
Abstract
Background:
Functional magnetic resonance imaging (fMRI) of spontaneous brain activity permits the identification of functional networks on the basis of region synchrony. The functional coupling between the elements of a neural system increases during brain activation. However, neural synchronization may also be the effect of inhibitory gamma-aminobutyric acid (GABA) neurons in states of brain inhibition such as sleep or pharmacological sedation. We investigated the effects of an oral dose of alprazolam, a classical benzodiazepine known to enhance inhibitory neurotransmission, using recently developed measures of local functional connectivity.
Methods:
In a randomized, double-blind, placebo-controlled, crossover design, 32 non-treatment-seeking individuals with social anxiety underwent two identical resting-state fMRI sessions on separate days after receiving 0.75 mg of alprazolam and placebo. Functional connectivity maps of the cerebral cortex were generated by using multidistance functional connectivity measures defined within iso-distant local areas.
Results:
Relative to placebo, increased intracortical functional connectivity was observed in the alprazolam condition in visual, auditory, and sensorimotor cortices, and in areas of sensory integration such as the posterior insula and orbitofrontal cortex (OFC). Alprazolam significantly reduced subjective arousal compared with placebo, and the change was associated with variations in multidistance functional connectivity measures in the OFC.
Discussion:
In conclusion, we report evidence that alprazolam significantly modifies neural activity coupling at rest in the form of functional connectivity enhancement within the cerebral cortex. The effect of alprazolam was particularly evident in the cortical sensory system, which would further suggest a differentiated effect of GABA inhibition on sensory processing.
Impact statement
Magnetic resonance imaging (MRI) signal synchronization between brain regions is regularly used as a measure of functional connectivity denoting activity of a network or co-activation of its elements. Less studied in the functional MRI field, however, is neural synchronization in states of brain inhibition. We mapped the effect of a pharmacological agent known to enhance gamma-aminobutyric acid inhibitory neurotransmission on the local functional structure of the cerebral cortex. We observed that alprazolam significantly increased intracortical activity coupling at rest, particularly in the cortical sensory system. The results emphasize that changes in functional connectivity measures may also reflect the synchronization effect of inhibitory neurons.
Introduction
At all levels of the nervous system, from single neurons (Tsodyks et al., 1999) and cortical columns (Arieli et al., 1995) to the whole brain (Biswal et al., 1995), there are spontaneous activity oscillations related to the functional organization of the brain (Greicius et al., 2009; Vincent et al., 2007). Based on low-frequency (below 0.1 Hz) fluctuations in the blood-oxygen level dependent (BOLD) signal, functional magnetic resonance imaging (fMRI) of spontaneous brain activity permits the identification of functional networks on the basis of region synchrony, defined as functional connectivity (Biswal et al., 1995; Fox and Raichle, 2007). Several different methodologies such as independent component analysis (ICA) and region-of-interest (ROI)-based connectivity have predominantly been used for characterizing the intrinsic architecture of large-scale brain networks. However, there is growing interest in the study of local cortical connectivity accessed through fMRI, as activity integration at this level is one of the most important organizational principles of the brain in vertebrates (Abeles, 1982; Felleman and Van Essen, 1991; Tononi et al., 1994).
To characterize the continuous smooth spatial gradient of local fMRI correlations, our group has recently described a novel imaging approach based on Iso-Distant Average Correlation (IDAC) measures (Macià et al., 2018). The IDAC measures are designed to be able to capture the average coupling of a given voxel with all its neighboring voxels as other featured connectivity methods (Sepulcre et al., 2010; Tomasi and Volkow, 2010; Zang et al., 2004). However, by targeting local functional connectivity at varying distances, this approach extends preceding efforts in the field by capturing the distance-graded change in local functional connectivity, thus providing anatomically rich representations of the dynamics of local functional connectivity. Importantly, using a display strategy consistent with the representation of three distance intervals at a time, our functional connectivity mapping was able to parcel the cerebral cortex revealing borders consistent with cytoarchitectonically distinct cortical areas (Macià et al., 2018). The degree of correspondence with classical anatomo-functional areas suggests that IDAC mapping captures primary features of the functional structure of the cerebral cortex.
A subsequent study interestingly illustrated how a pathological condition may, indeed, alter the functional structure of the cortex by differentially affecting each local distance (Pujol et al., 2019). The whole cerebral cortex was mapped using this new imaging approach in a large sample of 160 obsessive-compulsive disorder (OCD) patients and a comparative control subject group at rest (Pujol et al., 2019). The results indicated reduced functional connectivity in terms of the patients' IDAC measures in primary areas of each sensory modality, and in the orbitofrontal cortex (OFC). Moreover, symptom severity was significantly associated with the degree of functional structure alteration in OCD-relevant regions. This weaker intracortical activity coupling at rest was interpreted as a neural activity desynchronization likely to reflect deficient cortical inhibition (Pujol et al., 2019). Such results emphasize that variations in functional connectivity measures do not necessarily express variations in the number of co-activated principal neurons (Macià et al., 2018; Niessing et al., 2005), but they may also reflect the synchronization effect of inhibitory gamma-aminobutyric acid (GABA) neurons on principal neurons (Buzsáki and Watson, 2012; Mathalon and Sohal, 2015; Turkheimer et al., 2015). Therefore, a deficient GABA inhibition would result in cortical desynchronization as proposed in OCD (Pujol et al., 2019), whereas enhanced GABA inhibition would be associated with cortical synchronization (Alonso et al., 2010; Fingelkurts et al., 2004; Kiviniemi et al., 2005; Licata et al., 2013).
In the present study, we tested the effect of a pharmacological agent known to enhance the brain's GABA inhibitory neurotransmission on our IDAC measures of cortical synchronization. Thirty-five non-treatment-seeking individuals with high social anxiety scores received a single oral dose of alprazolam, a classical benzodiazepine acting as a positive allosteric modulator at GABAA receptors (Verster and Volkerts, 2004), in a randomized, double-blind, placebo-controlled, two-period crossover design. We predicted that our IDAC measures of cortical activity coupling would be altered after the administration of alprazolam as compared with placebo in the form of significant enhancement, reflecting the pharmacologically induced increase in neural inhibitory activity.
Materials and Methods
Study participants
All participants were recruited through public media advertisement (local newspapers and poster advertisements). Participants contacted the study center (Pharmacology Research Unit) by email and, subsequently, a clinical researcher would conduct a preliminary interview by telephone. A screening visit was then carried out to confirm inclusion/exclusion criteria and good physical health by means of a complete physical examination. Individuals with a Liebowitz Social Anxiety Scale (LSAS; Liebowitz, 1987) score ≥50 and aged between 18 and 64 years were eligible. Subjects with relevant medical or neurological disorder, substance abuse history, or other psychiatric illness were not included. All participants underwent a urine test for drugs of abuse and a breath alcohol test at screening to discard any pre-study positive results. Importantly, the selected subjects were not seeking treatment for their social anxiety and were not receiving any psychotherapy or course of medication that could confound the study results. From an initial sample of 35 randomized participants, 3 subjects were excluded after image preprocessing, two owing to excessive head movement during MRI acquisition and one due to image normalization artifacts and, therefore, the final sample for the main analysis included 32 participants (21 women; mean ± standard deviation [SD] [range] age = 25.9 ± 8.6 [18–52] years). Participants' mean LSAS score was 88.8 points (SD = 17.8, range = 53–126; maximum possible score 144 points). All participants were right-handed and were either studying or had completed higher education.
The study was in compliance with the Code of Ethics of the World Medical Association (Declaration of Helsinki) and was approved by the local Ethics Committee (CEIC-IMAS, Barcelona), and it was registered at EudraCT (200600421248). Written informed consent was obtained from all the participants, and all of them were compensated for their participation.
Study design
The investigation was designed as a randomized, balanced, double-blind, placebo-controlled, within-subject crossover study. All participants underwent two identical fMRI scan sessions on separate days occurring within a maximum period of 3 weeks (mean ± SD [range], 10 ± 4 days [6–21]). On 1 day, subjects received 0.75 mg oral alprazolam (Xanax® Tablets—Pfizer Ltd.; experimental condition), and on the next day an identical-appearing placebo (control condition) with complete counterbalancing, with half the subjects in this order and the other half receiving the placebo first. To ensure blinding, both alprazolam and placebo were administered in capsule form and were identical in both appearance and administration method. Subjects were instructed to refrain from tobacco, alcohol, and caffeine consumption 12 h before the fMRI session. After oral administration of alprazolam, peak plasma concentration is usually achieved within 1–2 h (Barbanoj et al., 2007; Verster and Volkerts, 2004), to ensure an appropriate exposure to the drug, the 0.75 mg dose of alprazolam or placebo was administered 1.5 ± 0.5 h before fMRI acquisition.
Behavioral assessment
On both MRI scanning days, immediately before the fMRI scan, subjects were requested to fill in the state portion of the Spielberger State-Trait anxiety inventory (STAI-S; Spielberger et al., 1983). In addition, four 0–100 mm visual analogue scales (VAS) were administered to participants at each test time to evaluate changes in subjective anxiety and arousal states that might result from benzodiazepine administration. The scales used represented a continuum along a 100 mm horizontal line with 0 on the left and 100 on the right between the pairs of extremes as follows: alert-drowsy, calm-excited, sleepy-awake, and tense-relaxed. To avoid susceptibility to left-right response bias, a counterbalance design was used with two VAS marking the most negative connotation for a specific aspect on the leftmost side of the line and the most positive connotation on the rightmost side (sleepy [0]—awake [100], tense [0]—relaxed [100]), whereas in the other two VAS the opposite pattern was presented (alert [0]—drowsy [100], calm [0]—excited [100]). A new variable was calculated from the average of the four VAS scores to give a “global measurement of the arousal state,” including the counterbalance of responses.
MRI acquisition
Participants were assessed by using a 1.5-T Signa Excite system (General Electric, Milwaukee, WI) equipped with an eight-channel phased-array head coil and single-shot echo-planar imaging software. Functional sequences consisted of gradient recalled acquisition in the steady state (repetition time, 2000 ms; echo time, 50 ms; and pulse angle, 90°) in a 24-cm field of view, with a 64 × 64 pixel matrix and a slice thickness of 4 mm (interslice gap, 1.5 mm). Twenty-two interleaved slices, parallel to the anterior–posterior commissure line, were acquired to generate 180 whole-brain volumes (total duration of 6 min). The first four (additional) images in each run were discarded to allow magnetization to reach equilibrium. Participants were instructed to simply relax, stay awake, and lie still, while keeping their eyes open throughout the session.
We also acquired a high-resolution T1-weighted anatomical image for each subject by using a three-dimensional (3D) fast spoiled gradient inversion-recovery prepared sequence. A total of 130 contiguous slices were acquired with repetition time 11.8 ms, echo time 4.2 ms, flip angle 15°, field of view 30 cm, 256 × 256 pixel matrix, and a slice thickness of 1.2 mm.
MRI preprocessing
Anatomical and functional images were all visually inspected before analysis to detect possible acquisition artifacts. Imaging data were processed by using MATLAB version 2014b (The MathWorks, Inc., Natick, MA) and Statistical Parametric Mapping software (SPM12; The Wellcome Department of Imaging Neuroscience, London).
As in our previous study (Macià et al., 2018), fMRI images were slice-time corrected and realigned to their mean (motion-corrected). Image volumes were then coregistered to their anatomical images with an affine transformation using SPM and smoothed by means of a 4 × 4 × 4 mm3 full width at half maximum Gaussian kernel. A warping matrix was also estimated for every subject to match a group template created from the 3D anatomical individual acquisitions and then normalized to the Montreal Neurological Institute (MNI) space using DARTEL (see IDAC maps below). Finally, image volumes were resliced to 3 × 3 × 3 mm resolution.
Filtering and denoising of data
All fMRI time series were band-passed with a Discrete Cosine Transform filter letting through frequencies in the 0.01–0.1 Hz interval. We regressed all time series on the 6 rigid body realignment parameters and their first-order derivatives, and on the average white matter, CSF, and global brain signals extracted from the native tissue masks.
Control of potential head motion effects
Participants were excluded from analyses when showing evident head displacements throughout the course of the scan (translations >3 mm in any of the x, y, or z directions, rotations >3°) or residual motion artifacts in the first-level image (typically appearing in tissue boundaries, brain ventricles, and white matter). In addition, within-subject, censoring-based MRI signal artifact removal (scrubbing) (Power et al., 2014) was used to discard motion-affected volumes using affine realignment parameters obtained during image preprocessing. For each subject, inter-frame motion measurements (Pujol et al., 2014) served as an index of data quality to flag volumes of suspect quality across the run. At points with inter-frame motion >0.3 mm, the corresponding volume, the immediately preceding volume and the two successive volumes were all discarded. Using this procedure, the average number of censored volumes per acquisition was 1% (SD, 2%, range, 0–19%). Censored volumes in excluded cases were 36 in one participant and 9 in another participant, who additionally showed translations >3 mm.
IDAC maps
Whole-brain IDAC maps were then generated for each subject by estimating the average temporal correlation (mean correlation z-score) of each voxel's fMRI signal and the fMRI signal of all its neighboring voxels placed at increasingly separated iso-distant intervals. Proof-of-concept and sensitivity analyses are extensively described in our early report, as well as precise image processing procedures (Macià et al., 2018). In short, the estimation of IDAC values for each voxel was carried out on all the subjects' native space within a gray matter mask separately for each hemisphere. The two hemispheres were brought back together once the IDAC values had been calculated. Estimated DARTEL normalizations (Ashburner, 2007) to the MNI space obtained in the image preprocessing steps were finally applied to the IDAC results to enable group inferences.
Three IDAC maps were obtained at the distance intervals 5–10, 15–20, and 25–30 mm. Multidistance IDAC color maps were obtained from the overlay of IDAC values from the different intervals by using a red, green, and blue (RGB) color codification, enabling the display of three values at the same time. RED was chosen to display results from 5 to 10 mm IDAC map analyses, GREEN from 15 to 20 mm, and BLUE from 25 to 30 mm. In this way, the resulting maps show a full range of secondary colors that are informative of variations in the functional structure of the cerebral cortex in both the comparison between alprazolam and placebo and correlations.
Statistical analysis
Behavioral analysis
Requirement for parametrical statistical tests was tested in all cases. STAI-S and VAS scores under both placebo and alprazolam conditions were compared by means of the parametric paired Student t-test. All analyses were performed by using SPSS Statistics v20.0.
Imaging analysis
Single-subject IDAC connectivity scores were carried forward to subsequent group-level linear models in SPM adopting a 2 × 3 design, factorial model (treatment [alprazolam, placebo] by distance [5–10, 15–20, and 25–30 mm]). Alprazolam and placebo conditions did not significantly differ in terms of total motion (mean ± SD, 0.4 ± 0.3 mm alprazolam; 0.3 ± 0.2 mm placebo; t = 2.87, p = 0.262). However, a small but significant difference was found for the mean interframe motion parameter (0.05 ± 0.02 mm alprazolam; 0.03 ± 0.01 mm; t = 2.50, p = 0.018) and this measure was included as a covariate in the analysis. We tested for bidirectional treatment differences across the 3 distances (Alprazolam > Placebo and Alprazolam < Placebo). Differences in connectivity were then estimated separately for each of the 3 IDAC maps at distances 5–10, 15–20, and 25–30 mm.
In addition, correlation analyses were performed to map the possible relationships between clinical measurements (STAI-S, VAS scores, and global arousal, separately) and whole-brain functional connectivity for each of the 3 IDAC maps. To identify the brain areas in which the magnitude of the connectivity changes by treatment was related to the change in the subjective anxiety and arousal states, we estimated voxel-wise the correlations between functional connectivity changes (i.e., alprazolam IDAC values minus placebo IDAC values) and behavioral changes (alprazolam STAI-S and VAS scores minus placebo STAI-S and VAS scores) by using linear regression in SPM. For each subject, “difference” images were computed over alprazolam and placebo conditions at each distance and the images were used to map the functional connectivity changes in the whole sample by means of one-sample t-tests.
Finally, a control analysis was performed to test the extent to which the social anxiety condition by itself was associated with variations in our cortical connectivity measures. The primary sample of 32 participants in the placebo condition was compared with an independent sample of 24 reference healthy volunteers with equivalent age, educational status, laterality, and gender distribution (14 women p = 0.78; mean ± SD [range] age = 27.0 ± 7.9 [21–44] years; p = 0.63), who underwent an identical assessment with a single fMRI scan session without treatment. A 2 × 3 factorial model in SPM (group [anxiety, control] by distance [5–10, 15–20, and 25–30 mm]) was used to compare IDAC measures.
Thresholding criteria
Results were considered significant when clusters formed at a threshold of p < 0.005 survived whole-brain family-wise error (FWE) correction (p < 0.05) calculated by using SPM.
Results
Behavioral results: STAI-S and VAS scores
On both MRI scanning days (alprazolam and placebo), immediately before the fMRI examination (peak drug effect), STAI-S and VAS scores were collected to estimate subjective anxiety and arousal levels. Alprazolam showed a tendency to reduce STAI-S scores compared with the placebo condition ([percentiles, mean ± SD] alprazolam: 41.2 ± 24, placebo: 46.9 ± 25; t = 1.76, p = 0.088) and significantly reduced general arousal (“global measurement of the arousal state” variable; alprazolam: 38.3 ± 14, placebo: 46.9 ± 16; t = −3.06, p = 0.005). We found significant effects of alprazolam in 3 out of the 4 administered VAS when tested separately. Specifically, the VAS statistics for each dimension were: (1) alert-drowsy: alprazolam = 54.0 ± 19, placebo = 45.5 ± 18, t = −2.25, p = 0.032 (alprazolam significantly increased drowsiness levels); (2) calm-excited: alprazolam = 30.0 ± 18, placebo = 38.0 ± 20, t = 2.22, p = 0.034 (alprazolam significantly increased calm sensation); (3) sleepy-awake: alprazolam = 43.9 ± 19, placebo = 56.4 ± 22, t = 2.40, p = 0.023 (alprazolam significantly increased the sensation of sleepiness); and (4) tense-relaxed: alprazolam = 66.9 ± 19, placebo = 61.3 ± 22, t = −1.66, p = 0.108. Changes in STAI-S and VAS measures are illustrated in Supplementary Figure S1.
Functional connectivity using IDAC measures
As an illustration of IDAC brain mapping, we produced a group-averaged map from the resting-state acquisition of the 24-participant reference group receiving no-treatment. IDAC functional connectivity measures at three different distances were overlaid together (multidistance) on the cortical surface by using an RGB color codification (Supplementary Fig. S2). Using this display strategy, IDAC maps were able to reveal a degree of brain parcellation into major anatomo-functional regions, which is consistent with our previous studies (Macià et al., 2018; Pujol et al., 2019). For instance, the visual cortex was distinguished from the temporal and parietal cortices and the perirolandic cortex from parietal and prefrontal areas. The angular gyrus was jointly identified with the supramarginal gyrus and the motor cortex with the somatosensory cortex.
Differences in IDAC measures between alprazolam and placebo
A factorial model was used to test whether there were any significant differences between alprazolam and placebo conditions in IDAC functional connectivity measures across the 3 distances (Effect of Treatment). Participants showed stronger IDAC functional connectivity in the alprazolam condition (alprazolam > placebo) bilaterally in ventral and lateral visual areas, the somatosensory cortex extending to the motor cortex and medial frontal cortex, auditory cortex and related areas of the temporal lobe, posterior insula, and OFC (Fig. 1; Table 1; Supplementary Figs. S3 and S4). No significant results were found for the opposite contrast alprazolam < placebo.
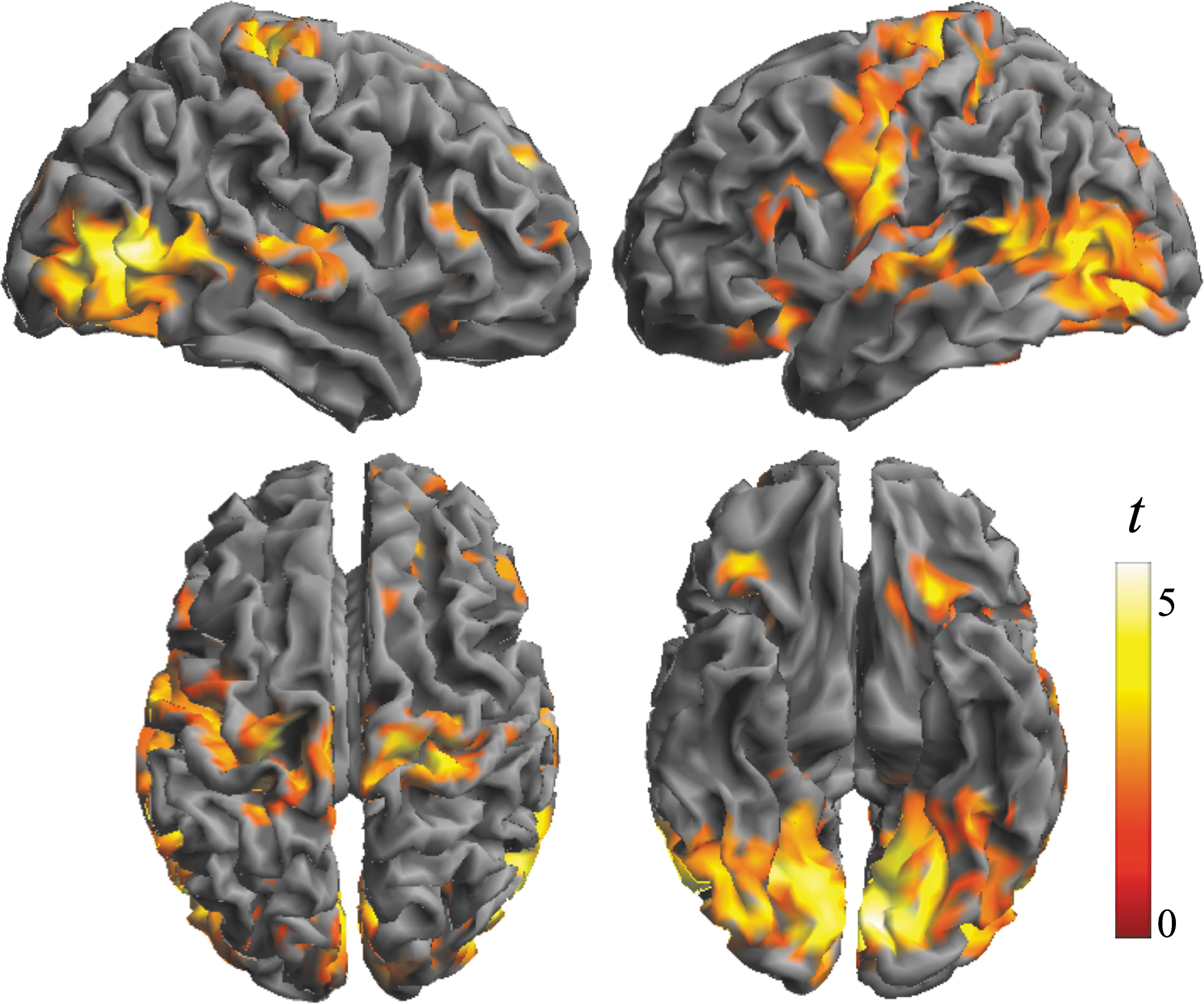
Differences in the local architecture of functional connectivity as assessed with IDAC measures between alprazolam and placebo (effect of treatment). The brain views illustrate significant findings obtained in the contrast Alprazolam > Placebo (positive treatment effect—stronger functional connectivity). Top row: lateral view of right and left hemispheres. Bottom row: dorsal and ventral surfaces. Color bar represents t value. IDAC, Iso-Distant Average Correlation.
Differences between Alprazolam and Placebo in Iso-Distance Average Correlation Functional Connectivity Measures Across the 3 Distances (5–10, 15–20, and 25–30 mm)
Same cluster.
PFWE-corr, P (Family-Wise Error corrected); SMA, supplementary motor area; x y z, coordinates given in Montreal Neurological Institute space.
Differences in local functional connectivity were then estimated separately at each distance (5–10, 15–20 and 25–30 mm; Fig. 2). The pattern of findings reproduced the results of the combined analysis, but it identified some differences across the maps. In general, the alprazolam connectivity enhancement effect was more notable in sensory areas at short and mid-distances. The results of the analysis of variance interaction test between short (5–10 mm) and long (25–30 mm) distance and treatment confirmed such a distance effect (difference at short distance > difference at long distance) in the sensorimotor cortex and ventral and lateral aspects of the visual cortex. By contrast, in association areas of the prefrontal lobe and parieto-occipital junction, the increase in IDAC values was significantly greater at long versus short distance (difference at short distance < difference at long distance; Supplementary Fig. S5; Supplementary Table S1). Figure 3 provides an integrated RGB display of the 3 distance map analyses to summarize the effect of alprazolam on the functional structure of the cerebral cortex.
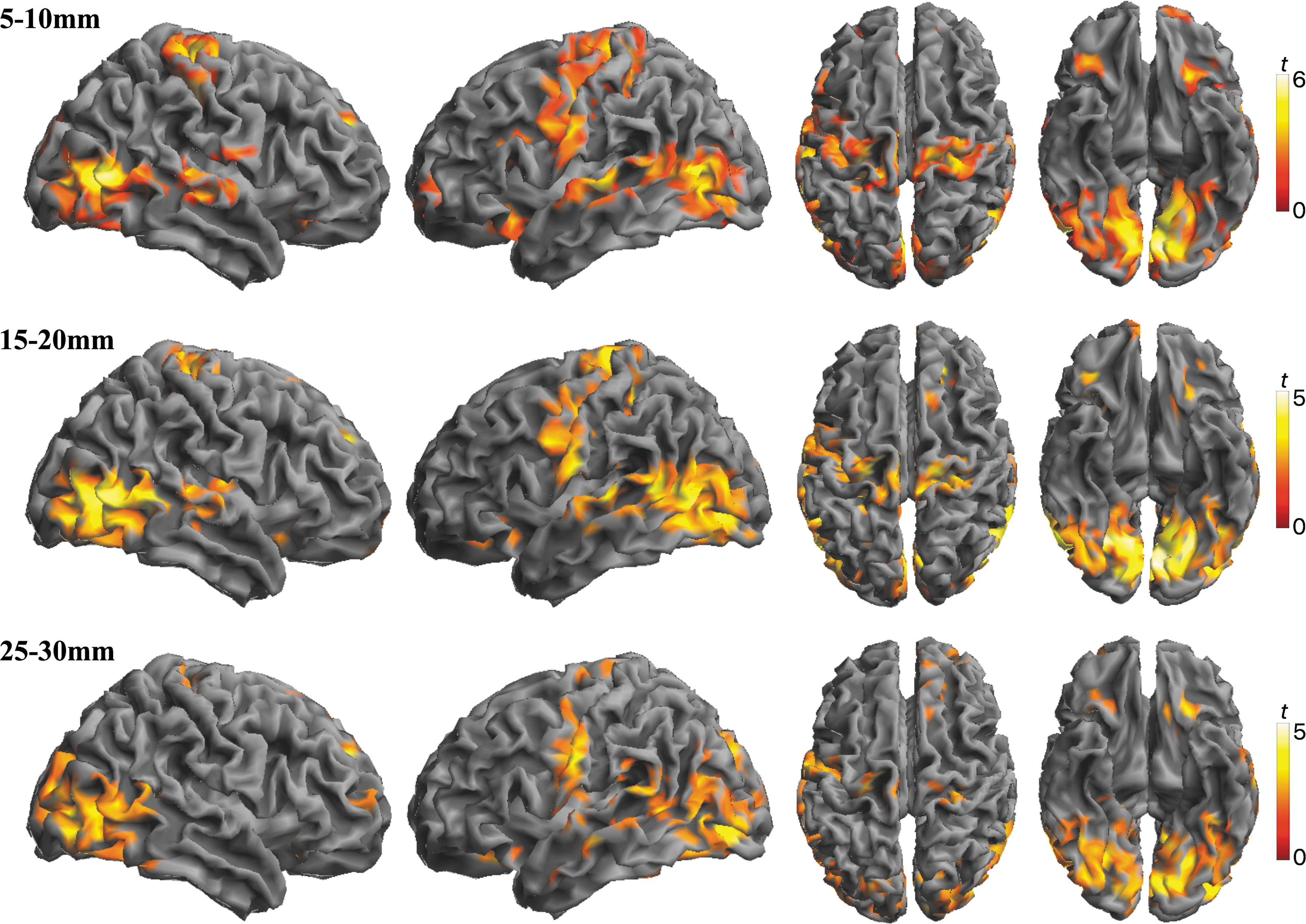
Differences in IDAC measures between alprazolam and placebo (contrast Alprazolam > Placebo) at 3 functional connectivity distances: 5–10, 15–20, and 25–30 mm. Alprazolam showed higher functional connectivity measures in the 3 analyses. However, the effect was not identical for each distance. Data are displayed on the lateral, dorsal, and ventral surfaces. Color bars represent t values.

RGB display summarizing alterations in the local architecture of functional connectivity under alprazolam relative to placebo. The color map corresponds to the superimposition of within-group differences (contrast Alprazolam > Placebo) in IDAC measures at functional connectivity distances 5–10 mm (red), 15–20 mm (green), and 25–30 mm (blue). That is, the RGB combination of the 3 sets of results is shown in Figure 2. The final output is made up of primary RGB colors and their secondary combinations. Color bars represent t values. RGB, red, green, and blue.
Correlations between IDAC measures and STAI and VAS scores
In the analysis of correlations, no significant results were demonstrated at whole-brain FWE correction between functional connectivity changes (i.e., alprazolam IDAC values minus placebo IDAC values) and changes in STAI-S and VAS scores (alprazolam score minus placebo score). Subthreshold (p < 0.01, cluster size >1.4 mL), a correlation was found between VASdrowsy–alert and VASsleepy–awake score reduction and functional connectivity increase in a region extending from the OFC to the medial prefrontal cortex across the 3 IDAC distance maps (Fig. 4; Supplementary Table S2). Findings were observed in the direction of a greater decrease in self-reported arousal scores associated with a greater increase in functional connectivity measures.
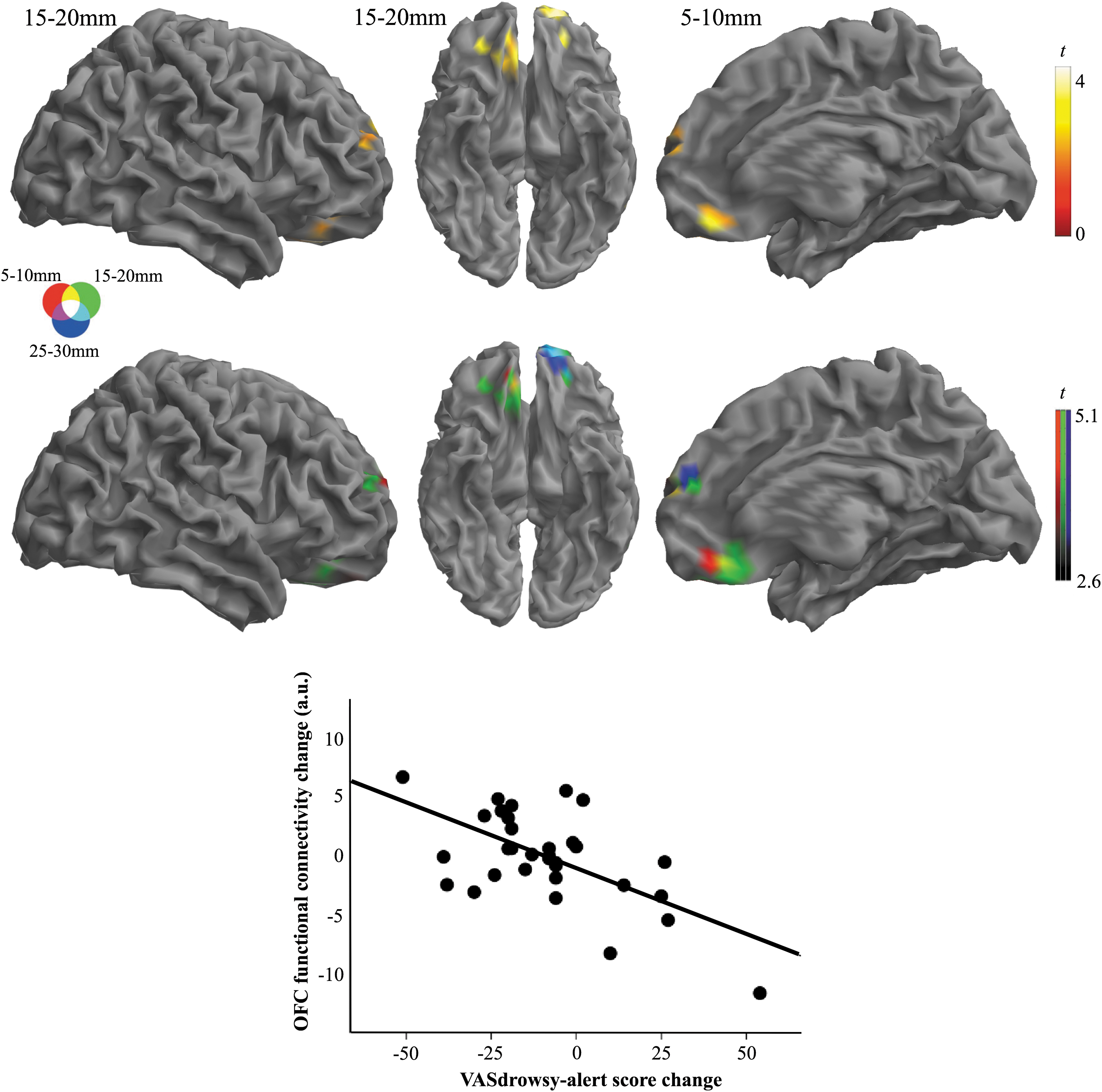
Correlation between changes in VAS scores for alertness (alprazolam score minus placebo score) and IDAC functional connectivity changes (alprazolam IDAC values minus placebo IDAC values). Top row: regions showing significant inverse correlation between VAS score reduction and connectivity increase in different IDAC maps. Middle row: results obtained in separate maps superimposed by using RGB display. Data are displayed on the right lateral, ventral, and medial cortical surfaces. Color bars represent t values. Bottom: The scatter plot illustrates the correlation between self-reported VAS score change and OFC functional connectivity change (R 2 = 0.38, p < 0.001) at coordinates x = 7, y = 36, and z = −21 for the IDAC distance 15–20 mm. OFC, orbitofrontal cortex; VAS, visual analogue scale.
Testing the effect of social anxiety condition on IDAC measures
A control analysis was performed to test whether the social anxiety condition itself was associated with variations in our cortical connectivity measures. We found no significant results when comparing the primary sample of 32 participants in the placebo condition with an independent sample of 24 reference healthy volunteers receiving no treatment.
Discussion
In the present study, we investigated the effects of an oral dose of alprazolam, a classical benzodiazepine, using a multidistance measure of functional connectivity in a randomized, double-blind, placebo-controlled, cross-over design. Our findings revealed that acute administration of alprazolam increased intracortical activity coupling at rest. Specifically, an increase in local functional connectivity was identified in primary visual, auditory, and sensorimotor cortices, and in areas of sensory integration in the occipito-temporal, posterior insula, and orbitofrontal cortices. A significant distance effect was observed, with a different degree of alteration in several areas in the short, middle, and long-distance maps. Subjective arousal reduction in the participants was associated with the degree of functional connectivity changes, specifically in the OFC.
In accordance with the nonspecific, dose-related, sedative effects of benzodiazepines (Lader, 2011), the administration of a single dose of 0.75 mg of alprazolam produced measurable subjective changes in the participants with a reduction of self-reported ratings of anxiety and general arousal relative to placebo. Changes in the VAS indicated that alprazolam significantly increased the level of drowsiness, calm and sleepy sensations, and showed a tendency to increase relaxation, which is in agreement with results from previous studies using alprazolam (Barbanoj et al., 2007; De Witte et al., 2002; Reissig et al., 2015; Roy-Byrne et al., 1993; Streeter et al., 1998) in terms of subjective sedation (Lader, 2011; Verster and Volkerts, 2004).
Alprazolam facilitates GABAergic neurotransmission by potentiating GABA-induced chloride flux at the GABAA-benzodiazepine receptor complex (Verster and Volkerts, 2004). Although GABA receptors are widely expressed throughout the brain (Andersson et al., 2019; Brown et al., 2011; Zilles et al., 2004), neuro-anatomical and histological studies have demonstrated a heterogeneous distribution with particularly high densities of GABAA-benzodiazepine binding sites in visual cortical areas, somatosensory and motor cortices, hippocampus, basal ganglia, thalamus, brainstem, and also in the OFC (Faull and Villiger, 1988; Fioravanti et al., 2019; Northoff et al., 2002; Puts et al., 2018; Young and Chu, 1990; Zilles et al., 2002). As the distribution of GABAA receptors varies across the brain, it is conceivable that the effect of alprazolam on functional connectivity varies with the local receptor concentration. As seen in Figures 1–3, only specific brain regions showed significant connectivity changes in our study. In particular, we observed that alprazolam affected the local connectivity in areas primarily or secondarily related to sensory processing and motor function. We also found significant changes in the OFC, which has a role in both sensory integration (Rolls, 2019a,b) and emotional experience (Milad and Rauch, 2007; Northoff et al., 2002). Interestingly, a relationship was observed between the increase in functional connectivity and subjective arousal reduction in the OFC. Behavioral studies support the existence of a relevant effect of alprazolam on sensory, motor, and emotional processing (Verster and Volkerts, 2004). Our pattern of findings is, therefore, in accordance with a differential behavioral effect of alprazolam on these systems.
The alprazolam-induced increase in functional connectivity demonstrated here using multidistance IDAC measures is consistent with results from previous studies investigating the effects of different sedative-type compounds on the resting-state BOLD signal based on other methodological approaches. For example, increased functional connectivity within sensory-related networks during light sedation with midazolam, another benzodiazepine, has been reported using both ICA of resting-state networks and ROI-based fMRI analysis (Greicius et al., 2008; Kiviniemi et al., 2005; Liang et al., 2015). Similarly, an acute oral dose of zolpidem, a non-benzodiazepine hypnotic, enhanced functional connectivity within sensory, motor, and limbic networks identified by using resting-state ICA (Licata et al., 2013). Compared with placebo, and using networks-of-interest, alcohol administration significantly increased the overall connectivity strength of the visual network, with the primary visual cortex showing the most prominent alcohol-induced changes (Esposito et al., 2010; Khalili-Mahani et al., 2012). By contrast, decreased resting-state functional connectivity has been observed after the administration of a single dose of caffeine in the motor and visual cortices (Rack-Gomer et al., 2009; Wu et al., 2014).
Beyond merely substantiating results from previous research, our data are uniquely detailing the functional anatomy and specificity of the alprazolam effect on local functional connectivity. At the typical alprazolam dosage used in our study, neural synchronization was almost selectively observed in sensory cortices. Importantly, we also observed a level of selectivity within the sensory areas. Indeed, the effect within sensory areas was not general, but it differed across local distances. All in all, the functional anatomy of our results may indicate that the inhibitory neural elements are distributed according to a defined pattern consistent with the sensory cortex “functional structure” defined by local IDAC measures.
In the behavioral context, the notable selectivity of alprazolam on sensory cortices suggests that the sedative effects of alprazolam at relatively low doses may be achieved, at least in part, via a reduction of sensory-related neural arousal. On the other hand, sensory processing dampening may potentially contribute to the common benzodiazepine adverse effects related to decreased alertness (e.g., reducing driving skills).
The inhibitory activity of alprazolam has been demonstrated in neurophysiological studies, and its effects on electroencephalography (EEG) are characteristic of benzodiazepines in that they cause a decrease in cortical alpha and an increase in beta activity (Allen et al., 1991; Barbanoj et al., 2007), generally associated with such inhibitory states. On the other hand, studies recording event-related potentials have shown that alprazolam administration led to significantly reduced N1 and P1 amplitude, which are believed to reflect the inhibitory influence on stimulus-induced cortical arousal (Allen et al., 1991; Bond et al., 1992; Premoli et al., 2014; Semlitsch et al., 1995), and they also gave rise to significantly decreased P300 amplitude and increased P300 latency, thus reflecting a reduction in information processing and prolonged stimulus evaluation time, respectively (Münte et al., 1996; Semlitsch et al., 1995). In line with our findings, pharmaco-EEG studies investigating the consequences of inhibition have also documented an increase in EEG connectivity after the administration of alprazolam (Alonso et al., 2010) and other benzodiazepines (Fingelkurts et al., 2004), showing GABA signaling to reorganize the dynamics of local neuronal populations and GABAergic inhibition to be a powerful mechanism of neuronal synchronization (Fingelkurts et al., 2004).
MRI signal synchronization between brain regions is regularly used as a measure of functional connectivity denoting activity of a network or co-activation of its elements. Similarly, lower connectivity is generally associated with decreased activity. Nevertheless, this association is not that straightforward insofar as variations in functional connectivity measures do not necessarily express variations in the number of co-activated principal neurons (Macià et al., 2018; Niessing et al., 2005), but may also reflect the synchronization effect of GABA neurons on principal neurons (Buzsáki and Watson, 2012; Mathalon and Sohal, 2015; Turkheimer et al., 2015). Therefore, a deficient GABA inhibition can result in cortical desynchronization (Pujol et al., 2019) and the reverse also holds true for the association of lower activity with higher connectivity, which is well established in the EEG literature (Fingelkurts et al., 2004), but less studied in the fMRI field. Our study adds to existing research by emphasizing that neural synchronization is also possible in states of brain inhibition.
Limitations
The interpretation of fMRI functional connectivity can be complicated by the dependence of the BOLD signal on both neural and vascular factors (Liu, 2013). Whole-brain cerebral blood flow (CBF) and regional cerebral blood volume have been found to decrease in response to acute alprazolam administration (Roy-Byrne et al., 1993; Streeter et al., 1998; Verster and Volkerts, 2004). Similarly, physiological factors, not considered in our study, such as cardiac and respiratory fluctuations are potentially capable of modulating resting-state connectivity, which may represent a limitation in the interpretation of the present results. However, alprazolam did not affect mean ventilation or heart rate in healthy individuals (Carraro et al., 2009), and other studies using pharmacological agents also demonstrating a decrease in CBF, such as caffeine, have found opposite effects (i.e., decreases) on functional connectivity that were not correlated with the observed changes in baseline CBF (Rack-Gomer et al., 2009), which does not support the vascular origin of our findings. Controlling these possible confounding variables would be useful in future studies for a more accurate interpretation of connectivity changes.
Also, alprazolam was administered at 0.75 mg to obtain mild-to-moderate sedative effects within the range of clinical use. Therefore, the present results apply to this dose but may not be generalizable to all dosage regimes. We assume that the modulatory effect of alprazolam on the different brain systems could change with either a lower or higher dosage, producing different results.
Conclusion
We report evidence that alprazolam significantly alters intracortical neural activity coupling at rest. Our previous study using IDAC measures demonstrated neural activity desynchronization in OCD patients, a population exhibiting deficient neuronal inhibition. Accordingly, we now show the opposite in a pharmacologically induced inhibitory state. The direction of the alteration is also consistent with previous research using other sedative agents. However, by using a multidistance functional connectivity measure, we go beyond identifying the anatomical location of changes, and also provide details as to the pattern of alterations in the functional structure of cortical connections at varying local distances. The effect of alprazolam was particularly evident in the cortical sensory system, which would further suggest a differentiated effect of GABA inhibition on sensory processing. It may be of interest for future studies to disentangle how the different behavioral effects of alprazolam relate to the connectivity changes observed in our study.
Footnotes
Acknowledgments
The Agency of University and Research Funding Management of the Catalonian Government participated in the context of Research Groups SGR2017/1198 and SGR2017/134.
Authors' Contributions
J.P. and J.D. contributed to the study design, and L.B.H., J.P., G.M.V., and J.D. to data acquisition and analysis; D.M. developed fMRI analysis tools; and L.B.H. and J.P. contributed to drafting the article. G.M.V., J.D., D.M., R.M.S., and V.P.S. provided a critical revision of the article for important intellectual content. All authors have revised and approved the article for publication.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was partly supported by the Institute of Health Carlos III-CIBER, Government of Spain, reference number C1589923, and Centers of Excellence for Drug Discovery (CEDD), GSK, grant number CRH108751.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.