
Abstract
Background:
The corpus callosum (CC) plays a key role in mediating interhemispheric connectivity and developing functional hemispheric asymmetries. The purpose of this study was to investigate the changes in CC microstructure accompanying interhemispheric language reorganization in patients with brain arteriovenous malformations (AVMs).
Methods:
Forty-one patients with an unruptured AVM located in anatomically defined language areas underwent functional magnetic resonance imaging and diffusion tensor imaging. Hemispheric dominance in Broca's area (BA) and Wernicke's area (WA) was assessed separately. Right-sided or bilateral language dominance was classified as atypical lateralization. The CC was segmented into five subregions, and the mean fractional anisotropy (FA) was extracted. The relationship between callosal FA and language lateralization patterns was statistically analyzed.
Results:
We observed atypical language lateralization in 16 (39.0%) patients. Patients with atypical lateralization exhibited significantly higher mean FA values in the total CC (p = 0.002) and the anterior (p = 0.047), midanterior (p = 0.001), and midposterior (p = 0.043) subregions. Significant interaction effects of BA and WA lateralization were found for FA values in the total CC (p = 0.005) and the midanterior subregion (p = 0.004).
Conclusions:
These results indicate that AVM patients with atypical language lateralization exhibit higher callosal FA values, reflecting greater interhemispheric connectivity. Our findings contribute additional insights into the understanding of functional and structural plasticity of the human brain under pathological states.
Impact statement
Brain arteriovenous malformations (AVMs) are congenital lesions that frequently lead to interhemispheric language reorganization. In this study, by combining diffusion tensor imaging and functional magnetic resonance imaging, we investigated the relationship between callosal fractional anisotropy (FA) and language reorganization in patients with AVMs. We found that callosal FA was significantly higher in patients with atypical language lateralization, especially in those with crossed lateralization of Broca's and Wernicke's areas. This study demonstrated the remodeling of the corpus callosum microstructure accompanying language reorganization in AVM patients, providing insights into the structural and functional plasticity of the human brain associated with congenital cerebrovascular disease.
Introduction
Brain arteriovenous malformations (AVMs) are generally considered congenital lesions, often causing abnormalities in brain blood perfusion and dysfunction of adjacent brain tissue (Solomon and Connolly, 2017). Cortical functional reorganization has been frequently reported in AVM patients with eloquent areas involved (Alkadhi et al., 2000; Vikingstad et al., 2000). The affected eloquent cortex can be shifted to nearby sites or to homologous regions of the contralateral hemisphere. Using functional magnetic resonance imaging (fMRI), Deng et al. (2015) reported a rather high incidence (36.5%) of right-sided atypical language lateralization in patients with left-hemispheric AVMs.
It is believed that structural connections are the physical substrates of functional interactions across the brain (Zhang et al., 2011). Given that the corpus callosum (CC) is the largest fiber tract connecting the left and right hemispheres, there may be changes in callosal organization accompanying interhemispheric language reorganization in AVM patients. Prior studies have revealed the critical role of the CC in the development of language lateralization (Hinkley et al., 2016), but the concept of callosal function is currently divided into two conflicting models: the inhibitory model and the excitatory model (van der Knaap and van der Ham, 2011). Using fMRI, Josse et al. (2008) found that individuals with larger callosal size exhibited stronger left lateralization of language activation. Diffusion tensor imaging (DTI) is sensitive to subtle changes in white matter microstructure, and fractional anisotropy (FA) has been shown to be a more accurate approach than callosal size for quantifying interhemispheric connectivity (Barbeau et al., 2015; Chepuri et al., 2002; Putnam et al., 2008). Combining DTI and fMRI, Putnam et al. (2008) found that anterior callosal FA values were negatively correlated with language activation of the right homologous Broca's area (BA). These results support the inhibitory model, indicating an inverse relationship between interhemispheric connectivity and the language contribution of the nondominant hemisphere. Conversely, Haberling et al. (2011) reported significantly higher callosal FA values in subjects showing bilateral or right-sided language lateralization than in those showing typical left-sided lateralization. The excitatory model suggests that greater interhemispheric communication is present in individuals with the nondominant hemisphere involved in language function.
Recently, several studies have suggested that the macro- and microstructure of the CC could be remodeled to accommodate interhemispheric language reorganization in specific disease states, including brain tumors (Cho et al., 2018; Tantillo et al., 2016; Wood et al., 2008). Compared with brain tumors, AVMs have a much more chronic disease course and produce effects on the brain much earlier in life, affording more time for language reorganization to occur. Deng et al. (2015) found that brains with AVMs have a greater capacity for language reorganization than brains with glioma. However, there is a paucity of studies investigating the accompanying changes in the callosal microstructure in the presence of AVMs. Characterizing the relationship between callosal FA and language lateralization will help us understand brain plasticity associated with AVMs.
We hypothesized that the CC plays a role in interhemispheric language reorganization associated with brain AVMs. Here, we used DTI to calculate callosal FA values and fMRI to determine language lateralization, aiming to investigate the changes in CC microstructure accompanying language reorganization in patients with brain AVMs.
Materials and Methods
Subjects
This study was approved by the Institutional Review Board of Beijing Tiantan Hospital, Capital Medical University. All subjects provided written informed consent before participation. Between June 2016 and December 2019, 41 patients with brain AVMs were extracted from a prospective clinical trial (ClinicalTrials.gov, identifier: NCT02868008). All patients were native Chinese speakers and right-handed. All lesions were located in the perisylvian areas of the left hemisphere, overlaying or adjacent to the BA (the pars opercularis and pars triangularis of the inferior frontal gyrus) or Wernicke's area (WA, the supramarginal, angular, and the posterior portion of the superior and middle temporal gyrus). Patients were excluded if they met any of the following criteria: (1) history of intracranial hemorrhage, (2) apparent perifocal edema on T2-weighted images, (3) apparent space-occupying effect, or (4) lesions involving the CC. Clinical data collected included patient demographics, clinical presentation, and lesion characteristics.
Image acquisition
All subjects underwent T1-weighted imaging, T2-weighted imaging, fMRI, and DTI examination. All MRI scans were performed with a 3.0T MR system (Magnetom Trio, Siemens Healthineers, Erlangen, Germany). The scanning parameters were as follows: T1-weighted images were acquired with a three dimensional magnetization-prepared rapid gradient-echo sequence, with time of repetition (TR) = 2300 ms, time of echo (TE) = 2.98 ms, flip angle = 9°, field of view (FOV) = 240 × 256 mm, matrix size = 240 × 256, slice thickness = 1 mm, voxel size = 1 × 1 × 1 mm, and 176 sagittal slices. Functional images were acquired with an echo-planar-imaging (EPI) sequence, with TR = 3000 ms, TE = 30 ms, flip angle = 90°, FOV = 210 × 210 mm, matrix size = 64 × 64, slice thickness = 4 mm, voxel size = 3.3 × 3.3 × 4 mm, and 30 slices. DTIs were acquired with an EPI sequence, with b value = 1000 along 30 diffusion directions, TR = 6800 ms, TE = 93 ms, flip angle = 120°, FOV = 230 × 230 mm, matrix size = 128 × 128, slice thickness = 2.8 mm, voxel size = 1.8 × 1.8 × 2.8 mm, 50 slices, double averaging, and parallel acquisition (generalized autocalibrating partially parallel acquisition [GRAPPA] factor = 2).
Functional task
In our institution, patients were tested with two fMRI paradigms, a verb-generation task (Benson et al., 1999; Paulesu et al., 1993) and a picture-naming task, for preoperative evaluation of language function. Verbal fluency tasks have been historically used as the gold standard for determining language lateralization with fMRI because of their reliability and robustness for assessing both frontal and temporoparietal areas (Bradshaw et al., 2017; Rowan et al., 2004; Rutten et al., 2002). Therefore, data from the verb-generation task were used to calculate the language laterality index. The verb-generation task was block-designed, consisting of eight 21-sec baseline blocks and eight 15-sec task blocks. During the task period, five common concrete nouns (written) were presented to subjects at a rate of one every 3 sec. Subjects were instructed to generate a verb associated with each noun silently (e.g., books = read). During the control period, subjects were asked to fix their gaze on a pound sign (#) presented in the center of the screen. Before scanning, all subjects were carefully trained to ensure that they had the ability to complete the tasks.
Lesion mapping
We manually outlined the boundaries of AVMs on anatomical T1-weighted images slice by slice using MRIcron software. Next, using the Clinical Toolbox, the lesion masks were transformed into the international consortium for brain mapping (ICBM)-152 standard space of the Montreal Neurologic Institute (MNI) and overlapped on the ICBM-152 template for display. The normalized volume of the lesions was estimated, and the overlapping areas of the lesion and language region of interest (ROI) were extracted. The percentage of the language area involved by a lesion was calculated as the volume of overlapping area divided by the volume of the corresponding language ROI.
fMRI processing
fMRI data were processed using SPM 12 on MATLAB 2017b (The MathWorks, Inc., Natick, MA). Functional images were spatially realigned to the first image in the time series and were coregistered to the T1-weighted anatomical images. Then, the functional images were normalized to the MNI space and smoothed with an isotropic 6-mm full-width at half-maximum Gaussian kernel. Importantly, to overcome possible deformation caused by the signal abnormalities of focal lesions, we used the enantiomorphic method (Nachev et al., 2008) for normalization using the Clinical Toolbox (Rorden et al., 2012). The enantiomorphic method is supposed to yield better transformation results than the cost function masking method (Brett et al., 2001) for normalization of brains with focal lesions. First, mirrored T1-weighted images were created and coregistered to native T1-weighted images. Next, an intact chimeric anatomical image was created based on the actual T1-weighted image with the lesion area replaced by normal tissue from the homologous area of the contralateral hemisphere (the smoothed lesion mask was used for inserting tissue). Then, the unified segmentation normalization method (Ashburner and Friston, 2005) was used to warp the chimeric images into the MNI space, and the transformed parameters were applied to the actual T1-weighted images, the lesion masks, and the functional images. The normalization results were carefully visually inspected before the next step to avoid possible errors.
First-level analysis was performed for each subject. The general linear model was set up as a boxcar function with “task” versus “control” conditions. Activation maps were generated at the threshold of p < 0.001 with cluster size ≥10 voxels. It has been observed that BA and WA can be reorganized asynchronously (Deng et al., 2015; Vikingstad et al., 2000); therefore, language lateralization was studied for the BA and the WA separately for each subject. The ROI of BA was defined as the pars opercularis and pars triangularis of the inferior frontal gyrus, and the ROI of WA was defined as the supramarginal, angular, and posterior portion of the superior and middle temporal gyrus. The laterality index (LI) was calculated with the formula LI = (L – R)/(L + R), where L and R represent the number of voxels activated in the ROIs of the left and right hemispheres, respectively. According to the LI value, hemispheric dominance was determined as left-sided (LI ≥0.2), right-sided (LI ≤ −0.2), and bilateral (−0.2 < LI <0.2) dominance (Deng et al., 2015; Lee et al., 2010; Seghier, 2008). Left-sided dominance was classified as typical lateralization, while right-sided or bilateral dominance was classified as atypical lateralization.
LI is a single value indicating the relative activation strength of the left and right hemispheres. It was noted that the fMRI signal might be susceptible to flow abnormalities associated with AVMs, causing false-negative activation in the vicinity of the lesions (Ulmer et al., 2004). Consequently, it could not be excluded that an LI <0.2 was due to an artifactual decrease in language activation in the left hemisphere without a concomitant increase in the right hemisphere. To corroborate the determined language dominance, we performed a group-level analysis based on the number of activated voxels in the right hemisphere (Lee et al., 2010) and an individual-level corroborative analysis based on the natural feature of crossed cerebrocerebellar language lateralization (Mendez Orellana et al., 2015). The details and results of the corroborative analyses are presented in Supplementary Table S1.
In addition, the absolute activation strength of the right hemisphere was estimated following the steps below. First, the mean activation map was generated for the patients showing atypical lateralization. Next, two peak points of the maximal t value within the right homologous BA and WA were localized at (51, 18, 3) and (51, −30, 0), respectively. Frontal and temporoparietal spherical ROIs (sROIs) with a radius of 5 mm were created using the MarsBaR toolbox centered on the two peak points (Supplementary Fig. S1). The activation strength of the right homologous BA and WA was expressed by the average t value of voxels within the two corresponding sROIs. A similar approach has been described by Putnam et al. (2008). The sum of the activation strength of the two sROIs was deemed to be the overall right-hemispheric activation strength. As the activation of the right hemisphere was not affected by possible artifacts related to AVMs, we took it as an alternative indicator of interhemispheric language reorganization to supplement the main analyses.
DTI processing
We performed DTI analysis using FreeSurfer software. First, the CC was segmented into five equal subregions by the automated FreeSurfer pipeline following the common approach (Collinson et al., 2014; Fischl, 2012; Fischl et al., 2002), with each subregion representing a fifth of the total CC, that is, anterior, midanterior, central, midposterior, and posterior segments (Fig. 1). The reproducibility and reliability of automated FreeSurfer segmentation of the CC have been consistently reported (Guenette et al., 2018; Herron et al., 2012). This automated method can reduce random errors, rater bias, and interparticipant variability compared with manual segmentation. Notably, to overcome signal loss caused by the lesions, we used the generated enantiomorphic-filling anatomical images instead of the actual T1-weighted images for CC segmentation. A similar approach has been previously described (Solodkin et al., 2010). Careful visual inspection and additional manual editing were required to ensure accuracy of the segmentation results.
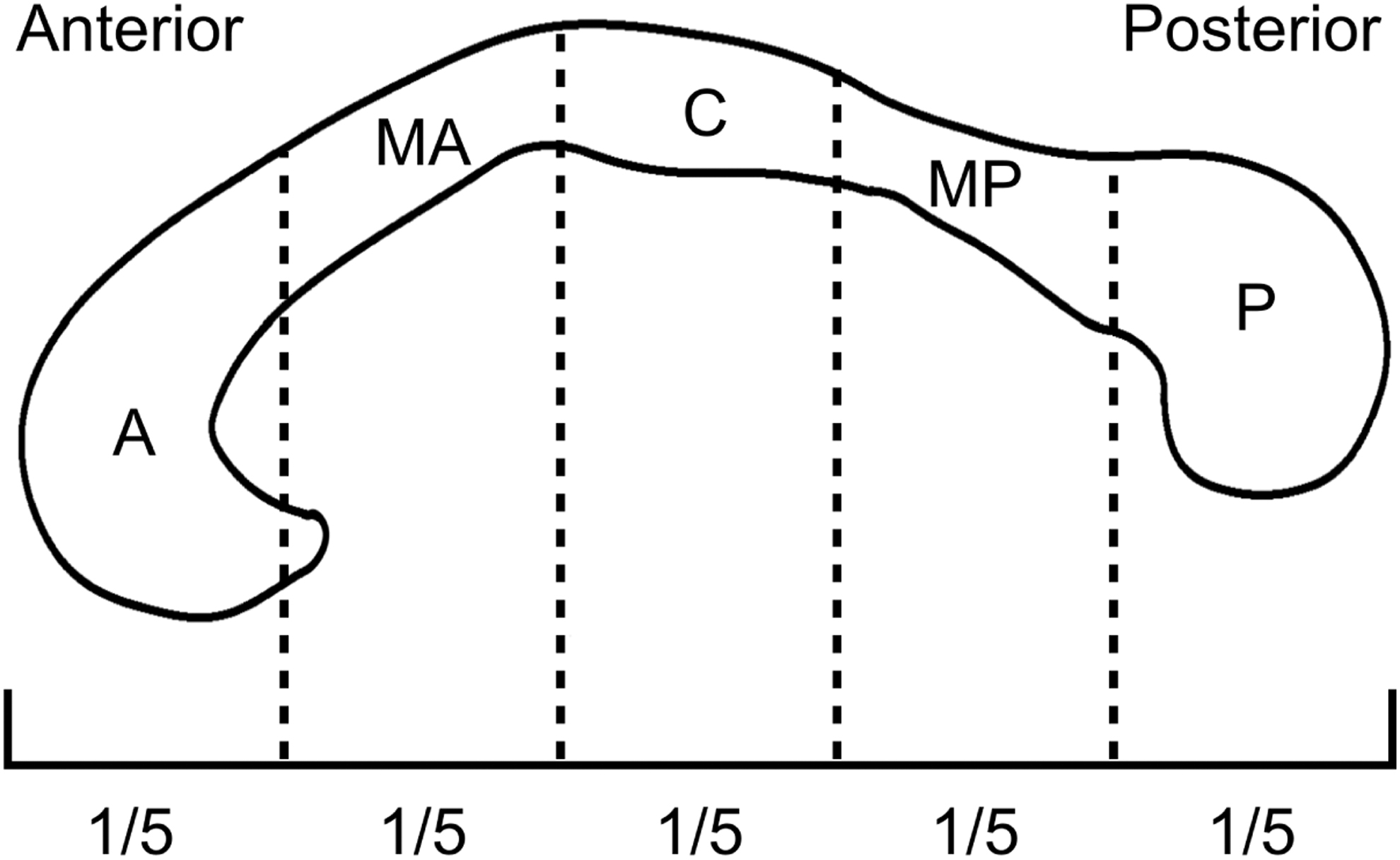
Schematic showing five equal subregions of the CC, segmented using FreeSurfer software. A, anterior; C, central; CC, corpus callosum; MA, midanterior; MP, midposterior; P, posterior.
The patient's head was immobilized using a head immobilizer during image acquisition to minimize possible interleave artifacts associated with head motion. A parallel imaging method (GRAPPA) was used to reduce susceptibility distortions. Eddy currents and residual head motion artifacts were corrected in preprocessing the DTI data. FA maps were acquired and coregistered to individual anatomical images using the diffusion pipeline of FreeSurfer. Then, we resampled the created CC segmentations from the anatomical data into the diffusion space. Finally, the mean FA values of the five subregions and the total CC were extracted.
Statistical analysis
We performed statistical analysis using SPSS 25.0 (SPSS Inc., Chicago, IL). Patients with atypical lateralization in the BA, WA, or both areas were classified as the atypically lateralized group, and the rest with typical lateralization in both areas were classified as the typically lateralized group. Chi-square tests and Mann–Whitney U tests were used to determine whether patient demographics and lesion characteristics affected language lateralization. Factorial analysis of variance (ANOVA) was performed to test for possible effects of language lateralization on the FA values of the total CC and the subregions. To adjust for multiple comparisons, the Bonferroni correction was applied to the significance level by dividing it by five given that five callosal segments were investigated, resulting in a corrected significance level of 0.01. Two-way ANOVA was performed to test for possible separate and combined effects of BA and WA lateralization (i.e., atypical or typical). Linear regression analysis was used to test for the correlation between the FA of the total CC and the right-hemispheric activation strength. All analyses were controlled for patient age, lesion location, and normalized lesion volume. A p value of <0.05 was considered statistically significant.
Results
Patient and lesion characteristics
The clinical data of the study population are presented in Table 1. Of the 41 patients with AVMs, 27 (65.9%) were male, 14 (34.1%) were female, and the mean age was 28.5 ± 10.7 years. Nineteen (46.3%) lesions were located in the BA, and 22 (53.7%) lesions were located in the WA. Only 1 patient with AVMs in the WA presented with intermittent aphasia; the remaining 40 patients did not exhibit language dysfunction before participation. The mean maximal diameter of the lesions was 3.7 ± 1.2 cm. Figure 2 shows the lesion overlay of all the patients on the MNI ICBM-152 template. The average normalized lesion volume was 21.3 ± 16.1 mL, and the average volume percentage of the language area involved was 16.4% ± 17.4%.
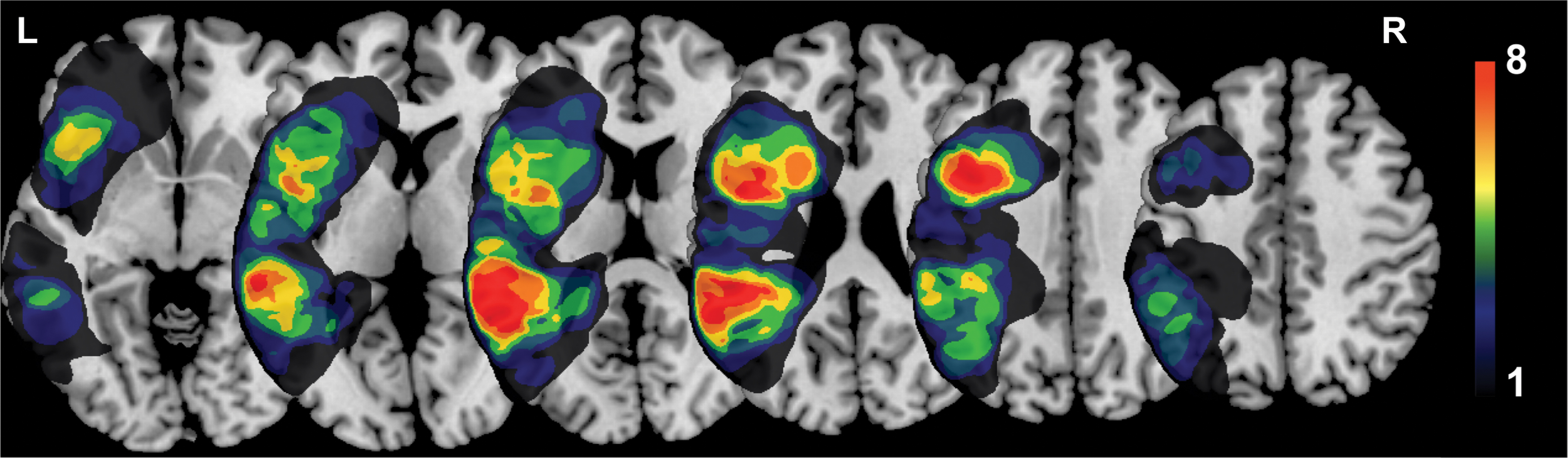
Lesion overlay of all patients (n = 41). Colors represent the number of patients with a lesion in the area. Warmer colors indicate greater overlap. Color images are available online.
Clinical Data of the Study Population
Categorical variables are presented as n (%), and continuous variables are presented as mean ± standard deviation.
BA, Broca's area; WA, Wernicke's area.
Language lateralization
All patients completed the verb-generation task. We observed atypical language lateralization in 16 (39.0%) patients, including 2 patients with atypical lateralization in the BA alone, 8 in the WA alone, and 6 in both areas, and the remaining 25 patients were classified as the typically lateralized group. Figure 3 shows example cases with different patterns of atypical language lateralization. The number of patients with left-, bilateral, and right-hemispheric dominance is detailed in Table 2. For the 19 lesions located in the BA, 4 patients exhibited atypical lateralization in the BA, and 5 patients showed atypical lateralization in the WA. For the 22 lesions located in the WA, 4 patients presented atypical lateralization in the BA, and 9 patients showed atypical lateralization in the WA. Statistical analysis revealed no significant differences in patient demographics or lesion characteristics between the typically and atypically lateralized groups (Table 3).
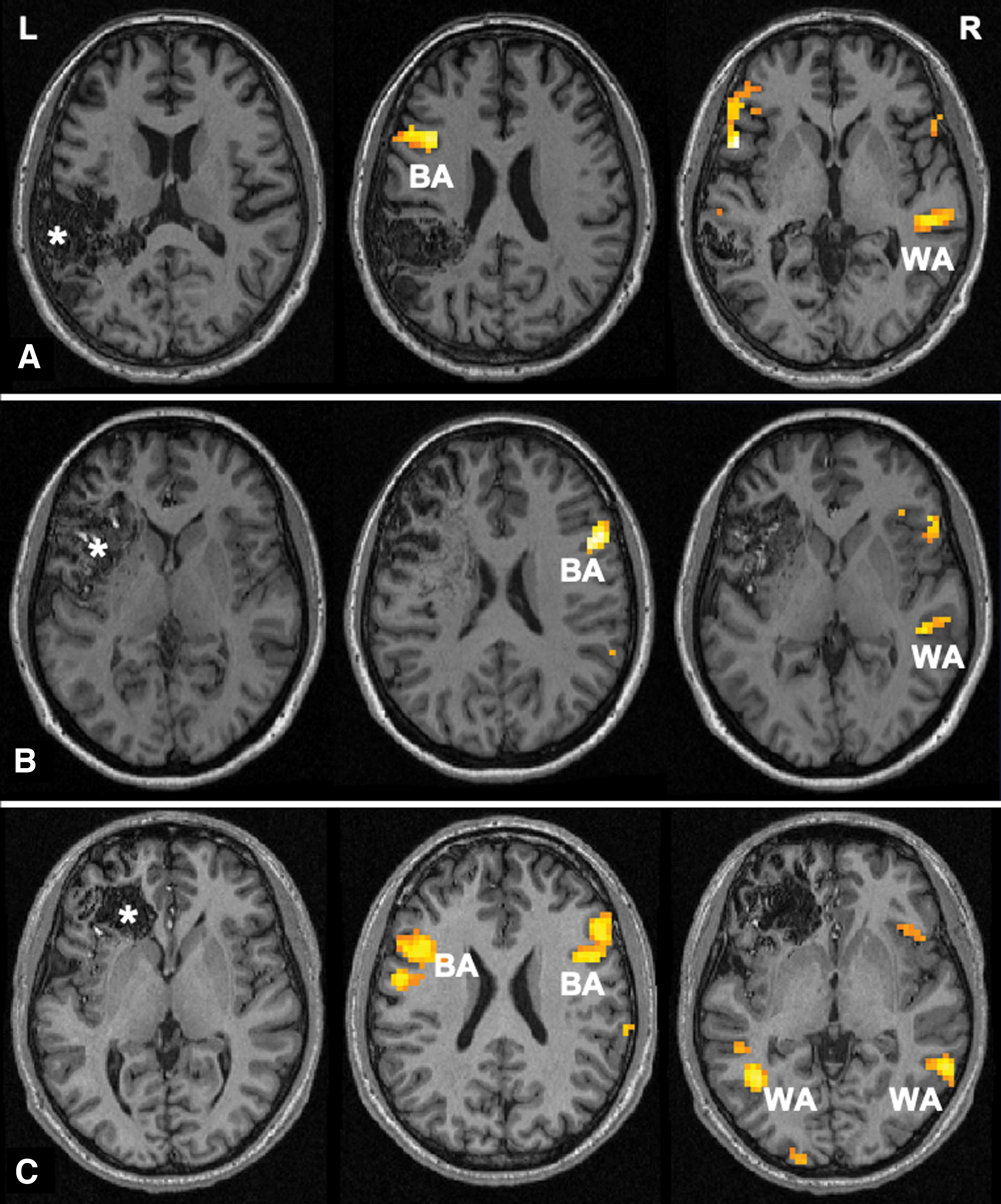
fMRI activation maps for the verb-generation task.
Number of Patients with Left-, Bilateral, and Right-Hemispheric Dominance in Broca's Area and Wernicke's Area
Relationship Between Clinical Characteristics and Language Lateralization
Categorical variables are presented as n (%), and continuous variables are presented as mean ± standard deviation. p Values were calculated using the Wilcoxon rank-sum test for continuous variables and the chi-square test for categorical variables.
Effects of atypical lateralization on callosal FA
We performed analyses for the difference in CC diffusion anisotropy between the typically and atypically lateralized groups, with patient age, lesion location, and normalized lesion volume as covariates. Figure 4 compares the mean FA values of the total CC and the segmented CC subregions between the two groups. Significant main effects of atypical lateralization were observed for the mean FA values of the total CC [F (1, 35) = 11.36, p = 0.002] and three subregions, including the anterior [F (1, 35) = 4.24, p = 0.047], midanterior [F (1, 35) = 13.82, p = 0.001], and midposterior subregions [F (1, 35) = 4.41, p = 0.043], of which the association of the midanterior subregion survived Bonferroni correction for multiple comparisons (p < 0.01). Patients in the atypically lateralized group exhibited significantly higher mean FA values in the total CC and the above subregions than those in the typically lateralized group. However, no statistical significance was found for the central subregion or the posterior subregion. There were no significant effects of patient age, lesion location, normalized lesion volume, or their interactions.

Mean FA values of the total CC and the five subregions in typically and atypically lateralized groups. After controlling for patient age, lesion location, and normalized lesion volume, the atypically lateralized group exhibited higher mean FA values in the total CC and the anterior, midanterior, and midposterior subregions. *p < 0.05, **survived Bonferroni correction for multiple comparisons. FA, fractional anisotropy.
Combined effects of BA and WA lateralization on callosal FA
To test for possible separate and combined effects of BA and WA lateralization (atypical vs. typical) on CC diffusion, we performed separate two-way ANOVAs on FA values of the total CC and its subregions. The main effect was not statistically significant for either BA or WA lateralization. However, we found significant interaction effects on the FA values of the total CC [F (1, 31) = 9.29, p = 0.005] and the midanterior subregion [F (1, 31) = 9.46, p = 0.004]. Further simple effects tests showed that crossed lateralization of BA and WA was associated with higher callosal FA compared with patients with colateralization of both areas (Fig. 5). Specifically, patients with atypical BA and typical WA exhibited higher FA of the total CC than those with both typical areas [F (1, 34) = 6.36, p = 0.017]; patients with typical BA and atypical WA exhibited higher FA of the total CC than those with both typical areas [F (1, 34) = 8.69, p = 0.006], and higher FA of the midanterior subregion than those with both typical areas [F (1, 34) = 13.92, p = 0.001] and those with both atypical areas [F (1, 34) = 4.53, p = 0.041]. The other comparisons were not significant.

Margin plots showing significant interaction effects of BA lateralization and WA lateralization on FA values of the total CC and the midanterior subregion.
Supplementary analysis of the correlation between right hemispheric activation strength and callosal FA
Compared with the typically lateralized group, the activation strength of the atypically lateralized group was significantly greater in the right frontal sROI (1.96 ± 0.78 vs. 0.45 ± 1.28, p < 0.001) and the right temporoparietal sROI (1.79 ± 1.10 vs. 0.24 ± 1.36, p < 0.001). Multiple linear regression showed that the FA of the total CC was positively correlated with the activation strength of the right frontal sROI (p = 0.041), the right temporoparietal sROI (p = 0.032), and the overall right-hemispheric activation strength (p = 0.018; Supplementary Fig. S2).
Discussion
In this series of patients with unruptured AVMs involving the anatomically defined language cortex, only 1 (2.4%) patient had a history of language dysfunction before participation. It is thought that because of the frequently occurring functional reorganization (Alkadhi et al., 2000; Vikingstad et al., 2000), an unruptured AVM usually does not lead to focal neurological dysfunction. fMRI has been extensively used as a noninvasive method for language mapping, with sufficient accuracy for predicting hemispheric dominance comparable with the Wada test (Dym et al., 2011). Previously, several studies focusing on the concordance of fMRI with electrocortical stimulation mapping (ESM) demonstrated the high sensitivity of fMRI for determining language localization in patients with brain lesions, including vascular malformations (Benson et al., 1999; Cannestra et al., 2004; Lee et al., 2010; Pouratian et al., 2002). Based on the verb-generation task, we found that the incidence of atypical language lateralization in the current series was 39.0% (16 of 41), with rates of 22.0% and 34.1% for BA and WA, respectively. This incidence was significantly higher than the reported incidence of 4% to 9% in healthy right-handers (Haberling et al., 2011; Isaacs et al., 2006; Knecht et al., 2000). We found that lesion size and the Spetzler–Martin grade were not associated with atypical lateralization, which was consistent with the results reported by Lee et al. (2010). Our results further proved the great capacity for language reorganization of the brain with congenital lesions, which has previously been revealed by a number of studies (Deng et al., 2015; Lehericy et al., 2002; Vikingstad et al., 2000).
It is worth noting that the development of language lateralization in the human brain is not well characterized. In one hypothesis, it is believed that language lateralization is primarily ontogenetically determined at an early developmental stage, suggesting that gene expression asymmetries assign language to one hemisphere (Bishop, 2013; Vingerhoets, 2019). Recently, some researchers have indicated that environmental factors may greatly affect the ontogenesis of language lateralization (Ocklenburg and Gunturkun, 2019). Forming from the fetal stage, an AVM may be regarded as an internal environmental factor given that it continues to affect the blood perfusion of the surrounding parenchyma with aging. In our view, most language reorganization in AVM patients may emerge because of progressive tissue damage during children's language learning, a time when typical lateralization of language is strengthened in healthy individuals. However, since AVMs are not completely static (Komiyama, 2016), it cannot be ruled out that this compensatory reorganization is triggered by AVM growth later in life in some cases. In another hypothesis, language is thought to be represented a priori in both hemispheres, and callosal development plays a key role in hemispheric lateralization through mediating interhemispheric communication (Hinkley et al., 2016). However, the relationship between callosal connectivity and language lateralization remains ambiguous, dividing views of callosal function into two models: the inhibitory model and the excitatory model (van der Knaap and van der Ham, 2011), as described in previous sections.
Our recent research demonstrated that AVMs in language areas can lead to wide-ranging changes in white matter organization (Li et al., 2020). The coupling of functional and structural connections has been found to be disrupted under pathological states (Zhang et al., 2011). Given the chronic disease course and high incidence of language reorganization, the current series provides a proper model to investigate the remodeling of callosal organization accompanying atypical language lateralization caused by focal brain damage. FA promises to be a proper approach for quantifying interhemispheric connectivity, since it reflects the underlying microstructure (e.g., axonal density, extent of myelination) directly related to the speed of interhemispheric transfer (Barbeau et al., 2015; Chepuri et al., 2002; Putnam et al., 2008). In this study, we found that higher callosal FA was associated with atypical language lateralization, in agreement with the excitatory model reported in healthy individuals, as well as brain tumor patients (Haberling et al., 2011; Tantillo et al., 2016). These findings indicate the excitatory function of the CC in the brain with focal lesions for maintaining the contralateral homologous regions in a functionally activated state. It can be inferred that acquired damage to the anatomical eloquent areas leads to cortical reorganization in the contralateral hemisphere to compensate for the damaged function, and correspondingly, greater information transfer between the two hemispheres is needed. This is somewhat in agreement with the idea proposed by Gazzaniga (2000) that interhemispheric information transfer is less needed in individuals with naturally strong language lateralization.
With respect to the conflicting hypotheses on the role of the CC, one possible explanation is that interhemispheric communication involves a dynamic and probably simultaneous interplay between inhibitory and excitatory effects (Bloom and Hynd, 2005; Haberling et al., 2011), which might vary according to the complexity of the task, as well as the health of the brain. This is based on the hypothesis that the interhemispheric conduction delay is partly the reason for functional lateralization (Aboitiz et al., 2003; Ringo et al., 1994). In healthy individuals, for less cognitively demanding tasks, the CC would play an inhibitory role in suppressing the nondominant hemisphere because information can be processed more efficiently by the single dominant hemisphere without interhemispheric delay. Conversely, for more cognitively demanding tasks, the CC may become excitatory for recruiting the nondominant hemisphere because a collaboration of both hemispheres is required for completing the complex tasks. We suppose that in patients with brain disorders, the lesioned hemisphere cannot afford enough information processing for intact language function, and therefore, the CC exerts excitatory effects to activate the contralateral hemisphere (van der Knaap and van der Ham, 2011; Welcome and Chiarello, 2008).
In addition, we sought to investigate the relationships between language lateralization and the FA of callosal subregions. We found that atypical lateralization was associated with significantly higher mean FA values in the anterior, midanterior, and midposterior subregions, but not in the central or posterior subregions. The five equal subregions broadly correspond to specific functional subdivisions. According to prior partitioning schemes of the CC (Hofer and Frahm, 2006; Witelson, 1989), the anterior subregion contains fibers mostly projecting to the prefrontal region; the midanterior subregion contains fibers mostly projecting to the premotor and supplementary motor cortex; and the midposterior subregion contains fibers projecting to the posterior parietal and superior temporal cortex. As expected, these cortical regions serve language functions. Our results suggest that the anterior, midanterior, and midposterior subregions of the CC may be responsible for mediating interhemispheric communication for language reorganization in these patients.
Consistent with previous studies, our results showed that the BA and WA can be asynchronously reorganized (Deng et al., 2015; Vikingstad et al., 2000), leading to crossed lateralization of the BA and WA (e.g., right-lateralized BA with left-lateralized WA). Compared with the direct interconnection between the BA and WA located in the same hemisphere, the connections between the crossed BA and WA remain unclear. A recent study demonstrated a remodeled functional connection in a glioma patient, where the right-lateralized BA was indirectly connected to the left-lateralized WA, relayed by the premotor and supplementary motor area (Li et al., 2019). We suppose that the callosal microstructure might be reorganized to serve the connection between the crossed BA and WA. We investigated the interaction effects of BA and WA lateralization on the callosal FA and found that patients with crossed lateralization of the BA and WA were associated with a higher FA value in the midanterior subregion compared with those with colateralization of both areas. Given that the midanterior CC mainly contains fibers projecting to the premotor and supplementary motor cortex, our results suggest that the midanterior subregion may play a role in the interhemispheric communication of the contralaterally located BA and WA.
From the perspective of structure-function relationships, our current results demonstrate changes in the largest white matter structure underlying interhemispheric language reorganization caused by brain AVMs, providing insights into the understanding of compensatory plasticity under pathological states. Clinically, our findings may highlight the importance of protecting transcallosal fibers during the treatment of AVMs in language areas. Given that high callosal FA may indicate a greater chance for language reorganization, this metric may be a promising marker for the recovery of language after hemorrhage or treatment. However, the interplay between functional and structural connections under pathological states remains intricate and needs to be investigated in depth.
The major limitation of the current study is that we lacked ESM or Wada test data to validate the fMRI results. As mentioned earlier, there is a possibility of pseudodominance of language activation as a consequence of neurovascular uncoupling associated with AVMs (Ulmer et al., 2004). For this issue, we performed corroborative analyses at the group and individual levels. The results suggested that the influence of neurovascular uncoupling seemed to be limited in these patients, in agreement with prior studies (Lee et al., 2010; Pouratian et al., 2002). Moreover, supplementary analysis revealed a significant correlation between callosal FA and the activation strength of the right hemisphere, which strengthened our main findings.
This study has other limitations. First, combined-task analysis (CTA) is thought to yield a more reliable determination of language lateralization than a single-task analysis (Dodoo-Schittko et al., 2012; Ramsey et al., 2001). However, we did not use a CTA approach in the current study due to a lack of reliability of the picture-naming task (Bradshaw et al., 2017; Jansen et al., 2006; Rutten et al., 2002). Second, the five subregions of the CC defined in this study might not exactly correspond to actual functional subdivisions, and the interplay between one subregion and a specific eloquent cortex should be interpreted with caution. Third, the sample size of this study was small, which might limit the ability to draw significant conclusions from multiple comparisons. Last, data from healthy controls collected by the same method and equipment were unable to be included in this study, hindering us from observing callosal changes attributed to the lesion itself. Future studies with larger sample sizes and functional results of the ESM or Wada test are required to verify our findings.
Conclusions
Herein, we demonstrated that atypical language lateralization was associated with higher callosal FA values, suggesting that interhemispheric connectivity was strengthened to accommodate the interhemispheric language reorganization associated with brain AVMs. Furthermore, the significant interaction effects of BA and WA lateralization on callosal FA suggested a greater demand for interhemispheric communication in individuals with crossed lateralization of the BA and WA. The current study contributes additional insights into the understanding of functional and structural plasticity of the human brain under pathological states.
Footnotes
Acknowledgments
We thank Prof. Ke Li, Prof. Yawei Zeng, and the technicians for their assistance in MRI acquisition.
Authors' Contributions
S.W. was in charge of supervising the whole study. M.L. and J.W. contributed to the conception or design of the work. M.L. was responsible for drafting and revising. R.G., S.Y., Y.Y., and M.L. contributed to data collection. M.L., P.J., and Y.C. were responsible for analysis and interpretation of data. All authors contributed to article revision, read, and approved the submitted version.
Author Disclosure Statement
The authors declare that they have no conflict of interest.
Funding Information
This work was supported by the Beijing Municipal Science and Technology Project (grant number: D161100003816006, S.W.) and the National Key Research and Development Program of China (grant number: 2016YFC1301800, S.W.).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.