
Abstract
Background:
The neural mechanisms of highly superior autobiographical memory (HSAM) are poorly understood. To shed light on the functional magnetic resonance imaging (fMRI)-informed neurobiology of this condition, in this study we characterize for the first time the neurofunctional architecture of a 20-year-old individual (B.B.) with HSAM and no concurrent neurological/psychiatric or other clinical conditions.
Materials and Methods:
Relying on t-test inferential models comparing a single observation with a control group, we processed B.B.'s resting-state fMRI signal and compared it with the neurofunctional architecture of 16 young adults with normal autobiographical memory. Specifically, we analyzed large-scale brain networks, region-to-region functional connectivity, and connectivity indices informed by graph theory.
Results:
B.B. showed higher expression of large-scale and region-to-region connectivity, larger segregation of the pallidum and enhanced centrality of the temporal pole, orbitofrontal cortex and cerebellar lobule IX.
Conclusion:
These findings indicate that HSAM is associated with increased expression of neural pathways that support memory encoding, retrieval, and elaboration, but also with reduced expression of patterns typically involved in information control and metacognition, the use of which would be minimized thanks to automatic and accurate memory processing.
Impact statement
The findings that highly superior autobiographical memory (HSAM) is supported by increased expression of large-scale brain networks, upregulated memory-centered region-to-region pathways, and downregulated expression of pathways that normally support memory operations that are underutilized in HSAM greatly contribute to clarifying HSAM-specific mechanisms. They also inform computational models of retrograde and anterograde memory processing, and impact on the application of artificial intelligence methods for the modeling of memory or for solving memory-related classification problems in clinical populations. They might also be of impact on research in forensic science when relying on the assessment of content and accuracy of autobiographical retrieval.
Introduction
Highly superior autobiographical memory (HSAM) is a rare condition whereby individuals remember easily and accurately almost every day of their life in response to dates. This superior memory for personal events (Parker et al., 2006) typically starts from age 11 (Ally et al., 2013; Mazzoni et al., 2019), when mental time travel seems to develop (Piolino et al., 2007) and children can link personal occurrences to a reliable autobiographical timeline (Habermas and de Silveira, 2008). Moreover, functional connectivity within the default-mode network (DMN), which in part overlaps with the so-called “autobiographical-memory network” (AMN), does not fully develop until age 9–12 (Thomason et al., 2008).
Differently from all other forms of exceptional memory, HSAM seems exclusively limited to personal events (Mazzoni et al., 2019; Patihis et al., 2013). Moreover, there is no evidence that these individuals use specific strategies during autobiographical encoding or retrieval (LePort et al., 2017; Mazzoni et al., 2019). They have, however, a special ability to calculate dates and days of the week mentally, in the absence of autism or savant behavior (Mazzoni et al., 2019). Some previously described cases (e.g., LePort et al., 2016; LePort et al., 2017; Parker et al., 2006) indicate presence of obsessive compulsive disorder (OCD) traits; for example, keeping diaries and compulsively rehearsing personal events.
Although a link with OCD has also been reported in group studies (LePort et al., 2016), there is one recently described HSAM case with no OCD or other clinical traits (Mazzoni et al., 2019), suggesting that, although OCD can co-occur with HSAM, it is not a prerequisite.
Attempts have been made to understand the underlying pivotal mechanisms in HSAM. LePort and colleagues (2017) ruled out a role of higher cognitive skills, since only modestly better performance is observed in HSAM individuals, and only in few tests. In another study, this team focused on retention of autobiographical memories, and concluded that HSAM most likely relies on greater long-term consolidation (LePort et al., 2016).
Whether HSAM might be due to specific and idiosyncratic neuroanatomical features has also been investigated, as there is strong evidence of changes in brain structure being associated with changes in behavior, including memory (e.g., Boyke et al., 2008; Draganski et al., 2006). Ally et al. (2013) described a 20-year-old born-blind man with HSAM, finding reduction in his cortical and subcortical volumes, with the exception of the right amygdala, which was larger than in controls. The latter finding led the authors to highlight the amygdala as a pivotal structure in HSAM. In addition, increased connectivity was observed in postcentral and thalamic networks, and between the right amygdala and the right hippocampus. The posterior cingulate/ventral-precuneus network (which includes major DMN areas) showed instead decreased connectivity. While these findings are important, blindness from birth can also have induced crucial neuroarchitectural alterations. The risk of interpretative misconstrual, therefore, should not be neglected.
Regional anatomy was also analyzed in a study comparing a group of 11 individuals with HSAM with an age- and sex-matched control group through three types of macro/microstructural analyses: voxel-based and tensor-based morphometry for the modeling of tissue density and shape, respectively, and tract-based spatial statistics for the modeling of white-matter diffusivity (LePort et al., 2012). Higher gray-matter density was found in the HSAM group in the vicinity of the right anterior-ventral putamen, caudate, and posterior pallidum, while lower gray-matter density was instead found bilaterally in the anterior portion of the middle temporal gyrus and in the posterior intraparietal sulcus. White-matter hypodensity was then found bilaterally in several small regions proximal to anterior putamen, temporal pole, mid-temporal gyrus, right posterior pallidum, and right lingual gyrus. Shape differences were found in the left posterior insula, in the left anterior internal capsule, in the right lingual gyrus, and in the temporal lobe bilaterally. Finally, higher fractional anisotropy was observed bilaterally in tracts supplying parahippocampal gyrus and intraparietal sulcus, in the right lingual gyrus, and in posterior callosal fibers. These results were interpreted as linked to the superior abilities seen in HSAM, as several regions in which structural differences were found are part of the AMN (e.g., Maguire
Meta-analytical evidence indicates that OCD is associated with neurostructural abnormalities (i.e., reduced volume in left anterior cingulate and orbitofrontal cortex, bilaterally), which suggests alterations in thalamocortical pathways (Rotge et al., 2009). As a result, the interpretation of the structural and functional-connectivity results obtained so far remains incomplete.
Functional-activation data have been recently reported on one individual (“B.B.”) unaffected by any detectable psychopathology (Mazzoni et al., 2019). When provided with a date, B.B. is able to recognize it as “autobiographically relevant” in ∼2 sec (while typical times in controls are approximately five times longer), showing predominant activation of the precuneus and visual areas, and of most of the areas that are part of the AMN. This activation occurs during simple access to personal memories, before the development of a complete and detailed recollection, which suggests optimized transfer of information across the AMN. While the predominance of visual processing is in line with the role of visual imagery in autobiographical memory, optimization of region-to-region processing (possibly, in a cost-effective way) might be a rather unique and pure feature of HSAM, when no other psychopathologies interfere.
To address this hypothesis, we investigated the resting-state neurofunctional architecture of B.B. and of a group of matched controls through three independent and largely complementary analytical procedures: the study of selected large-scale brain networks known to sustain autobiographical memory and the exploratory modeling of the entire connectome through region-of-interest (ROI) analyses and graph theory.
Materials and Methods
Participants
B.B. was selected out of a pool of 156 adults who self-reported as having HSAM abilities. At time of testing, he was a 20-year-old British right-handed male adult with 15 years of educational attainment, normal health, and normal sociocognitive functioning. He also had autobiographical-memory screening and was the only one out of 15 individuals who passed prescreening (during which memories for 500 dates were collected) and completed successfully two semistructured phone interviews during which participants were asked to describe in detail any memory associated with each of 20 dates randomly selected by the experimenter out of the initial pool of dates.
B.B. had accurate recollections in 88.4% of cases, while the other participants were accurate only in 0.03% of cases (p < 0.001). His recollections were also significantly faster (1.8 vs. 11.52 sec, p < 0.001). B.B. also recalled more events related to his birthdays than control participants (9 vs. 2.06, p < 0.001). After the initial interviews, additional testing for consistency of B.B.'s recollections was also done by phone at time intervals ranging from 1 to 3 months using 300 of the 500 initial dates. This additional testing confirmed B.B.'s exceptional autobiographical recall abilities.
His performance on a battery of neuropsychological tests indicated cognitive abilities within normality, with an intelligence quotient (I.Q.) in the top 90th percentile (De Bartolo et al., 2016). An overview of his neurocognitive profile is included in Table 1. For the purpose of this investigation, a group of 16 healthy young male adults was included in the study comparisons. This group included professional and technical workers (i.e., trainees and PhD students, a neuropsychologist, a physiotherapist, two physicians, a test engineer, and a radiographer), as this occupational category is typically characterized by I.Q. levels comparable with B.B.'s (Reynolds et al., 1987). Age (mean 28.7 years, standard deviation [SD] 4.7 years) and educational attainment (mean 16.6, SD 3.2 years) did not differ between B.B and controls (t = −1.939, p = 0.072; t = −0.456, p = 0.655, respectively).
Narrative and Mathematical Explanation of Graph Descriptors
Performance Shown by B.B. on a Number of Neuropsychological Tests
B.B.'s performance on these neuropsychological tests was compared with the data acquired in a sample of 26 male participants aged 18–25 years. No p-value survived correction for multiple comparisons.
No neuroanatomical differences existed between B.B. and the group of control participants. All participants provided their written informed consent before study inclusion. All procedures were carried out in compliance with the 1964 Declaration of Helsinki and its subsequent 2013 revision. This study was approved by the Institutional Review Board of the IRCCS Foundation Hospital San Camillo (Venice, Italy) (Protocol No. 11/09 version 2).
Functional magnetic resonance imaging acquisition
The resting-state functional magnetic resonance imaging (fMRI) sequence was acquired immediately before the task-based experimental procedure that investigated B.B.'s memory activation in an autobiographical-memory retrieval and elaboration paradigm (Mazzoni et al., 2019). Two 200-volume runs were acquired at rest with the following specifications, designed to maximize full coverage from the brain vertex to the cerebellum: Philips Achieva 1.5 T scanner, TR 2 sec, TE 50 msec, voxel size 3.28 × 3.28 mm, slice thickness 6 mm, 20 contiguous axial slices acquired bottom-up per volume, 64 × 64 matrix size, 230 mm field of view, flip angle 90°. At least 200 volumes were acquired on the same scanner and with the same acquisition parameters for each of the 16 controls. Twenty seconds of dummy volumes were set before each acquisition to enable the scanner to reach equilibrium.
An anatomical Turbo Field Echo T1-weighted image was also acquired as part of each MRI protocol (1.1 × 1.1 × 0.6 mm3 voxel dimension, 0.6 mm gap, 256 × 256 × 280 matrix size, 250 mm field of view, 7.4 msec repetition time, 3.4 msec echo delay time, and 8° flip angle). The processing of anatomical scans served to confirm absence of neurostructural differences between B.B. and controls before the analysis of fMRI signal.
fMRI preprocessing
Processing of MRI images was run with MATLAB (version R2014a, Mathworks Inc., United Kingdom), statistical parametric mapping (SPM) 12 (Wellcome Centre for Human Neuroimaging, London, United Kingdom), and a number of toolboxes. A pre-processing procedure was launched to prepare the images for modeling. Scans were initially slice timed to homogenize each volume temporally. This was followed by spatial realignment in which volumes were superimposed in space, and volume-to-volume in-scanner motion was estimated. The graphic representation of the six rigid linear and rotational motion parameters was visually inspected to identify the possible presence of any major problems. None of the movements made by B.B. or by any of the other participants was >0.5 voxels in any direction.
Following the procedures described by Power and colleagues (2014), indices of absolute and framewise displacement were calculated. No differences between B.B. and the control group were detected for any of these indices (all p-values >0.7). Images were then coregistered to their respective T1-weighted image, normalized and registered to the standard Montreal Neurological Institute echoplanar template, and a band-pass filter (0.01–0.1 Hz) was applied. Finally, scans were smoothed with a 6 mm full-width at half-maximum Gaussian kernel.
Data processing
B.B.'s functional architecture was investigated following three parallel and complementary methodologies, each aimed at a specific set of functional characteristics: large-scale brain networks, ROIs functional connectivity, and nodal features estimated through graph theory. By doing so, we emphasized theory-based neural pathways known to be at the basis of autobiographical memory abilities, as well as the exploration of the entire connectome through data-driven approaches.
Methodology 1-large-scale brain networks
An independent component analysis was run to identify the DMN and separate it from other sources of variability (Calhoun et al., 2001). The assumption behind the application of this statistical technique to fMRI is that the signal time course in each voxel (observed variables) is a linear mixture of multiple independent sources of variability (latent variables). Independent component analysis estimates an “unmixing matrix” that quantifies these latent source signals, minimizing mutual information or maximizing non-Gaussianity (Rosazza et al., 2012). The outcome is a series of independent components that are associated with a certain topography and are interpreted as “brain networks.” These are characterized by high levels of consistency from subject to subject (Damoiseaux et al., 2006).
The GIFT toolbox, running in SPM 12, was used for this purpose. The Infomax data reduction principle was chosen to estimate 20 independent components from the 17 datasets. The output was inspected to detect the map having the spatial features of the DMN (Fig. 1a). Beta-value maps were then extracted for inferential analyses.
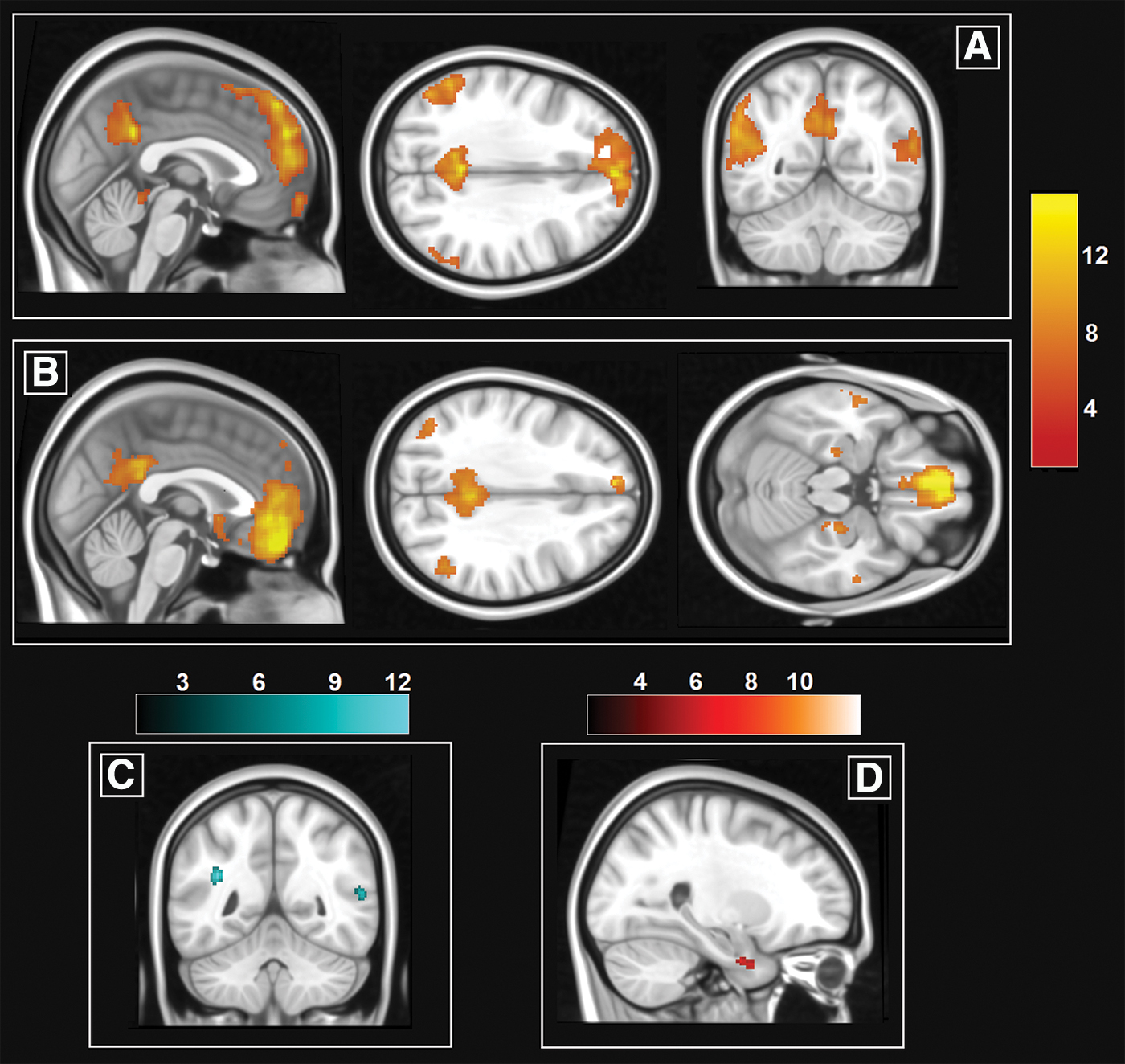
The two large-scale hemodynamic patterns investigated in this study.
The AMN was instead computed with a seed-based approach. Although the topographies of the DMN and of the AMN overlap to a considerable extent, there are a number of regions that show higher blood oxygenation level dependant (BOLD) signal during autobiographical retrieval than during rest (Ino et al., 2011). One of these, the ventromedial prefrontal cortex, was selected as seed.
SPM 12 and the CONN (version 18b) toolbox (Whitfeld-Gabrieli and Nieto-Castanon, 2012) were used to extract the BOLD signal time course from the seed and from the global maps of white matter and cerebrospinal fluid. As part of denoizing procedures, all images were scrubbed before signal extraction, and a series of variables were regressed out to minimize the impact of non-neural physiological variability (i.e., due to cardiorespiratory rhythms) and in-scanner motion. These included the first 10 principal components calculated within the tissue-specific map of white matter (5 components) and cerebrospinal fluid (5 components), following the aCompCor procedure (Muschelli et al., 2014). In addition, the global signal from the segmented maps of white matter and cerebrospinal fluid was regressed out.
A one-sample t-test was run to visualize the group-level map of functional connectivity of the ventromedial prefrontal seed. This pattern closely resembled the DMN, but included additional regions in the mediotemporal complex and in the anterior temporal lobe (Fig. 1b).
Methodology 2-regions-of-interest functional connectivity
A total of 132 ROIs derived from the Harvard-Oxford Atlas maximum likelihood cortical atlas and the Automatic Atlas Labeling were investigated to incorporate cortical (91 ROIs) and subcortical (15 ROIs) regions plus a detailed parcellation of the cerebellum (26 ROIs). Following denoizing procedures as illustrated above, the statistical association between each pair of ROIs was calculated with correlational models, and the correlation coefficients were converted into z scores using Fisher's r-to-z transformation. ROI-to-ROI functional connectivity calculated from B.B.'s dataset and from the group of controls is shown in Figure 2.
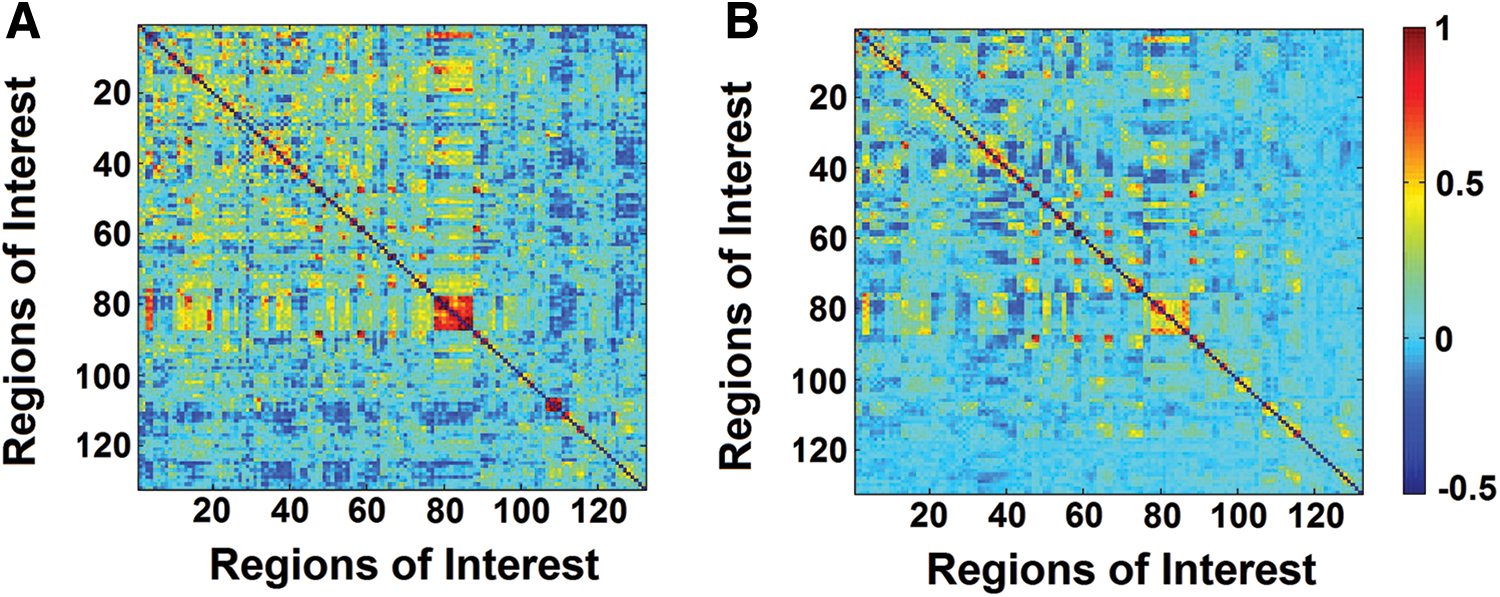
Correlation matrices illustrating the statistical strength of region-to-region functional connectivity calculated from resting-state fMRI time courses. B.B.'s matrix is shown on the left
Methodology 3-graph-theory functional architecture
To investigate in more detail fine-grained properties of functional architecture, a methodology informed by graph theory was adopted. Graph theory is a branch of mathematics that addresses systems made of interconnected elements. These elements, labeled as “nodes,” are connected by “edges” (Fig. 3a, b). When graph theory is applied to fMRI data, atlas-derived ROIs represent the nodes, and the strength of the statistical association between two time courses characterizes their edge (Wang et al., 2010). The bivariate correlations calculated among the 132 atlas ROIs were used to shortlist the edges having the strongest correlation coefficients. Multiple “absolute” and “proportional” edge-defining rules were set up using the “correlation” and “cost” options, respectively.

Fictitious example of graph-theory network
An index of global cost efficiency (Box 1) was calculated to quantify the performance of each rule and select the top 50 best performing absolute and proportional rules. As a result, individual binary graphs were constructed retaining edges: (1) associated with a correlation coefficient between 0.234 and 0.258 (one for each of the 25 “+0.001” increases); (2) retaining the top 17.1% to 19.5% (one for each of the 25 “+0.1%” increases) strongest associations within each individual graph (Fig. 3c, d). Seven indices were thus calculated for each node to quantify network properties based on the retained adjacency matrices. These (described in more detail in Box 1) were the cost and degree of each node (two basic descriptors), average path length and global efficiency (two indices of nodal integration), local efficiency and clustering coefficient (two indices of segregation) and, as an index of centrality, betweenness centrality (Rubinov and Sporns, 2010).
Inferential analyses
Linear group-comparison models were devised to test for the presence of differences between B.B.'s functional architecture and that computed in the group of healthy controls across the three methodologies. This was done using t-test models with pooled variances, to enable inferences between a single observation and a group (Crawford and Howell, 1998). Subject-specific maps associated with the DMN and with the AMN (Methodology 1) were modeled using SPM and a between-sample design. A conservative cluster-forming threshold of p < 0.0001 (uncorrected) was used for each analysis, and only clusters surviving a p < 0.05 (false discovery rate-corrected) were retained as significant. Clusterwise inference was preferred over voxelwise inference as it is more indicated for modestly powered studies (Woo et al., 2014).
The analysis of ROI-to-ROI patterns of connectivity (Methodology 2) was carried out with MATLAB and was thresholded at a Bonferroni-corrected p < 5.78−6 (i.e., controlling for 8646 independent comparisons).
Finally, for graph-theory analyses, differences in nodal descriptors across the entire graph were thresholded at a false discovery rate-corrected p < 0.05. In addition, findings were only considered significant when associated with an average t-score > 5 and when replicated in association with all 50 edge-defining rules.
Results
Methodology 1-large-scale brain networks
B.B.'s DMN was more expressed in the lateral temporal cortex and in the left inferior parietal lobule, bilaterally (Fig. 1c and Table 2), while his AMN was more expressed in the parahippocampal gyrus, uncus, and anterior temporal lobe (Fig. 1d and Table 2).
Differences in Large-Scale Brain Networks Between B.B. and the Group of Controls
BA, Brodmann area; FDR, false discovery rate; L, left; R, right.
Methodology 2-ROI-to-ROI functional connectivity
Of the entire set of t-tests, differences in three functional pathways survived the corrected threshold of significance (Fig. 4a, b). In all cases, B.B. showed higher functional connectivity compared with the group of controls. The pathways, illustrated in Figure 4c, included the connectivity between the left lingual gyrus and the right Heschl's gyrus, between the right orbitofrontal cortex and the right cerebellar lobule IX and between the right Heschl's gyrus and the contralateral planum temporale.
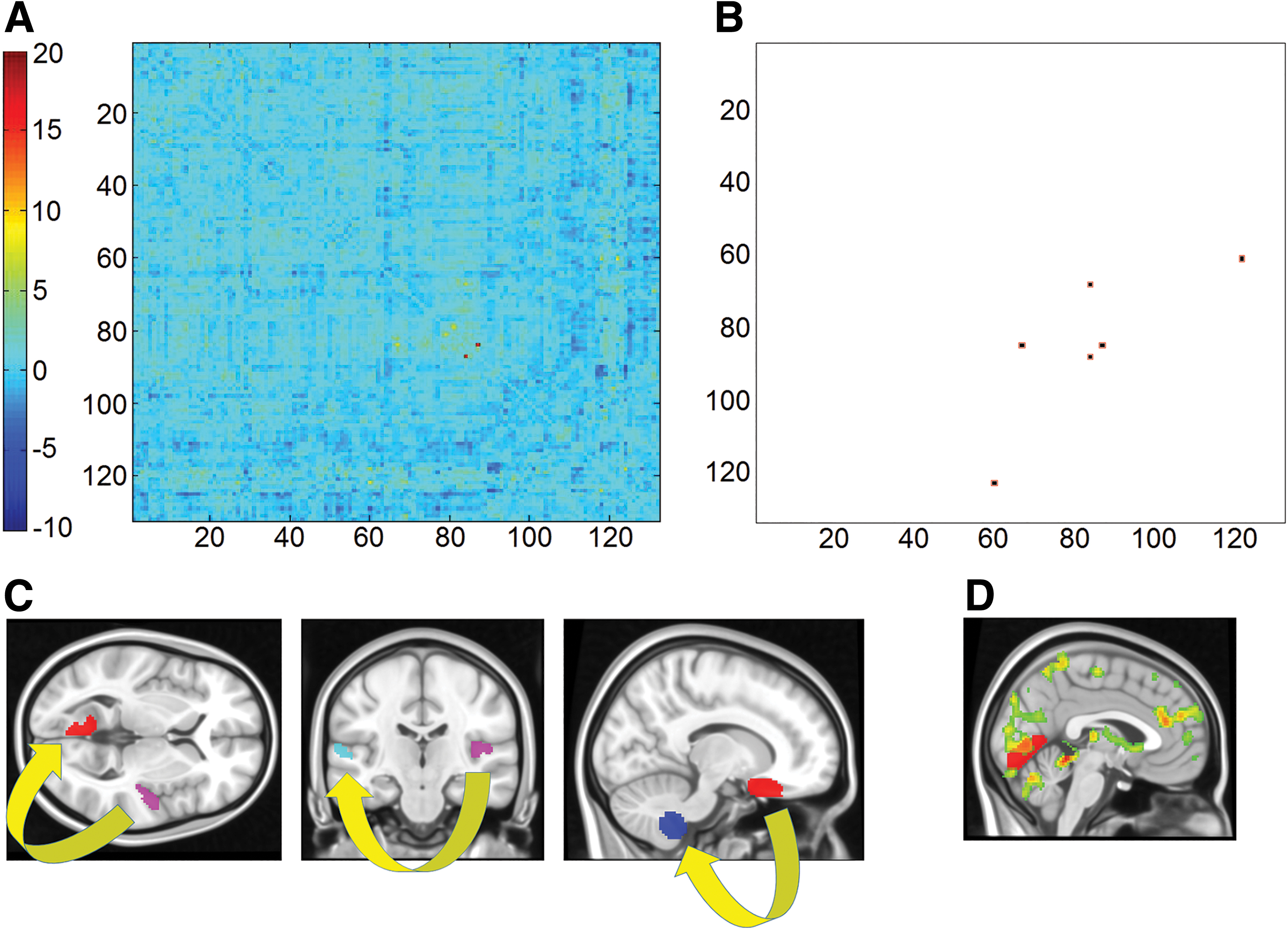
Differences in region-to-region pathways of functional connectivity between B.B. and the group of controls. Of the t statistics calculated over the entire set of region-to-region pathways
Methodology 3-graph theory
All results are included in Figure 5 and Table 3. No differences were found in cost, degree, average path length, or global efficiency for any of the nodes. B.B. had reduced local efficiency and reduced clustering coefficient in the left globus pallidus. B.B. also had higher betweenness centrality in the right temporal pole, in the right orbitofrontal cortex and in lobule IX within the right cerebellar hemisphere.
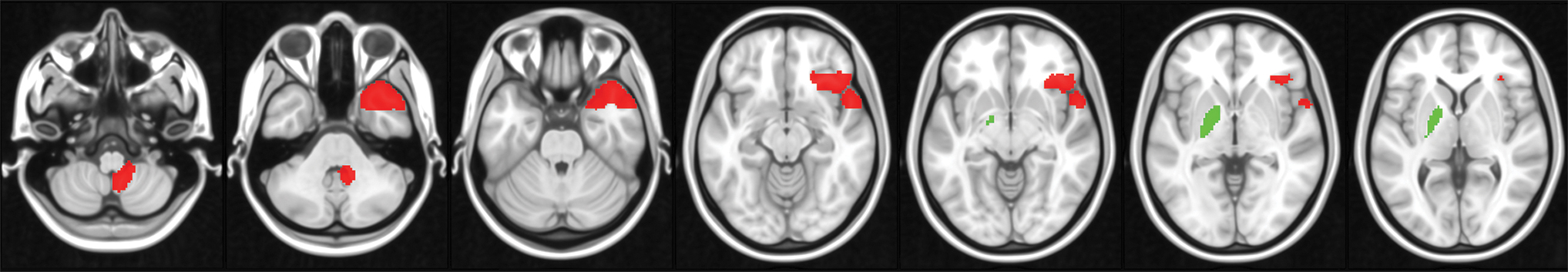
Differences between B.B. and controls for each of the three graph-theory descriptors that yielded significant findings. Green: controls > B.B.; red: B.B. > controls. Z slices in the Montreal Neurological Institute space are as follows: −54, −38, −12, −8, −2, and 2. Color images are available online.
Differences in Nodal Descriptors Between B.B. and the Group of Controls
Means and standard deviations of nodal t-scores were calculated across all graphs resulting from the application of the 25 edge-forming rules. Only findings emerged from all graphs and with an average t-score >5 are reported.
Discussion
The resting-state functional architecture of a young adult with HSAM was investigated, and compared with a group of young adults with normal memory and normal cognitive functioning. Multiple features were calculated from the BOLD signal of all participants to describe brain function according to multiple properties.
The analysis of large-scale brain networks showed that B.B. had increased connectivity within the DMN and within the AMN, in line with his abilities.
When specific region-to-region functional pathways were explored, B.B. showed a pattern of significantly increased connectivity in three of the tested models. One of these was located at the center of a pathway responsible for visual processing (involving the left lingual gyrus). Interestingly, this was aligned with the nature of the processing on which B.B.'s autobiographical memory relies, as described by B.B. himself (Mazzoni et al., 2019). In fact, as shown in Figure 4d, the activation associated with the elaboration of the retrieved memory reported by Mazzoni and collaborators (2019) extended to this region as well.
Two pathways in which B.B. had more functional connectivity than the group of controls were found along the patterns linking regions located in proximity of the Sylvian fissure (Fig. 4c). These regions normally support a range of abilities associated with auditory and linguistic processing (Dorsaint-Pierre et al., 2006; Griffiths and Warren, 2002).
The idea that B.B's brain connectivity is more expressed in regions deputed to this type of processing (with one of these patterns extending to the lingual gyrus) is consistent with the processing of inputs that prompt a visual elaboration of autobiographical memories (Gardini et al., 2006). A visual pathway is triggered once a date is mentioned and elaborated through a pathway that connects auditory processing to language comprehension. Arguably, the continuous engagement in such types of skills through this computational route results in the automatization of auditory and linguistic processing of dates and in the corresponding automatic triggering of visual elements associated with previous autobiographical memories. From this point of view, the functional correlate emerging from the analyses of B.B.'s resting-state circuitry is that of a lifelong repeated reinforcement of these pathways.
The third and final pathway in which B.B. showed increased functional connectivity was that between the right orbitofrontal cortex and the right lobule IX of the cerebellum. These two regions were also characterized by higher betweenness centrality, indicating a foreground role within B.B.'s functional architecture.
The orbitofrontal cortex plays a central role in autobiographical memory, interacting with the hippocampus during the formation of long-term memories (Ramus et al., 2007). Moreover, this region is responsible for processing the temporal context of memory (Duarte et al., 2010).
The right cerebellar hemisphere is another key region supporting autobiographical-memory retrieval, as revealed by meta-analytical evidence (Svoboda et al., 2006). Lobule IX is the cerebellar component of the DMN (Habas et al., 2009) and, in its right portion, is involved in the process of elaboration of past events during autobiographical memory (Addis et al., 2007). Temporal connotation of autobiographical memories and event elaboration are two elements of excellence that characterize B.B.'s performance, and it is particularly significant to note that the results of two analytical methodological approaches concur in highlighting a potential role played by these structures.
Finally, when B.B.'s pathways of connectivity were analyzed with methods of graph theory, two additional regions other than those mentioned above emerged as yielding significant differences consistently with all edge-forming rules. The first of these, the right temporal pole showed higher betweenness centrality, indicating that in B.B. this region is a node significantly more at the center of the graph compared with the group of controls. The right anterior temporal lobe plays an important role in autobiographical retrieval, as it supports the process of ecphory (Fink et al., 1996); that is, the interaction between the retrieval cue (the administered date, in this case) and stored information (the autobiographical content). This is also confirmed by the descriptions of clinical cases with autobiographical-memory impairment resulting from damage in this area (Calabrese et al., 1996; Kroll et al., 1997). On these grounds, a centrality of the right anterior temporal cortex would account for the simplicity with which B.B. ecphorizes personal memories from cue dates.
The globus pallidus was a second region characterized by significant graph theory-informed differences. Although the dorsal and ventral portions of this nucleus are involved in distinct functions, the region defined by the atlas implemented in our methodology encompassed them both. B.B.'s globus pallidus was supported by an adjacency matrix less intraconnected (lower clustering coefficient) and with longer internodal paths (lower local efficiency) than that of the controls. Studies in nonhuman primates have shown that the globus pallidus plays a central role in controlled information-processing tasks such as establishing goals (Arimura et al., 2013) and suppressing irrelevant stimuli (Kim et al., 2017). This is also confirmed by the description of deficits experienced by patients who underwent surgical pallidotomy (Piron et al., 2016). Moreover, the ventral pallidum is functionally coupled with the dopaminergic circuit that links the ventral tegmental area to the hippocampus when the stimuli are characterized by a degree of novelty (Lisman and Grace, 2005). Certainly, the lack of cognitive control shown by B.B. during the processing of autobiographical memories is symptomatic of the fact that these memories are not characterized by any novelty but are based instead on automatic computations.
To put things in perspective, resting-state function of the left globus pallidus is significantly upregulated in individuals with post-traumatic stress disorder (Disner et al., 2018), who are known to suffer from distorted processing of autobiographical memory (Ehlers and Clark, 2000; Schönfeld and Ehlers, 2017). This set of converging pieces of evidence contributes to characterize the globus pallidus as a region involved in the controlled processing of information flow. We argue that the relative “computational isolation” of the globus pallidus shown by B.B. may reflect his lifelong automatization of autobiographical-memory retrieval.
It is particularly significant to note that mnemonists, on the contrary, have to rely on the volitional control of mental strategies to achieve superior results (Maguire et al., 2003).
It is unquestionable that autobiographical memory is a cognitive construct that is central to our identity as individuals. In fact, elderly adults who are referred to a neurological examination usually complain of memory lapses that affect their own autobiographical experience (Youn et al., 2009). In this context, B.B.'s autobiographical memory is automatic, accurate, and rich in situational details. The continuous, lifelong computational reliance on such automatic, precise, and efficient function inevitably results in a minimal need to implement satellite processes such as those of metamemory (Mazzoni and Kirsch, 2002) or the executive resources that put memory under scrutiny, for example, those theorized by the models that account for the presence of anosognosia in neurological patients (Agnew and Morris, 1998).
The evidence from this individual with HSAM is of particular relevance for the understanding of the underling neural substrates supporting autobiographical memory, notably because this is the first case study of a normally developing individual, with no psychiatric or psychological complications, showing a spontaneous and effortless capacity to retain and retrieve personal information to a high level of details. The study of differences and similarities in his brain connectivity from normally able individuals not only sheds significant light on the brain regions supporting autobiographical-memory retrieval, but also explains individual differences in circuitry connectivity that can be exploited in the context of brain damage.
As final considerations, first it is worth stressing the importance of the evidence obtained from the detailed functional connectivity study of B.B. since this is the first set of findings of this kind obtained in a healthy individual. All previous sets of evidence, although valuable to help clarify this exceptional ability more broadly, remain of limited application to the understanding of HSAM in relation to normal ability. Nevertheless, it is important to note that many of the neural pathways emerged from this investigation converge with the topography of the findings reported in the case descriptions of individuals with HSAM reviewed in the introduction. This indicates that the neural resources at the basis of HSAM could be, in part, shared by individuals with and without comorbidities.
We argue that, for the definition of a general, data-driven theoretical framework of HSAM, it will be important to factor in and recognize the role of memory abilities, together with the intervenient role played by psychopathology, sensory deficits, and potential developmental differences in maturation of brain processes and structures.
On this note, it is also important to consider that mechanisms emerging from a single-case description might be specific to the individual in question (as it could be, in part, the case for B.B.). Considering B.B.'s abilities, however, it is apparent that superior autobiographical-memory skills may not be a reflection of massive differences in brain functional organization. On the contrary, they seem to be supported by differences in function of pivotal structures within the whole cognitive architecture and more broadly within the AMN. It is not possible to ascertain what can trigger functional changes in these structures. It can only be speculated that, for B.B., the observed differences are the reflection of developmental changes fostered without conscious awareness by his preferred retrieval strategy.
It is also important to remark that the analyses of single observations often reveal significant differences on one of the dependent variables while no significance is reported on the other outcomes. Such dissociations are characterized by the inevitable intrinsic risk of committing both Type I and Type II errors (Crawford and Garthwaite, 2012) and, for this reason, we acknowledge that particular caution is exercised when interpreting these findings. We predict, therefore, that the future description of further individuals with HSAM will contribute to minimizing this risk by confirming genuine patterns of findings.
Conclusion
The neurofunctional architecture sustaining HSAM is characterized by large-scale, region-to-region, and graph-theory-informed differences from the functional circuitry supporting normal autobiographical memory. While a number of functional pathways are upregulated in HSAM, other pathways are downregulated. The latter supports cognitive operations that appear to be underutilized in HSAM. Although we cannot establish the extent to which these findings apply to HSAM in general or are instead proper of B.B., this study contributes to the literature on this rare cognitive skill.
Footnotes
Acknowledgments
The authors thank B.B. for his collaboration and his patience. This is a summary of independent research carried out at the NIHR Sheffield Biomedical Research Centre (Translational Neuroscience). The views expressed are those of the authors and not necessarily those of the NHS, the NIHR, or the Department of Health.
Authors' Contributions
M.D.M. contributed directly to data processing, analysis, and visualization and contributed to the writing of the article. R.M. contributed to the drafting of the article. G.M and A.V. conceived the project, devised, and conducted the experiments and contributed to the revision and finalizing of the article. All authors approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.