
Abstract
Introduction:
Lateralization in brain function has been associated with age and sex in previous work; however, there has been less focus on lateralization of functional networks during development.
Aim:
We aim to examine laterality in typical development; a clearer understanding of how and to what extent functional brain networks are lateralized in typical development may eventually prove to hold predictive information in psychopathology.
Material and Methods:
In this study, we examine the lateralization of resting-state networks assessed with a group-independent component analysis using resting-state functional magnetic resonance imaging from a large cohort consisting of 774 children, ages 6–10 years. This is an extension of our previous work on normal aging in adults, where we now assess whether there are similar patterns in children.
Results:
Unlike the results from our study of healthy aging in adults, which showed a decrease in laterality with increasing age, in this study we found both decreases and increases in lateralization in multiple networks with development. For example, auditory and sensorimotor regions had greater bilateral connectivity with development, whereas regions including the dorsolateral frontal cortex (Brodmann area left 9 and left 46) showed an increase in left lateralization with development.
Conclusion:
Our findings support a complex, nonlinear association between laterality and age in school-age children, a time when brain function and structure are developing rapidly. We also found brain networks in which laterality was significantly associated with sex, handedness, and intelligence quotient, but we did not find any significant association with behavioral scores.
Impact statement
Lateralization in brain function has been associated with age and sex in several previous studies; however, there has been less focus on lateralization of functional networks during development. A clearer understanding of how and to what extent functional brain networks are lateralized in typical development may eventually prove to hold predictive information in psychopathology. In this study, we examine the lateralization of resting-state networks assessed with a group-independent component analysis using resting-state functional magnetic resonance imaging from a large cohort consisting of 774 children, ages 6–10 years.
Introduction
Lateralization, brain hemispheric specialization for certain tasks, was first observed by Broca (1861) and Wernicke (1874), and was later confirmed and extended by the work of Sperry (1974), and since then there have been a number of studies investigating the differences between the two cerebral hemispheres. Lateralization is a part of normal brain organization, and abnormalities in laterality have been shown in individuals with mental illnesses, such as schizophrenia (Agcaoglu et al., 2018; Son et al., 2017). Therefore, understanding laterality in children, a time when rapid brain development is taking place, is needed to provide insight into this understudied period of development and also has the potential to identify biomarkers related to mental illnesses, many of which manifest in childhood or include a developmental component.
Lateralization has been shown to strongly associate with age and has some association with sex; Szaflarski and colleagues (2006) investigated language lateralization in adults and children using a verb generation task, and found that left lateralization increases from age 5 to 20 years, reaches a plateau from 20 to 25 years, and decreases between 25 and 70 years. Nielsen and colleagues (2013) found a small increase in lateralization with age but no sex effects. Zuo and colleagues (2010) observed homotopic connectivity of regions showing linear, quadratic, and cubic associations with age as well as some sex differences in homotopic functional connectivity. Kann and colleagues (2016) investigated the laterality of resting-state functional connectivity in the anterior insula, and found evidence of age, sex, and personality trait effects. These resting-state studies either used a seed-based approach or compared homotopic voxels to calculate laterality (Kann et al., 2016; Nielsen et al., 2013; Zuo et al., 2010).
We have previously (Agcaoglu et al., 2015) investigated the laterality of subject-specific spatial maps of the resting-state networks (RSNs) estimated through group-independent component analysis (gICA) in a sex-balanced dataset consisting of 603 participants ranging from 12 to 71 years of age, and reported a significant association of laterality with both age and sex. Agcaoglu and colleagues (2015) also showed a decrease in laterality with increasing age, supporting the theory that brain networks become more bilateral with increasing age in a possible attempt to compensate neural decline with aging (Cabeza et al., 2002; Reuter-Lorenz et al., 2000). Luo and colleagues (2016) used a similar approach in a longitudinal study of older adults and investigated the effect of cognitive training in lateralization, and found evidence that cognitive training prevented decrease in laterality in older adults. Gracia-Tabuenca and colleagues (2018) examined laterality in a resting-state functional magnetic resonance imaging (rs-fMRI) dataset containing 60 school-age children (age 6–10), and found a negative relationship between visuospatial test scores and asymmetry of functional connectivity in the temporo-occipital cortex, but did not report any significant age effects. These studies either do not cover developmental periods, which show more rapid brain changes, or used small sample sizes.
In this study, we analyze the laterality of RSNs estimated through a gICA in a large dataset consisting of 774 children from ages 6 to 10. Based on our previous findings on healthy adults where we observed more bilateral RSNs with aging in adults (Agcaoglu et al., 2015), we hypothesized a global increase in laterality in school-age children with development, as it is a period of rapid brain growth. Also, we investigated association between sex and laterality during this period before puberty, as well as association with intelligence quotient (IQ), handedness, and behavioral score. Everts and colleagues (2009) found a positive correlation between verbal-IQ and the laterality during a language task in their task fMRI study containing 20 healthy subjects aging 8–20 years. Based on previous evidence linking brain laterality with higher IQ and association with behavioral score, we further hypothesize that IQ, handedness, and behavioral score will affect the laterality across core brain networks. This study is one of the largest single site studies in laterality.
Materials and Methods
The study was approved and monitored by the Medical Ethics Committee at the Erasmus MC University Medical Center in Rotterdam, The Netherlands. All participants' legal guardians provided signed informed consent before study procedures. We previously investigated laterality of RSNs in a large dataset consisting of adults ranging in age from 12 to 71 (Agcaoglu et al., 2015). In this study, we apply a similar approach to a younger population (age from 6 to 10). We summarize the methods for clarity.
Participants
The participants were involved in the Generation R Study, which is a large pediatric population-based cohort in Rotterdam, The Netherlands (Jaddoe et al., 2012). A total of 1070 children ages 6 to 10 years participated in an imaging substudy between September 2009 and July 2013 (White et al., 2013). Subjects with severe motor or sensory disorders (deafness or blindness), neurological disorders, claustrophobia, contraindications to MRI, or having history of moderate to severe head injuries with loss of consciousness were excluded. After excluding subjects with excessive head motion, a total of 774 subjects (403 males and 371 females) between 6 and 10 years of age were retained in this study. Handedness information was available for 773 subjects (697 subjects were right handed, 72 subjects were left handed, and 4 [%] subjects were ambidextrous). IQ data were available for 737 subjects, and child behavior checklist (CBCL) scores were available for 708 subjects. Demographic information of the subjects is summarized in Table 1.
Demographic Information of the Participants, Age, Sex, Handedness, Intelligence Quotient, and Child Behavior Checklist Scores
CBCL, child behavior checklist; IQ, intelligence quotient; SD, standard deviation.
Behavioral and cognitive data
Behavioral problems in children were assessed through maternal report using the CBCL/1½-5 (Achenbach and Ruffle, 2000) as part of the age-6 assessment wave (Tiemeier et al., 2012). The CBCL is a 99-item inventory that uses a Likert response format (e.g., “Not True,” “Somewhat True,” “Very True”). Seven syndrome scales, five Diagnostic and Statistical Manual of Mental Disorders-oriented scales, and three broadband scales are commonly derived summary measures from the CBCL (Achenbach et al., 2000; Tick et al., 2007).
General intellectual functioning was assessed during the age-6 assessment wave using an abbreviated version of the Snijders-Oomen Niet-verbale Intelligentie Test–Revisie (SON-R 2½-7) (Tellegen et al., 2005; Tiemeier et al., 2012). The SON-R 2½-7 is a measure of nonverbal intelligence for children between 2.5 and 7 years of age, and was selected to minimize language-dependent confounds that may be present in a large, ethnically diverse sample such as the Generation R Study. IQ was estimated from the two SON-R performance subtests that were administered (Mosaics and Categories), which is highly correlated with estimates resulting from the complete version (Basten et al., 2014).
Data acquisition
MRI data were collected on a 3T General Electric Discovery MR750 scanner (General Electric, Milwaukee, WI) using a standard eight-channel, receive-only head coil. A three-plane localizer was run first and used to position all subsequent scans. Structural T1-weighted images were collected using a fast spoiled gradient-recalled echo sequence (TR = 10.3 ms, TE = 4.2 ms, TI = 350 ms, NEX = 1, flip angle = 16°, matrix = 256 × 256, field of view [FOV] = 230.4 mm, slice thickness = 0.9 mm). Echo planar imaging was used for the rs-fMRI session with the imaging parameters: TR = 2000 ms, TE = 30 ms, flip angle = 85°, matrix = 64 × 64, FOV = 230 × 230 mm, slice thickness = 4 mm, with no gap between slices.
Initially, a total of 250 volumes (duration: 8 min 20 sec) were collected. However, after it was determined that fewer volumes were required to obtain stable RSNs (White et al., 2014), the number of volumes was reduced to 160 (duration: 5 min 20 sec) (White et al., 2014). Children were instructed to stay awake and keep their eyes closed and to think of nothing in particular during the rs-fMRI scan. See White and colleagues (2013) for further details of the scanning protocol.
Preprocessing
The data were preprocessed using a combination of toolboxes AFNI (Analysis of Functional NeuroImages available online), SPM (Statistical Parametric Mapping available online), GIFT (Group ICA Of fMRI Toolbox available online), and custom scripts written in MATLAB. The rigid body motion correction was performed using the INRIAlign (Freire and Mangin, 2001) toolbox in SPM to correct for subject head motion followed by slice-timing correction to account for timing differences in slice acquisition. Next, the fMRI data were despiked using 3D despike algorithm in AFNI to mitigate the impact of outliers. The fMRI data were subsequently warped to a Montreal Neurological Institute (MNI) template, resampled to 3 mm3 isotropic voxels, and smoothed with a Gaussian kernel to 5 mm full width at half maximum. Each voxel time course was variance normalized before gICA as this has shown to better decompose subcortical (SC) in addition to cortical networks. To limit the impact of motion, we excluded subjects' data with a maximum translation of >5 mm and/or with signal-to-noise fluctuation ratio <200 from our analyses, resulting in a final dataset with 774 subjects. See Rashid and colleagues (2018) for more preprocessing details.
Group-independent component analysis
The preprocessed functional data were decomposed into 100 spatially independent components using spatial gICA, implemented in the GIFT software (Calhoun and Adali, 2012; Calhoun et al., 2002). Thirty-eight components of 100 were identified as RSNs based on criteria such as peak activated voxels location, showing less overlap with known vascular, susceptibility, ventricular, and edge regions corresponding to head motion. These 38 RSNs were also grouped based on their anatomical and functional properties, including SC, auditory (AUD), sensorimotor (SM), visual (VIS), cognitive control (CC), default-mode networks (DMNs) and cerebellum (CB). See Rashid and colleagues (2018) for more details of the gICA procedure.
Warping to symmetric template
Brain hemispheres generally appear as mirror images of each other; however, they are not exactly symmetric and exhibit important differences; therefore we need to account for the asymmetric regions before any anatomical or functional comparison. Swanson and colleagues (2011) and Stevens and colleagues (2005) chose to warp the data to a symmetric template in the spatial normalization step to ensure any identified asymmetries are not induced by the template itself. In this study, subjects' fMRI images were initially normalized to an MNI template followed by calculation of a study-specific template computed as the average of the first time point from each subject. Later, we symmetrized this template by flipping in the lateral axis and averaging with its unflipped version. Finally, we performed normalization to the symmetric template as an additional step. As the final warping from the subject-specific template to the symmetric template is applied to all subjects, this can alternatively be applied directly to the spatial components following the gICA (Agcaoglu et al., 2015). We prefer this method to facilitate direct comparison with Rashid and colleagues (2018).
One sample t-test on spatial maps
We calculated one sample t-tests of the subject-specific symmetrized spatial maps, and thresholded at three standard deviations from a Gaussian gamma fit to both positive and negative voxels. We also created masks that corresponded to these thresholded regions, called activation masks. Figure 1 shows the symmetrized spatial maps as t-statistics, and Supplementary Table S1 shows the corresponding MNI regions.
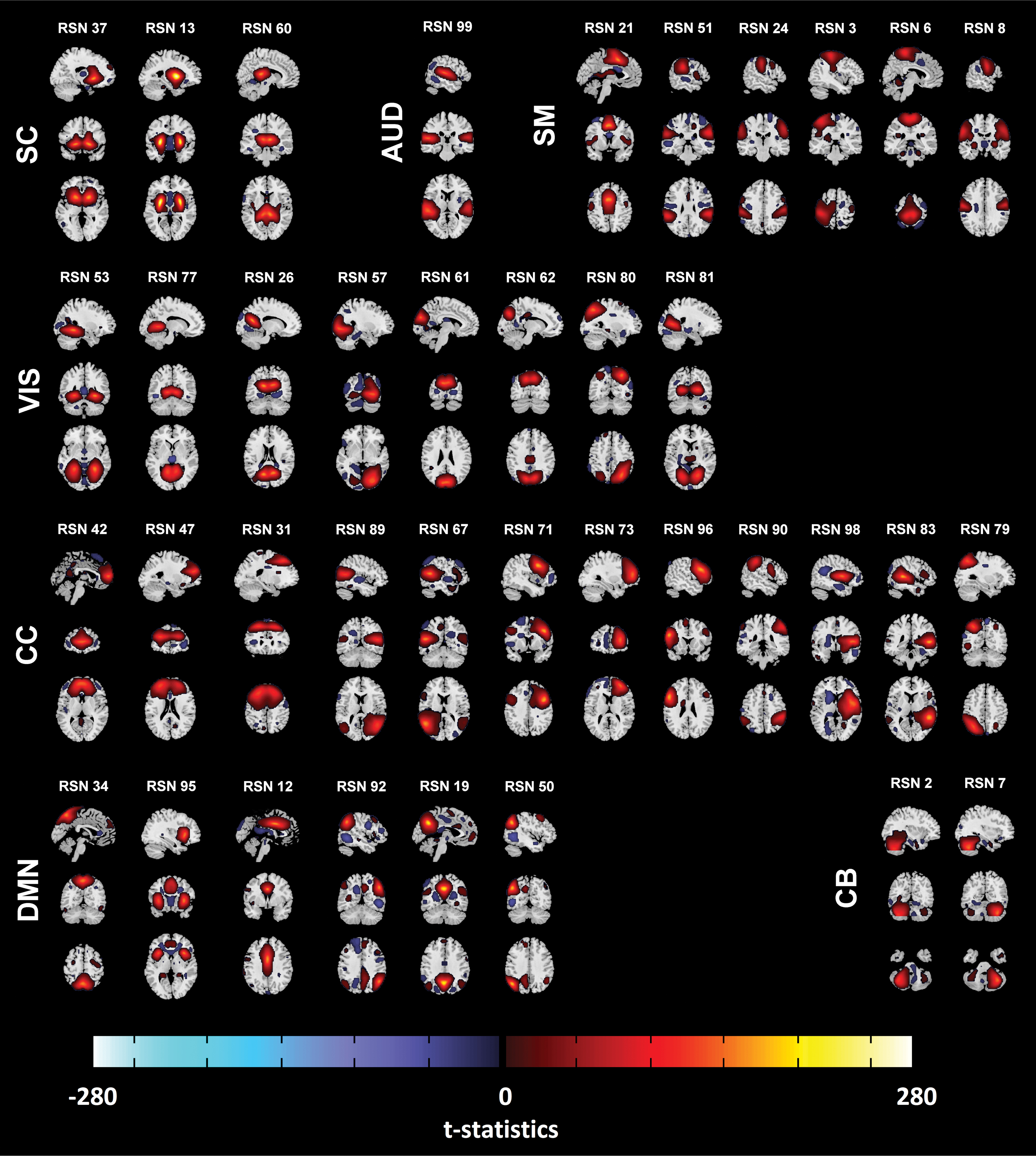
Thirty-eight resting-state components after symmetrizing, shown as t-statistics thresholded as three standard deviations using Gaussian gamma fit (both positives and negatives). The RSNs were grouped based on their anatomical and functional properties, including SC, AUD, SM, VIS, CC, DMN, and CB. See Rashid and colleagues (2018) for the details of grouping. AUD, auditory; CB, cerebellum; CC, cognitive control; DMN, default-mode networks; RSN, resting-state network; SC, subcortical; SM, sensorimotor; VIS, visual. Color images are available online.
Calculating laterality components
We calculated group-level and subject-specific laterality components, which show the regions that were lateralized and provided a visual summary about how the components were lateralized, and enable us to examine laterality in voxel level. For each component of each subject, we calculated the differences between each subject-specific component's voxel values on the right side of the cerebral cortex and its homotopic (geometrically corresponding) voxel on the left side. Later, we applied a one sample t-test for each component's difference map and thresholded t-statistics at three standard deviations using Gaussian gamma fit (only positive voxels). We used a positive cutoff since homotopic voxels have the same values with different signs on the laterality components. Finally, the activation masks were applied to include only the most functionally active parts of the components. This is called a group-level laterality component. An illustration of calculation of laterality components is displayed in Figure 2. The group-level laterality components are presented in Figure 3 as t-statistics, and corresponding MNI regions are presented in Supplementary Table S2.
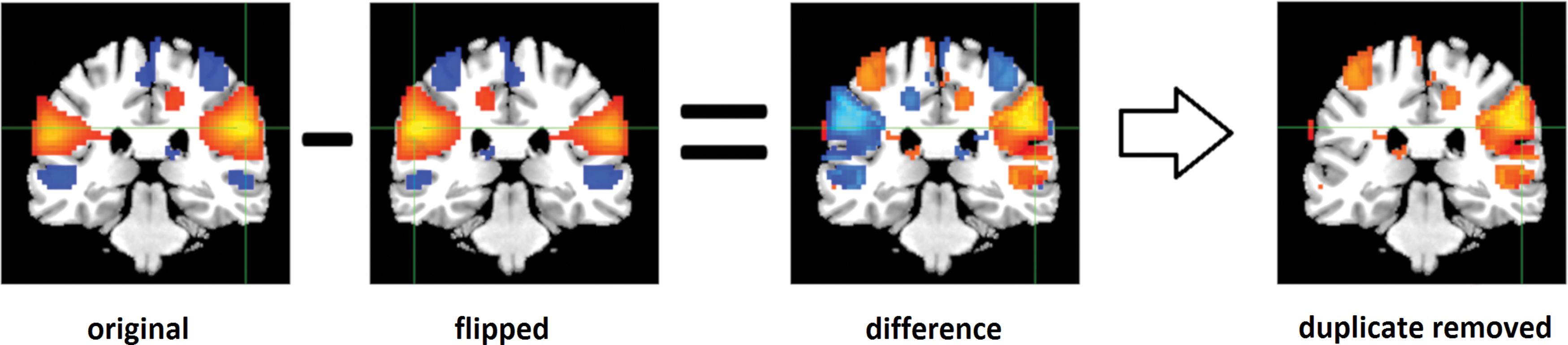
An illustration of calculation of laterality components, the original subject-specific spatial map was flipped, and subtracted from the original SM. Then one sample t-test applied and thresholded the maps at three standard deviations using a Gaussian gamma fit (only positive voxels are shown as the negative voxels are flipped duplicates of the positive voxels). Color images are available online.

The group-level laterality components displayed as t-statistics after thresholding as three standard deviations using Gaussian gamma fit (only positive) and masked with the activation masks. The laterality components are displayed at the same coordinates with the RSNs and grouped with a similar matter to Figure 1. Color images are available online.
In addition to the group-level laterality component, for each RSN of each subject, we calculated a subject-specific laterality component. To do so, for each RSN, we generated a mask using the group-level laterality component and then applied these masks to the components' difference maps.
Laterality metric: laterality index
We summarized the amount of the laterality of a component using a metric, called a global-laterality index. For each RSN, shown in Figure 1, the global-laterality index was calculated by taking the differences between the sum of all intensities on the right and left hemispheres with respect to the sum of all intensities across the brain. The regions with negative intensities on the spatial maps were also included as a positive intensity to the other hemisphere. The global-laterality indexes of RSNs are presented in Figure 4.
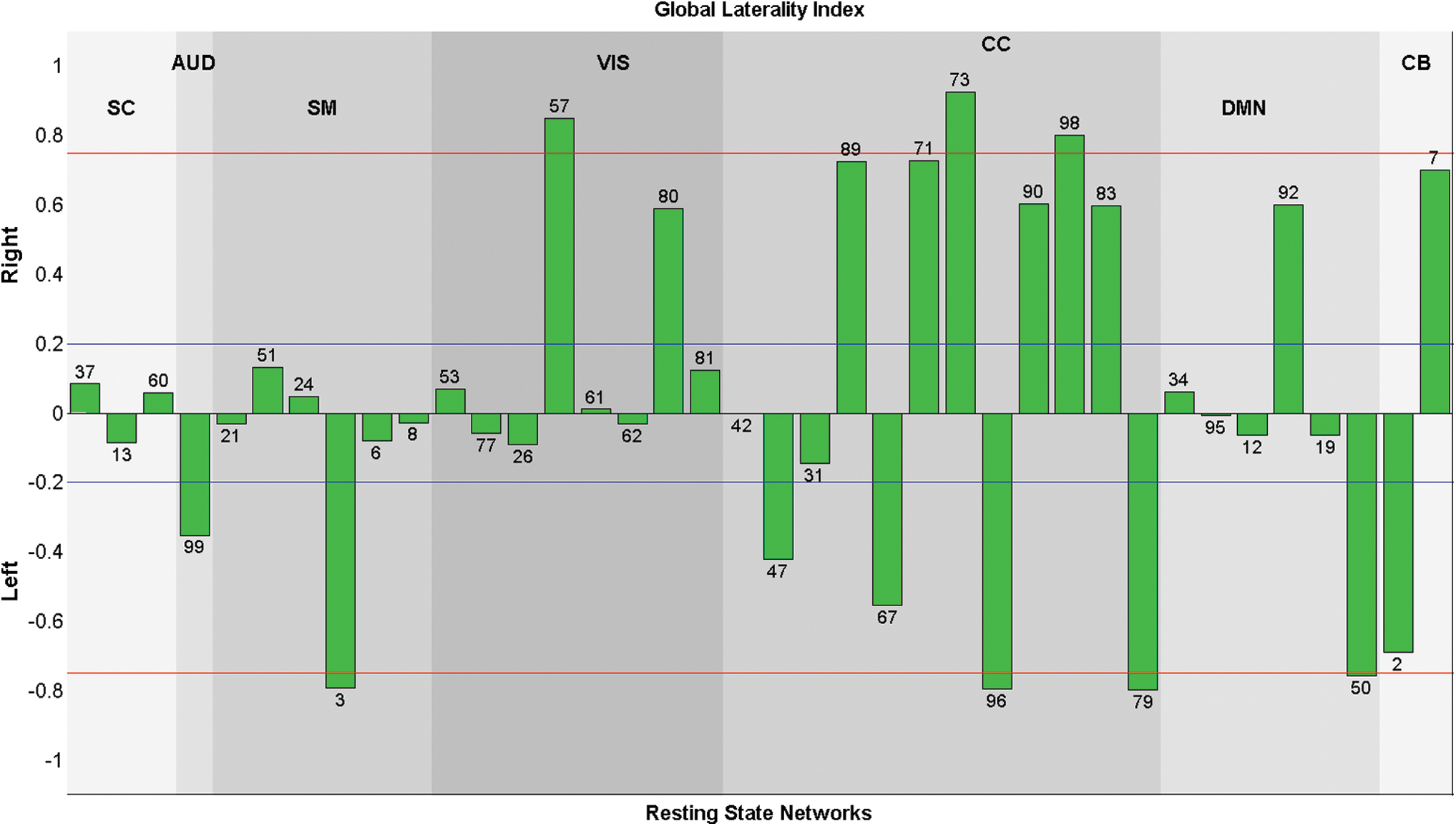
Global-laterality indexes for each component are presented. The indexes that had absolute value >0.75 (red line) were called highly lateralized and the indexes that have absolute value >0.2 (blue line) were called lateralized components. Almost all CC networks were highly lateralized with different directions, while SC and SM networks were mostly bilateral. Color images are available online.
where i ∈ RSN and || is the absolute value operation.
In addition to the global-laterality index, we calculated subject-specific laterality index for each component of each subject. We applied the activation mask that we previously calculated on the subject-specific spatial maps of the each RSN. Finally, we calculated the subject-specific laterality index using the above formula. The mean and the standard deviation of the subject specific laterality index are presented in the Supplementary Material, Supplementary Figure S1.
In this study, we used an improved laterality metric compared with the one used in our previous study (Agcaoglu et al., 2015), which had calculated the laterality metric on the laterality components, rather than the symmetrized components. To facilitate comparisons with our previous work, we recalculated the laterality of the dataset used in Agcaoglu and colleagues (2015) using this new metric and present the results in Supplementary Figure S2.
Voxel-wise modeling of age, sex, handedness, IQ, and behavioral scores effects on laterality components
In addition to evaluating the laterality of the brain networks based on average intensity across subjects, we tested the relationship of lateralization with age, sex, handedness, IQ, and the behavioral score on each voxel on subject-specific laterality components by a linear regression model:
where
Regions that were significantly (FDR corrected) affected in laterality by age. Blue color indicates a region that showed a decrease in laterality with development; while red color indicates a region that showed an increase in laterality with development. FDR, false discovery rate. Color images are available online.
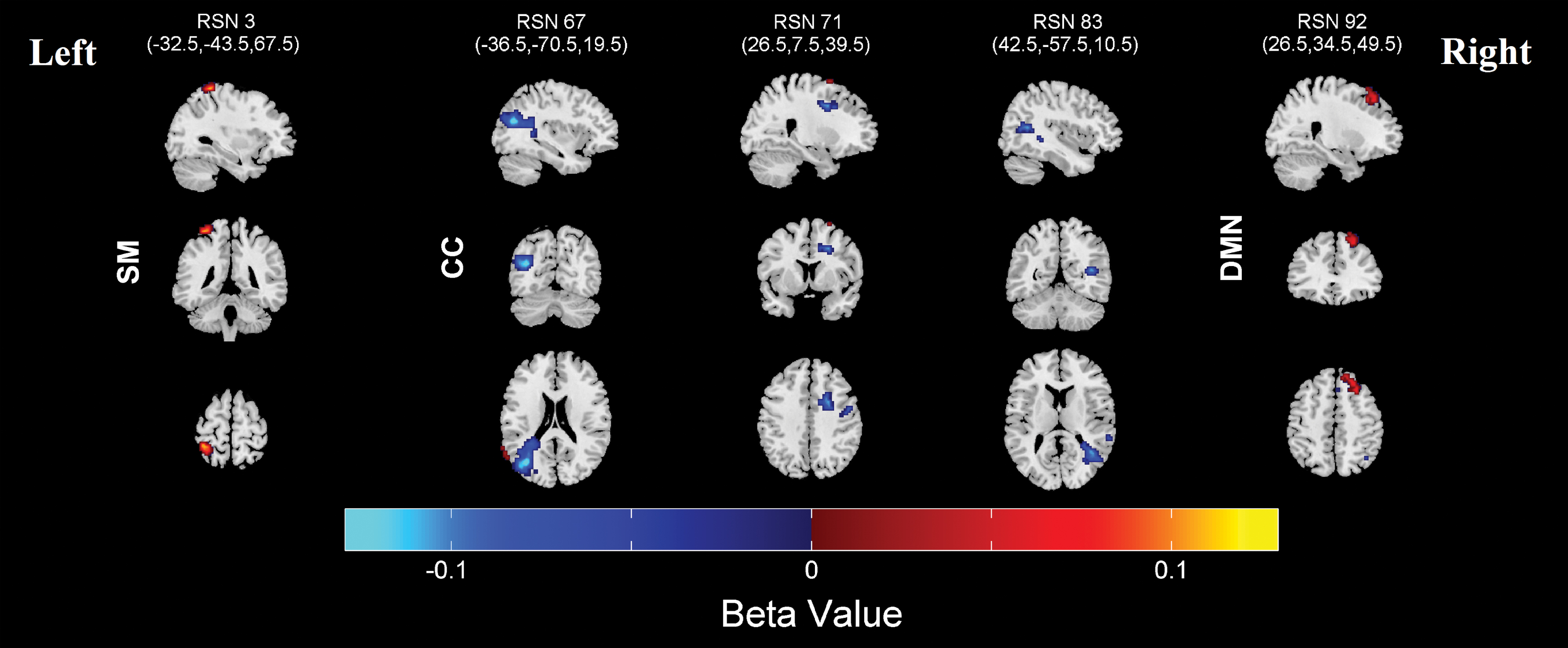
Regions that were significantly (FDR corrected) affected in laterality by sex. Blue color indicates regions that were more lateralized in males, while red color indicates regions that were more lateralized in females. Color images are available online.
Montreal Neurological Institute Regions Displayed in Figure 5, Showing Significant Decrease in Lateralization at the Top and Showing Significant Increase in Lateralization at the Bottom as Subjects Develop
BA, Brodmann area; RSN, resting-state network.
Montreal Neurological Institute Regions Displayed in Figure 6, Significantly More Lateralized in Males at the Top and Significantly More Lateralized in Females at the Bottom
Modeling of age, sex, handedness, IQ, and CBCL score effects on laterality indexes
We also test the effect of age, sex, handedness, IQ, and CBCL scores on subject-specific laterality indexes with a similar linear regression model to that used for the voxel-based analysis and corrected for multiple comparisons (0.05 levels, FDR). The subject motion as the maximum transition was also added as a nuisance variable. Results are presented in Figures 8 and 9 and Supplementary Figures S3–S5.
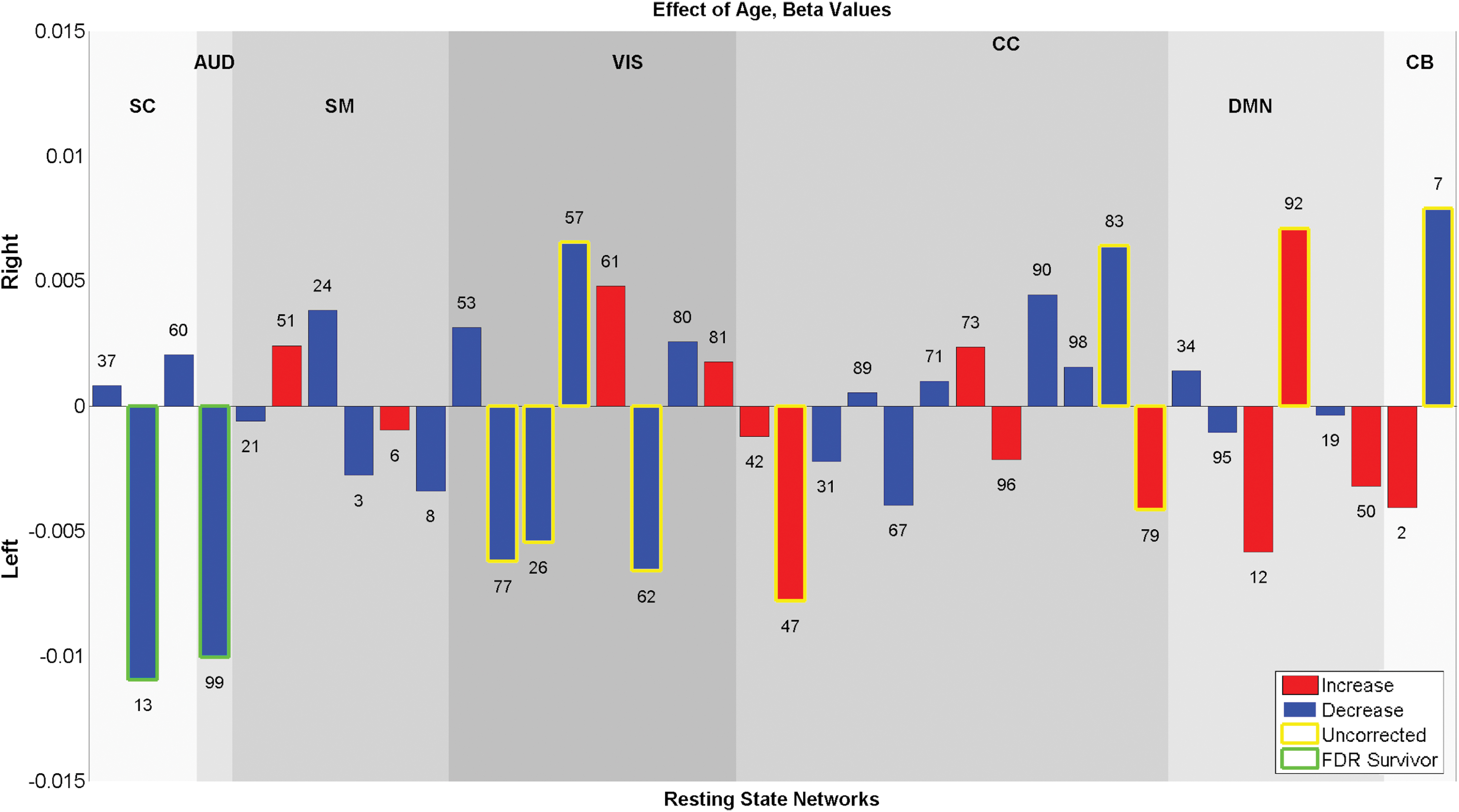
Age effect on laterality indexes displayed as −log10(p)*sign(B), blue bars show the components that have a decrease in laterality with development and red bars show the components that have an increase in laterality with development. SC component #13 and AUD component #99 showed significant (0.05 levels FDR) decrease in laterality. Some other components showed trend level association, for example: CC component #47 and #79 showed an increase in laterality with development (p < 0.05). Color images are available online.
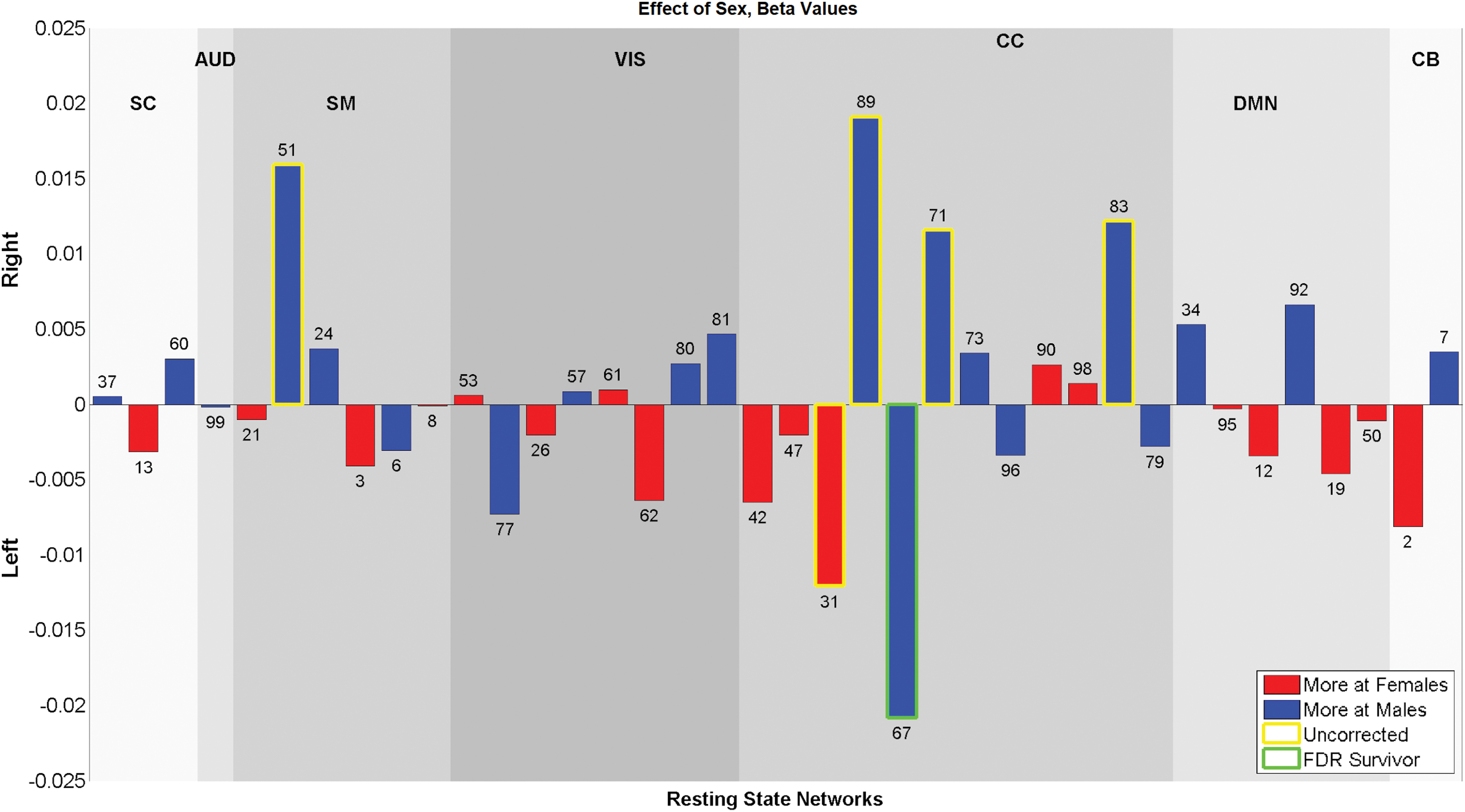
Sex effect on laterality indexes displayed as −log10(p)*sign(B), blue bars show the components that were more lateralized in males and red bars show the components that were more lateralized in females. CC network #67 is the only component that was significantly (0.05 levels FDR) affected by sex and more lateralized in males. Color images are available online.
Results
Laterality indexes
The global-laterality indexes provide a global summary of how the RSNs are lateralized. Heuristically, a component is labeled as “lateralized” if the absolute value of the laterality cofactor is >0.2 and “highly lateralized” if it is >0.75 (Agcaoglu et al., 2015). The global-laterality indexes for each component are displayed in Figure 4. CC components were the most lateralized group, with different components showing lateralization in both left and right hemispheres. CB networks were lateralized, with one left- and one right-dominated component. AUD network was left dominated, whereas SC networks were largely bilateral. SM networks were also mostly bilateral, though with one highly left lateralized network. Some VIS networks were right lateralized, and also other visual networks that were bilateral. DMN components were mostly bilateral, though one showed right lateralized network and one left lateralized network.
Voxel-wise analysis of age, sex, handedness, IQ, and CBCL scores effects on laterality component
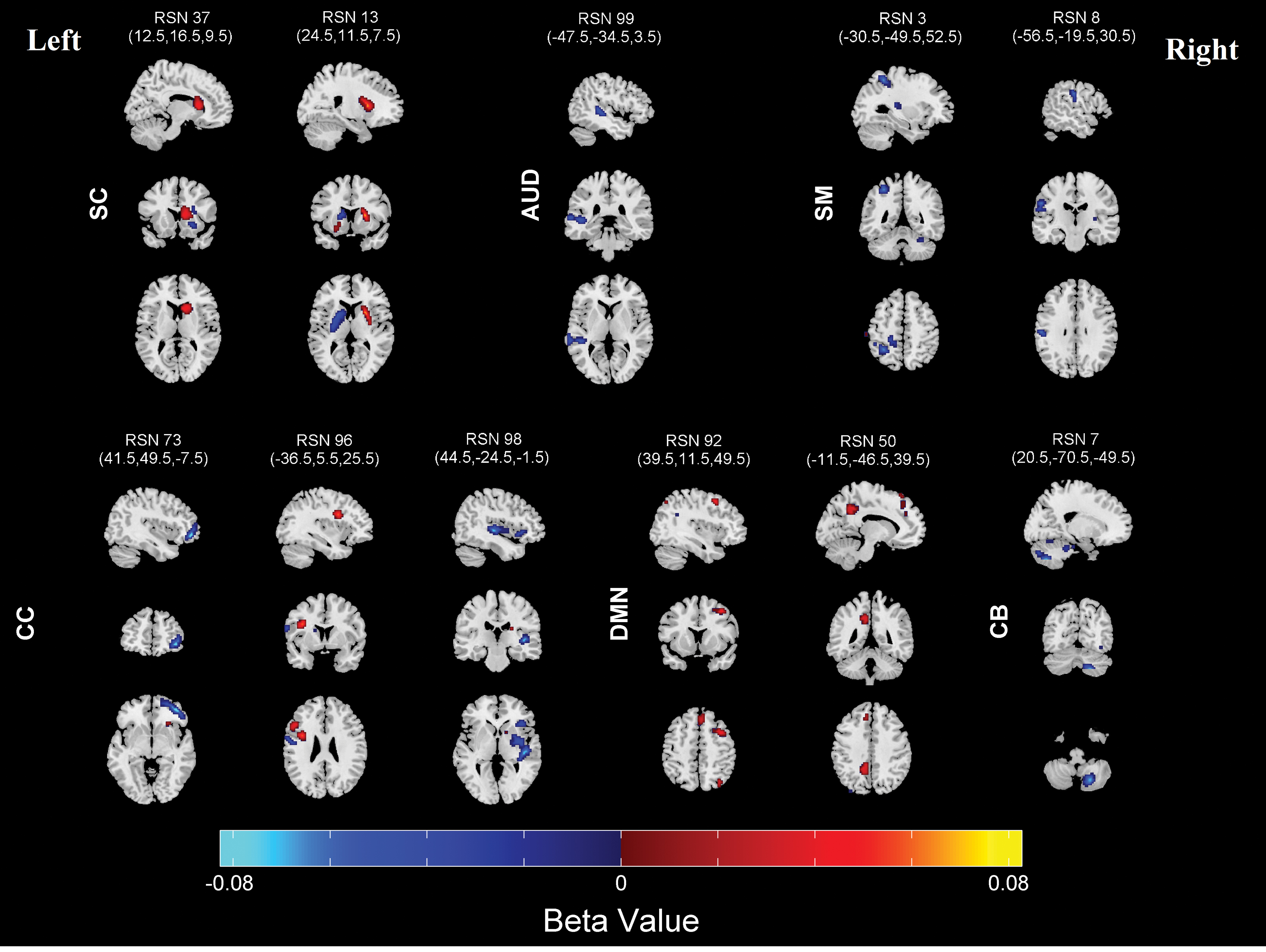
Regions where laterality is significantly associated with age are shown in Figure 5, and the corresponding MNI labels are presented in Table 2. Age was significantly related to laterality showing a relationship between multiple regions in multiple networks; certain networks showed greater laterality with development, while other networks show less laterality with development.
Bilateral SC network #37 shows an increase in laterality in right caudate nucleus and left inferior frontal gyrus; bilateral SC network #13 shows a decrease in laterality with development in left putamen, and an increase in laterality in right putamen and left amygdala. This suggested an overall decrease in laterality of the component with development as well as a shift in activity to the right hemisphere and to the lateral portion of the putamen. With the development, AUD network #99 in left superior temporal gyrus; SM networks #3 in left inferior parietal lobule and right cerebellum, SM network #8 in left postcentral gyrus showed a decrease in laterality. The CC networks were most sensitive to age and have a very complex pattern of age effects. Component #96 had a decrease in laterality in left postcentral gyrus with development. CC components #73 showed a decrease in laterality in right middle orbital gyrus, component #98 showed a decrease in right superior temporal gyrus and right insula lobe. DMN component #92 showed an increase in laterality in right precuneus and right superior medial gyrus, and another DMN component #50 showed an increase in laterality in left precuneus. Also CB component #7 showed a decrease in laterality in right cerebellum.
Sex showed fewer associations with laterality compared with age, with five networks significantly differing regions in laterality after correcting for multiple testing. Regions that were significantly affected by sex are shown in Figure 6, and corresponding regions are presented in Table 3. Three CC components (#67, 71, and 83) showed more laterality in males, while one SM component (#3) and one DMN component (#92) were more lateralized in females.
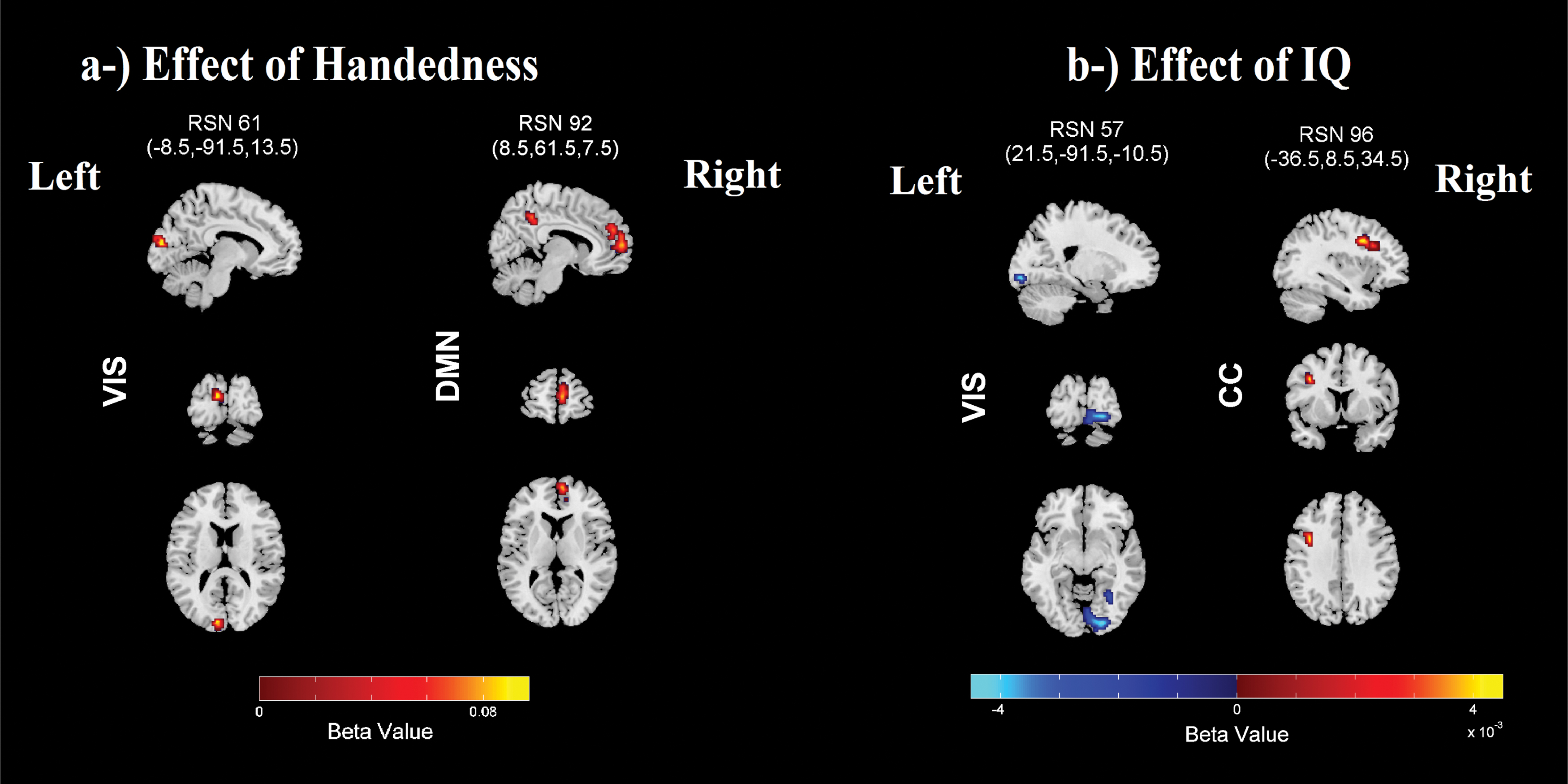
Handedness was significantly associated with the laterality maps in two regions after correcting for multiple comparisons. These regions are shown in Figure 7a, and the corresponding regions are presented in Table 4. VIS component #61 and DMN component #92 were both more lateralized in right-handers.
IQ was significantly associated with two networks after correcting for multiple comparisons, and the regions are presented in Figure 7b and the corresponding MNI regions are listed in Table 4. The highly right lateralized component #57 showed a decrease in laterality in right lingual gyrus with higher IQ, and the highly left lateralized CC network (component #96) showed an increase in laterality in left middle frontal gyrus with higher IQ.
The CBCL score did not show any significant associations with laterality after correcting for multiple comparisons.
Age, sex, handedness, IQ, and CBCL score association with laterality indexes
Age and sex showed significant (0.05 levels FDR corrected) association with the subject-based laterality indexes. Results for age and sex effects are presented in Figures 8 and 9. For the age analysis, SC component #13 and AUD component #99 showed a significant (0.05 levels FDR corrected) decrease in laterality with development. For sex analysis, only CC component #67 showed a significant (0.05 levels FDR corrected) difference between sexes and was more lateralized in males. In addition, some components showed a trend level (p < 0.05) relationship with age and sex. Handedness, IQ, and CBCL scores also showed trend level association with the laterality indexes. These results are presented in Supplementary Figures S3–S5.
Discussion
We examined the lateralization of RSNs and their association with age, sex, IQ, handedness, and behavioral scores in a large dataset of school-aged children using a similar method to Agcaoglu and colleagues (2015). Results from the voxel-based analysis showed that age differences during childhood had a strong association with lateralization. Also, sex was significantly associated with regions in five RSNs, and IQ and handedness were associated with regions in two RSNs. Consistent with our previous adult study (Agcaoglu et al., 2015), lateralization indexes showed lower associations with demographic factors compared with the voxel-based analysis in this developmental period of childhood as well.
We observed a decrease in laterality in the left putamen and increasing laterality in the right putamen and right caudate nucleus with development. Note that the putamen is associated with reinforcement learning and motor control, language and speech production (Vinas-Guasch and Wu, 2017). During developing age, this subcortical region might undergo critical developmental changes and therefore, the laterality effects might not be global within this region. Future work could directly evaluate the relationship of these laterality effects with speech and motor behavior.
Consistent with our previous work of adult (Agcaoglu et al., 2015), CC networks were the most lateralized as well as the most sensitive to age and sex effects in children. We mostly find decreased laterality with development, only CC #96 shows an increase in laterality in inferior frontal gyrus region. Gracia-Tabuenca and colleagues (2018) also investigated age effects in the hemispheric asymmetry in children from 6 to 10 years of age with a small sample size of 60 and did not report any significant effect in their functional connectivity-based laterality approach; this could be due to small sample size and small age difference. Gracia-Tabuenca and colleagues (2018) also reported a negative relationship between homotopic connectivity of lingual gyrus and visual long-term memory performance. In our study, we found that visual network, lingual gyrus, was more bilateral with the increasing IQ. Koyama and colleagues (2017) identified hemispheric asymmetries in the middle frontal gyrus, with left middle frontal gyrus showing association with literacy, and right middle frontal gyrus showing association with numeracy in their multivariate distance matrix regression based intrinsic functional connectivity study in healthy adults. We found an increase in lateralization in the left middle frontal gyrus region with increasing IQ. This is particularly interesting because in our study, the SON-R test with mosaics and categories was used to measure IQ, which is a nonverbal test, yet it showed a significant association with a literacy-dominated region. A possible reason for this might be the fact that our participants were ages 6 to 10 years, where education is mostly focused on reading and writing skills.
When we compared the voxel-wise analysis, results between adult study (Agcaoglu et al., 2015) and children study, the most remarkable difference was a decrease in laterality with increasing age in adults versus both decreases and increases in laterality with development in children. This provided evidence supporting nonlinear age effects in laterality. However, further examination with nonlinear models is required.
While voxel-based analysis was more sensitive to the covariates compared with the laterality indexes, the results were not contradictory. The results were dependent on the structure of the component. For example, component #73 was one sided and was >90% right lateralized, therefore even though the volume of the spatial maps varied between subjects with different age, the laterality index would not reflect that because it would be ∼1 for any case. On the contrary, we observed significant decrease in laterality with development in components #13 and #99 in both analyses. This was because of the fact that components #13 and #99 were bilateral. Results thus suggest that voxel-based analysis may be more sensitive for analyzing the relationship with covariates.
When we compared the laterality indexes from this study of children (Fig. 4) and our previously reported study of adults (Supplementary Fig. S2) (Agcaoglu et al., 2015), we observed interesting similarities and differences. For both datasets, CC and attention-frontal networks were the most lateralized networks. Also, VIS networks were mostly bilateral with having one or two components right dominated. In contrast, the AUD network was lateralized in children while bilateral in adults, which was consistent with our findings suggesting significant decrease in laterality in AUD network in laterality indexes and voxel-based method in children. The DMNs were mostly completely bilateral for both groups with only one lateralized component. Both datasets had bilateral SC networks. In children, we observed SM networks were mostly bilateral with one left lateralized component, while in adults we had SM networks as pair of one highly left lateralized and one highly right lateralized network, which was consistent with the fact that left hemisphere controls right side of the body and right hemisphere controls left side of the body (Janssen et al., 2011). However, in children, sensorimotor networks were not separated into left and right lateralized networks, presenting only one highly left lateralized network.
This study highlights that the extent of lateralization of functional connectivity is associated with age, as well as general cognitive ability. Future work should explore to what extent lateralization of RSNs is associated with aspects of atypical development, such as psychopathology. In addition, direct comparison across larger age ranges may provide additional insights into the changes in laterality across the life span.
Limitations
Some limitations should be considered while interpreting the results. First, this is a cross-sectional study examining a snapshot of age-related associations with lateralization. Longitudinal data will allow for tracking within-individual change over time. Also, based on prior work, we used a gICA model order of 100; however, examining the laterality with lower and higher gICA model order may also be interesting. Based on our extensive previous work on ICA analysis, we know that including more (or less) components in the model will enable us to evaluate the changes at different spatial scales. However, such an analysis is rather extensive and beyond the scope of this particular work. In addition, although, we applied a second normalization step and warped the components to a symmetrical template to account for structural asymmetries between hemispheres, it should be noted that some additional structural asymmetries might still be present.
Conclusion
In summary, we analyze the lateralization of RSNs in a large fMRI dataset consisting of 774 children in age ranging from 6 to 10 years. Our results indicated that lateralization shows strong association with age in several networks as well as associations with sex, IQ, and handedness in some networks. CC networks were the most sensitive networks to both age and sex effects, had components both increases and decreases in laterality with development in a complex pattern. Also, auditory network and sensorimotor regions showed a decrease in laterality with development, while regions including dorsolateral frontal cortex (Brodmann area L9 and L46) showed an increase in left lateralization with development. Our results provide evidence that association of laterality and age is nonlinear and the patterns are very complex in the ages (6–10) where rapid changes occur in brain function and structure.
Footnotes
Authors' Contributions
T.W., R.L.M., and H.T. designed the study; wrote the protocol; and collected the rs-fMRI data. B.R. preprocessed the data and identified RSNs. O.A. and V.D.C. processed the data and designed the experiment, O.A. ran the analysis, wrote the first draft of the article. All authors contributed to and approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported in part by National Institutes of Health (NIH) grants P20GM103472, R01MH121101, P20GM130447, and R01EB020407.
Supplementary Material
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.