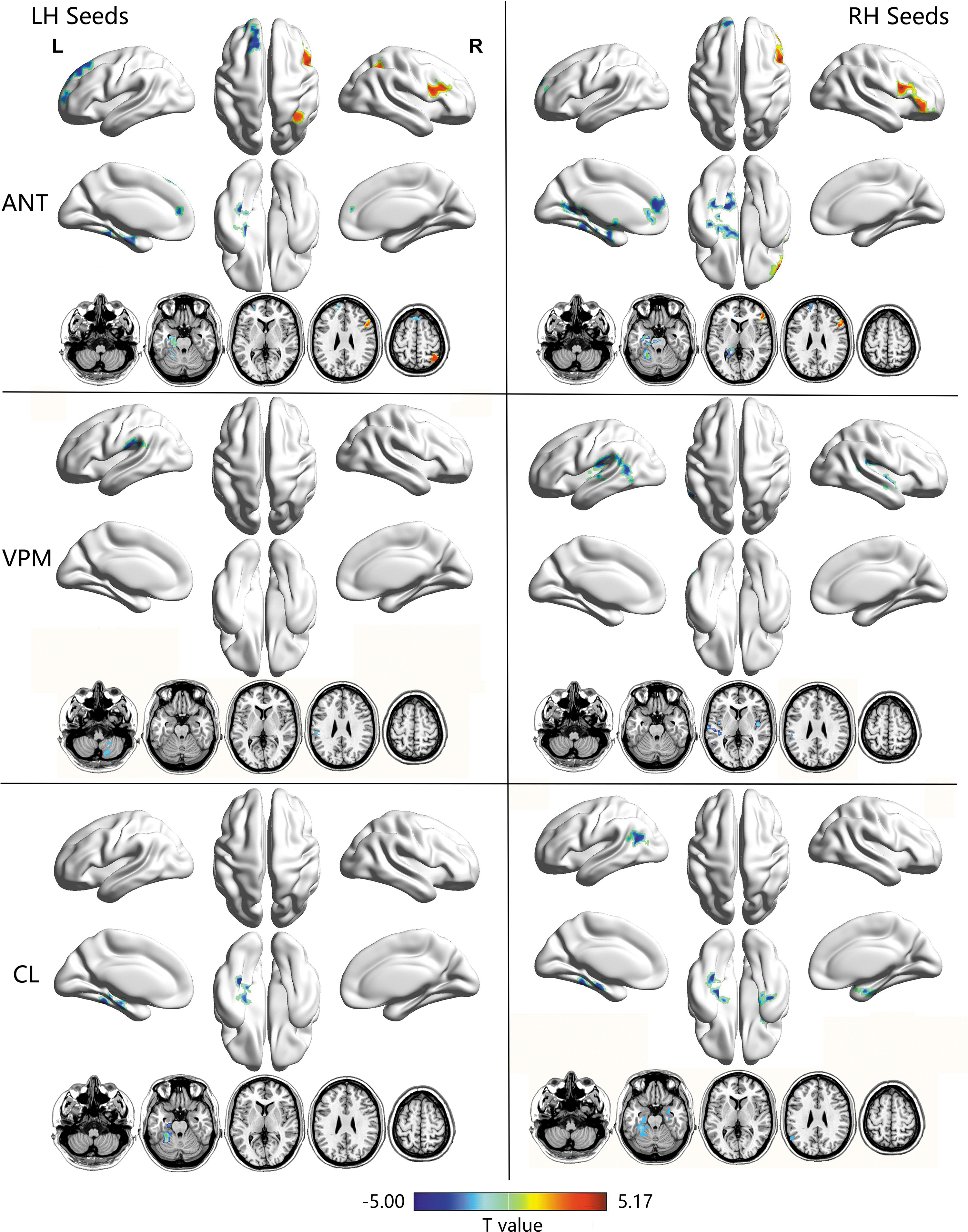
Functional connectivity maps of thalamic seeds showing significant group differences between TLE patients and healthy controls. Between-group results were obtained by two-sample t-test. Gaussian random field theory was used for cluster-level multiple comparisons correction (minimum z > 2.3 and cluster significance was set to p < 0.05). Cool colors display regions where functional connectivity was decreased in TLE; warm colors display regions where functional connectivity was increased in TLE. Color images are available online.
Regions Showing Group Differences Between Temporal Lobe Epilepsy and Healthy Controls in Thalamic Functional Connectivity
All clusters survived the minimum z > 2.3 and cluster significance p < 0.05, GRF theory corrected. Cluster size is reported in number of voxels, and stereotaxic coordinates are reported in the MNI standard space.
ANT, anterior nuclei of thalamus; BA, Brodmann area; CL, central lateral nuclei; GRF, Gaussian random field; L/R, left/right hemisphere; MNI, Montreal Neurological Institute; TLE, temporal lobe epilepsy; VPM, ventral posterior medial nuclei.
Correlations between the anterior thalamic–hippocampal/parahippocampal FC and the memory performance in TLE patients
Given the finding of decreased anterior thalamic–hippocampal/parahippocampal connectivity in patients and the important role of the ANT network in memory processes, we examined the relationship between the observed abnormal connectivity and memory scores in patients. Except for the correlation between the right ANT-hippocampal FC and immediate memory scores (r = 0.36, p = 0.0999), significant positive correlations between the immediate and delayed recall memory scales, and the mean z values of each individual hippocampal and parahippocampal ROI were identified, except for all p < 0.05 (shown in Fig. 4). In particular, the correlations between the left ANT–parahippocampal FC and immediate (r = 0.66, p = 0.0007) and delayed (r = 0.60, p = 0.00330) memory scores could survive multiple-comparison Bonferroni corrections. We found no significant correlation between the memory scores and other thalamocortical connectivity (Supplementary Fig. S9 and Supplementary Table S3), supporting the specific role of the ANT-hippocampal pathway in processing memory.
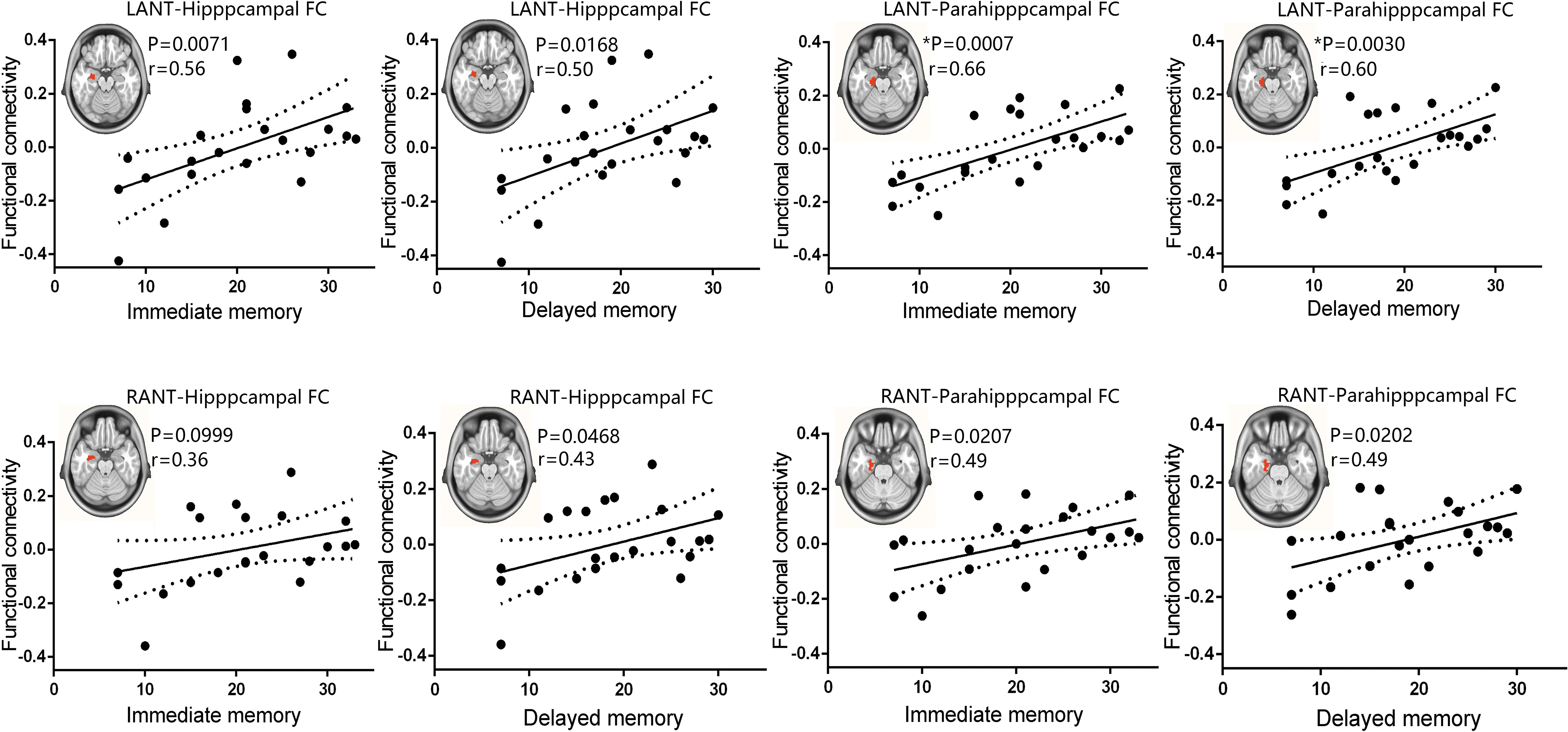
Correlations between ANT–hippocampal functional connectivity and memory performance of TLE patients. Functional connectivity between the left ANT and the left hippocampus, between the left ANT and parahippocampus, between the right ANT and the left hippocampus, and between the right ANT and the left parahippocampus was positively correlated with the immediate or delayed recall memory scores in TLE (Pearson correlation; p < 0.05, uncorrected; *p < 0.05, Bonferroni corrected). The FC between the right ANT and the left hippocampus was slightly correlated with the immediate memory scores (p = 0.0999). LANT, left anterior nuclei of thalamus; RANT, right anterior nuclei of thalamus. Color images are available online.
Verification analyses
The significant group differences of thalamic FC were reanalyzed with different variables of epilepsy duration and AEDs to confirm the reproducibility. In these verification analyses, the group differences of thalamic FC closely resembled the main findings in Figure 3. Compared with the controls, distinct thalamic FC changes were found in subcortical–cortical networks in TLE patients (Supplementary Figs. S3–S6). This indicates that the abnormal FC patterns of different thalamic nuclei are still present when controlling to seizure duration and AEDs. Finally, we examined the seizure lateralization effect on the main findings. In general, this analysis revealed very similar FC patterns to the main findings. In addition, we found that the left and right TLE patients exhibited decreased ANT–hippocampal connectivity in ipsilateral and contralateral hemispheres, whereas they exhibited only increased ANT-cortical connectivity in the right side of the brain. Meanwhile, the left TLE patients showed ANT-based decreased FC in the medial prefrontal cortex whereas the right TLE patients did not (Supplementary Figs. S7 and S8).
Discussion
In this study, we utilized seed-based resting-state fMRI to investigate the thalamic functional architecture in patients with TLE. Consistent with the functional heterogeneity of thalamus, we demonstrated that patients with TLE exhibited prominently abnormal thalamic FC with distinct subcortical and cortical networks. In particular, ANT demonstrated notably decreased FC with limbic structures, including the hippocampus, parahippocampus, anterior cingulate cortex, and the medial prefrontal cortex, whereas they exhibited increased FC with the dorsal lateral prefrontal cortex and parietal cortex. The VPM exhibited decreased FC mainly in the sensorimotor network and cerebellum. The CL showed significantly decreased FC with the temporal–occipital fusiform gyrus and multiple subcortical regions. These findings suggest that different thalamic nuclei own distinct connectivity patterns and may contribute to the understanding of network mechanism underlying focal temporal lobe seizures. More importantly, we found that FC between the ANT and hippocampus/parahippocampus was specifically correlated with the memory performance of TLE, indicating that the abnormality of the anterior thalamic–hippocampal pathway may be a potential factor that could contribute to memory deficits in TLE.
The typical distinct spatial FC patterns of different thalamic seeds we identified in healthy controls are in agreement with previous resting-state fMRI studies showing intrinsic connectivity between thalamus voxels and corresponding cortical ROIs (Zhang et al., 2008). For example, we found that the ANT showed prominent connectivity with prefrontal cortical areas whereas VPM corresponds approximately to the somatosensory cortex. Although abnormal thalamocortical FC in TLE has been previously reported (Barron et al., 2015; Xiaosong et al., 2015; Xue-Mei et al., 2015), the specific potential FC abnormalities involved in different thalamic nuclei have not been evaluated. Here, we investigated the FC patterns of the thalamocortical system by employing atlas-based anatomical thalamic subnuclei. The results revealed that different thalamic nuclei may be involved in distinct circuit systems, which could be disrupted by seizures, resulting in different clinical manifestations. An important finding of our study is that TLE patients showed significantly decreased FC between bilateral ANT and limbic structures, including the hippocampus, parahippocampus, and anterior cingulate cortex extending to the medial prefrontal cortex. Among the multiple functional pathways, ANT occupies the superior region of the thalamus with distinct connectivity with the hippocampal formation, mammillary bodies, anterior cingulate, and orbitomedial prefrontal cortex (Child and Benarroch, 2013). Early animal models have shown that the ANT circuit is one of the most important neuroanatomic circuits in focal seizure propagation (Takebayashi et al., 2007). In addition, ANT has been the target of DBS in several studies for seizure inhibition and treatment of intractable focal epilepsy (Lim et al., 2007). In a recent resting-state fMRI study, Morgan et al. (2015) showed that the anterior nucleus and pulvinar of the thalamus are highly susceptible to TLE seizures. It seems quite plausible that ANT stimulation or lesions may suppress the function of the ANT during the amplification, propagation, and synchronization of seizure activity. Consistent with the role of ANT in these processes, our work provides direct neuroimaging evidence, suggesting that the ANT–hippocampal–cortical circuit is disturbed by repetitive focal temporal lobe seizures.
It is interesting that increased right ANT-cortical connectivity was found in TLE patients. Increased activity or connectivity has been reported in areas outside the epileptogenic regions in previous epilepsy studies (Bai et al., 2011; Mankinen et al., 2011). Bettus et al. (2009) reported decreased FC within epileptogenic networks with concomitant contralateral increased connectivity in TLE. We speculate that the increased FC between ANT and higher association areas, including the dorsal lateral prefrontal cortex and parietal lobule in our study, may represent compensatory mechanisms aiming at functionally limiting the effects of brain injury inside the thalamolimbic circuits. Nevertheless, it remains unclear as to why the right TLE patients also showed ipsilateral increased FC with an uncorrected height threshold, which should be investigated further in future work. In addition, we observed no significant laterality of decreased ANT–hippocampal connectivity when separately comparing left and right TLE patients with controls, whereas there was a left laterality when patients merged together. This observation is likely related to a higher number of left TLE relative to the right TLE patients in our study. Another interesting observation is that the left TLE patients showed ANT-based decreased FC in the medial prefrontal cortex whereas the right TLE patients did not. This is in accordance with a previous study that demonstrated greater connectivity changes in left TLE compared with right TLE (Haneef et al., 2014). In investigations of memory function in clinical populations, TLE is of particular interest because of the frequent presence of hippocampal lesions. The ANT is assumed to be a key constituent of the neural circuits responsible for memory processing of the contents of stimuli for storage and recall (Aggleton et al., 2010; O'Mara, 2013). Previous studies have suggested that extensive direct and indirect anterior thalamic-hippocampal connections constitute a neuronal network that is crucial for memory processes (Bridge et al., 2017). Clinical and experimental evidence indicates that damage to the ANT or its inputs from mammillary bodies primarily contribute to episodic memory deficits (Law and Smith, 2012). In addition, as a structure lies adjacent to the hippocampus, the parahippocampus has extensive connections to hippocampus and also plays an important role in memory (Van Strien et al., 2009). The specific positive correlations between the anterior thalamic–hippocampal/parahippocampal FC and memory performance are particularly interesting in this context. This finding demonstrates that the multiple functional interactions between the anterior thalamus and the hippocampal and parahippocampal circuits have important implication in memory processes. The pathology of the anterior thalamic–hippocampal/parahippocampal pathway may be a potential factor contributing to memory impairments in TLE patients. Our findings point out the potential clinical application in future by monitoring anterior thalamic–hippocampal/parahippocampal FC in patients with TLE for long-term memory evaluation.
Evidence demonstrates that the VPM has direct connections to the somatosensory cortex and that electrical stimulation of the VPM may lead to postsynaptic activation in the epileptic focus (Caciagli et al., 2014; Lüttjohann and van Luijtelaar, 2013). The epileptic oscillation triggered by stimulation of the motor cortex also possesses direct connections to the epileptic focus, indicating that VPM serves as a crucial anatomical and functional bridge between the somatosensory cortex and epileptic onset zones (FitzGerald and Folan-Curran, 2002). Previous structural MRI study has revealed cortical atrophy predominantly in fronto-central regions in TLE patients, indicating that pathology in patients with hippocampal atrophy involves the somatosensory cortex (Bernhardt et al., 2010). In the current study, we observed decreased FC between the VPM and the sensorimotor network and cerebellum in TLE patients. This finding is likely to underlie the important role of the thalamic-sensorimotor system in the behavioral manifestation of movement-related dysfunctions. Future work through specific motor task-related fMRI studies could provide additional insights into how temporal lobe epilepsy could affect the remote sensorimotor cortex and motor functions via the VPM pathway.
Dysfunctions in the thalamic arousal network in TLE have attracted broad attention in the field of neurology. Researchers hold that recurrent focal seizures in TLE may incite aberrant changes in subcortical arousal systems and eventually result in cognition deficits. Our previous resting-state fMRI study confirmed that abnormal functional alterations and disruption of information processes may occur among a number of subcortical arousal systems, including the midbrain, thalamus, and cortical regions in TLE patients (Li et al., 2019). The CL has long been implicated in arousal and is known to work cooperatively and independently by projecting excitatory innervation to activate widespread regions of the cortex, including the frontal and occipital regions (Saalmann, 2014; Schiff et al., 2012). An fMRI study on animal model of focal limbic seizures displayed decreased BOLD signals in the CL during seizures. Our recent findings based on thalamic CL neuron activity recordings further confirmed the intimate relationship between CL deactivation and suppressed cortical function (Feng et al., 2017). Here, we identified decreased FC between CL and brain clusters of the hippocampus, parahippocampus, temporal–occipital fusiform gyrus, and midbrain in patients with TLE; such a change may represent the damaging effects of recurrent seizures on the connections between the CL and widespread functional networks.
The current work has several limitations, but it lays the groundwork for additional important future studies. A major limitation of the current study is its small sample size, which may lead to a risk of type I error. In addition, we did not observe abnormal FC in the left ANT–hippocampal circuitry in new-onset patients. One possible reason is the small sample size of the new-onset patient group (n = 7). On the other hand, it is possible that long-term effects of epileptic discharges (Tong et al., 2019), seizures frequency (Ke et al., 2020), and seizure duration (Liu et al., 2016) may potentially contribute to more severe functional impairments, including hippocampal connectivity and memory impairment, which needs further confirmation in future studies with a big sample size. In addition, we assessed medication effects on thalamic FC changes in our data; however, the mechanisms of contribution of TLE-related variables, including AEDs to patients' functional changes, are largely unclear, and our findings require replication in larger homogeneous investigations. An interesting finding in the current study is the significant relationships between the anterior thalamic–hippocampal/parahippocampal FC and the memory performance in TLE patients. Thus, an important direction for future work is targeting of the anterior thalamic–hippocampal pathway via a longitudinal investigation to further elucidate the effects of focal seizures on FC and memory performance in TLE. Another important direction for future work is evaluation of different changes in thalamic connectivity after surgery. Overall, this is an important first step in recognizing the complicated connections, functions, and organization of the thalamus in patients with TLE. These findings have the potential to greatly guide future clinical treatments aimed at preventing specific abnormal thalamocortical dysfunction to restore consciousness or improve cognition deficits by transcranial magnetic stimulation or DBS treatment in TLE patients.
Conclusions
In summary, we demonstrated that TLE patients exhibited prominently decreased thalamic FC with distinct subcortical and cortical systems. Importantly, we found significant relationships between the anterior thalamic–hippocampal/parahippocampal FC and the memory performance in TLE, implying that the specific pathology of the ANT–hippocampal pathway may be a potential factor that contributes to memory deficits in patients. Overall, our findings suggest that the distinct thalamocortical FC is damaged to some extent in TLE patients and could be related to abnormal behavioral manifestations and impaired memory performance. Further understanding of these different thalamocortical circuits may lead to improved treatments directly targeting the observed modes of impaired functions.
Footnotes
Acknowledgment
The authors are grateful to all of the participants in this study.
Authors' Contributions
L.F. and R.L. conceived the study; R.L., L.Z., X.W., T.Z., H.W., C.W., and J.L. performed the analyses with guidance from H.C.; and L.F., D.G., and D.L. recruited patients and acquired the data. F.L., R.L., and H.C. provided grant support. F.L., RL., and L.Z. wrote the article with review and edits from Z.Y., B.X., and H.C. All authors commented on the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Key Project of Research and Development of the Ministry of Science and Technology (No. 2018AAA0100705), the National Natural Science Foundation of China (Nos. 61906034, 82072006, 61533006, U1808204, 81771407), and the Sichuan Science and Technology Foundation (No. 2019YFS0429).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.