
Abstract
Background:
It is well recognized that semantic processing and auditory repetition facilitate subsequent naming of pictures. However, the neurocognitive mechanisms that underpin these facilitation effects remain unclear.
Materials and Methods:
The current study utilized a dynamic causal modeling (DCM) approach to examine high-density electroencephalographic (128-channel EEG) recordings and investigate connectivity modulations during facilitated naming of pictures in 18 healthy older adults (mean age 61.50 years). Source reconstruction of event-related potentials was performed in two specific time windows, (1) 150–250 msec and (2) 300–500 msec, to establish the timescale of significant cortical activations present during participation of semantic and phonological tasks. Hypothesis-driven DCM of source-activated regions was tested to ascertain which model most likely explained the semantic and phonological conditions, respectively.
Results:
DCM results indicated that a common cortical network comprising dorsal and ventral cortical connections best explained EEG task data derived from repetition and semantic tasks. For repetition (phonological) tasks, this model featured long feedback, bidirectional connections from inferior frontal gyrus (IFG) to occipitotemporal areas. Semantic tasks were most plausibly explained by a model that featured a self-inhibiting connection over the IFG only.
Conclusions:
Findings from this study reveal that a common cortical model comprising pathways that include dorsal and ventral regions is appropriate for characterizing EEG naming facilitation data, and that distinct cortical connections explain differences between semantic and auditory repetition processes. These models could be repurposed for naming facilitation paradigms in patients with language difficulties to optimize prediction and responsiveness to such paradigms.
Impact statement
The combination of semantic (word-level) and phonological (sound-level) processing in the cortex facilitates one of the most robust responses—the naming of pictures. Here, dynamic causal modeling of high-density electroencephalography during facilitated naming tasks revealed a model consisting of common dorsal and ventral connections that best explained response to semantic and phonological stimuli. Within this cortical network, phonological facilitation involved a long-range connection from inferior frontal gyrus (IFG) to occipitotemporal regions, whereas semantic facilitation contributed to self-inhibition of the IFG. The IFG is therefore a key region mediating cortical activity when switching between phonological and semantic conditions.
Introduction
Word finding difficulties (anomia) are a common symptom of aphasia, and both phonological and semantic treatment tasks have shown the capacity to improve such impairments for people poststroke (Hickin et al., 2002; Kiran, 2008; Leonard et al., 2008; van Hees et al., 2013). Studies suggest that the mechanisms that underpin successful treatment of anomia may be the same as those that underpin word priming in unimpaired naming (Nickels, 2002). The present study utilizes high-density electroencephalography (EEG) and dynamic causal modeling (DCM) to explore these mechanisms further, by examining cortical activity differences between semantic and phonological facilitation of picture naming in healthy older adults.
Neuroimaging evidences of naming facilitation
The process of spoken word production during picture naming is generally recognized to involve a lexical-semantic and a phonological stage. During the lexical-semantic stage, a lexical representation corresponding to the picture is selected, and this abstract lexical unit is then mapped to its phonological form (Levelt et al., 1999). A mapping operation between the lexical-semantic and phonological stages is assumed to provide a link between word meaning and word form (Nickels, 2001). Naming facilitation refers to the higher accuracy and/or faster response times to picture naming subsequent to a facilitation task. Accordingly, the presence of such effects suggests that one or more of the processes associated with spoken word production and recognition have been facilitated by the performance of the prior task, contributing to the efficiency of word retrieval processes.
To date, research on the facilitation of naming in healthy adults is limited. In a functional magnetic resonance imaging (fMRI) study, Heath et al. (2012c) examined the neural correlates of semantic facilitation in healthy older adults. Semantic facilitation involved answering a spoken yes/no question (e.g., does it grow on trees?) relating to each picture, and the effects of both short-term (minutes) and long-term (days) facilitation were examined. Critically, the semantic task did not involve overt production of the target item or hearing the name of the target item, thereby excluding direct exposure to the phonological form during task performance. In addition to faster naming latencies for short- and long-term facilitated items relative to unfacilitated items, the authors found that distinct mechanisms underpinned short- and long-term facilitation. Short-term effects were associated with increased neural activity for facilitated items in the right lingual gyrus (extending into the precuneus region) and the left inferior occipital gyrus (extending to the left fusiform gyrus). The authors concluded that the short-term facilitation effects were driven primarily by brain mechanisms relating to automatic recognition or recall of the visuo-perceptual representations associated with object priming, rather than the actual lexical-semantic mechanisms. In contrast, long-term facilitated items were associated with repetition suppression effects in the middle temporal gyrus, suggesting that longer term facilitation was indeed driven by effects at the semantic and/or lexical-semantic level.
In a separate fMRI study of healthy older adults, Heath et al. (2012b) examined the neural correlates of short- and long-term naming facilitation from auditory repetition of the target word. Long-term facilitated items were associated with repetition suppression effects in the left middle temporal gyrus (relative to unfacilitated items), an area known to be associated with lexical selection (Hickok, 2009; Indefrey and Levelt, 2004), as well as the left superior temporal gyrus (relative to short-term facilitated items), an area known to be associated with phonological access (Graves et al., 2008; Price, 2000). These findings suggested that long-term effects of auditory repetition may be driven by a strengthening of semantic-phonological connections. The authors also acknowledged that the repetition suppression effect for long-term items relative to short-term items in the left superior temporal gyrus may alternatively reflect a repetition enhancement effect within that region for short-term facilitated items. As such, it is possible that short-term facilitation via auditory repetition is also mediated by phonological processing. However, short-term facilitation was primarily associated with a reduction in activity within the left inferior occipital-temporal region. Accordingly, the short-term effects of auditory repetition appeared to be driven primarily by semantic and object recognition processes, suggesting a degree of semantic access when simply repeating a name in the presence of the object.
Dynamic causal modeling for semantic and phonological facilitation
Although fMRI research has identified how naming facilitation techniques alter neural activity within specific regions of the brain, computational measures of effective connectivity of such imaging paradigms, including EEG, have the capacity to further elucidate the neural mechanisms that underpin naming facilitation. DCM offers the means to pursue more detailed hypothesis-based connectivity inferences by testing the effect of one region's neural activity on another (Friston, 2011).
The combination of fMRI with DCM has been used to explore the effective connectivity associated with various language tasks, including naming. Hoyau et al. (2018) examined picture naming in healthy younger and older adults. The researchers identified bidirectional excitatory interactions between the left inferior frontal cortex (IFC) and the left lateral posterior temporal cortex (LTC), as well as an inhibitory unidirectional interaction between the left IFC and left occipital cortex. While the inhibitory connection between the left IFC and left occipital cortex was also maintained in the healthy older adults, the connection between the left IFC and left LTC was not. Instead, older adults displayed a bidirectional interaction between the left IFC and the left middle temporal cortex (MTC). The authors posited that the difference in older adults could potentially indicate that older adults use a different strategy to younger adults to access semantic memory during picture naming.
In another fMRI DCM study, Meier et al. (2018) investigated the effective connectivity within a left hemisphere network during a semantic feature detection task in both people with aphasia and healthy adults. For both groups, DCM results indicated that the left middle frontal gyrus (MFG) modulated the left posterior MTG, possibly reflecting assistance with the rapid activation of candidate semantic features during task performance, and that the left MFG also modulated the left inferior frontal gyrus (IFG) pars triangularis, potentially indicating involvement in the correct selection of the semantic features relevant for a target item. Besides fMRI, DCM application to EEG-based studies of language processing provides further insight into effective connectivity of cortical activity associated with the semantic and phonological processes of written words (Yvert et al., 2012). In a similar way, DCM can also be used to compare the connectivity pathways associated with semantic and phonological naming facilitation, which has not been investigated to date.
The present study aimed to compare the effects of short-term phonological and semantic facilitation on picture naming in healthy older adults using DCM analyses of high-density EEG recordings. We used DCM to test a hypotheses-driven model space that examined cortical activity modulations, within an event-related potential (ERP) framework, to assess engagement of purported dorsal pathways during phonological conditions and ventral pathways during semantic conditions. To test this, we assessed model variations of dorsal, ventral, and combined pathway models to assess whether cortical activity, measured during participation of short-term naming facilitation tasks, would likely subscribe to a model with both dorsal and ventral streams of cortical processing (Hickok and Poeppel, 2004). The study also aimed to determine whether these model variations would consist of longer cortical connections consistent with bottom-up processing from visual to frontal gyri, and vice versa top-down control from prefrontal cortices back to visual areas.
Materials and Methods
Participants
Participants were 18 healthy older adults (10 females) with a mean age of 61.50 years (standard deviation [SD] 11.59) and a mean education of 14.83 years (SD 4.18). All participants reported as right-handed, with English as a first language, normal hearing, and normal or corrected to normal vision. Based upon screening conducted using the Mini-Mental State Examination (Folstein et al., 1975) and the Geriatric Depression Scale-short form (Sheikh and Yesavage, 1986), no participants presented with cognitive impairment or depression. The study was approved by the Medical Research Ethics Committee at the University of Queensland, and all participants provided written informed consent before participation (project number: 2003000010).
Experimental design and statistical analysis
The picture stimuli consisted of 224 items from the International Picture Naming Project Database (Szekely et al., 2004), which had an average name agreement of 96.8% (SD 4.1%; minimum = 86%). These stimuli were subsequently assigned to 3 conditions: unfacilitated (96 items), facilitated (96 items), and fillers (32 items). The facilitated and unfacilitated conditions were matched for number of phonemes, syllables, percent name agreement, and mean response time (Szekely et al., 2004) (ps > 0.05). Stimuli were also matched on CELEX frequency (Baayen et al., 1995), obtained via N-watch software (Davis, 2005).
The paradigm consisted of two experiments: (1) semantic facilitation and (2) auditory repetition facilitation, hereby referred to as phonological facilitation. The same stimuli were used for both experiments, but with a different pseudorandomized order of stimuli that was held constant for each participant. The two experiments were held during separate testing sessions at least 1 week apart, and the order of experiment completion (semantic/phonological) was counterbalanced across participants.
For both experiments, the stimuli were divided into 16 blocks of trials, with each block consisting of a facilitation phase followed by a naming phase. Each facilitation phase contained 8 pictures, comprising 5–7 pictures from the facilitated condition and 1–3 pictures from the filler condition. The subsequent naming phase contained 14 pictures, consisting of the same 8 items presented during facilitation, together with an additional 6 pictures from the unfacilitated condition. There was a lag of 7–11 intervening items between presentation of a picture in a facilitation phase and its subsequent presentation in the naming phase.
In the semantic experiment, pictures presented to participants during the facilitation phase were accompanied by a spoken yes/no question. These questions were based on Garrard et al. (2001) semantic attributes, probing sensory (e.g., does it bark?), functional (e.g., is it used for cooking?), or encyclopedic (e.g., is it found on a farm?) attributes about the picture, ensuring semantic retrieval was required for answering the question (i.e., the answer could not be determined simply via analysis of visual information within the picture). As per Heath et al. (2012a), the auditory semantic questions for each stimulus item were digitally recorded in a soundproof recording studio at 44,100 Hz, mono, 32 bit, and spoken by a female speaker. The correct response for all questions relating to pictures from the facilitated condition was “yes,” to ensure that engagement of relevant semantic properties was required to be associated with each stimulus item for the provision of a correct response. In contrast, the correct response for questions relating to pictures from the filler condition was always “no,” to ensure a level of unpredictability regarding the correct response.
Participants were asked to respond to each question (yes/no) via button press on a response pad. No feedback was provided regarding response accuracy, and the questions never referenced the word form of the relevant item. For example, for the picture item “ball,” participants were asked “Can it bounce?” rather than “Can a ball bounce?” This approach ensured that participants did not hear, and were not required to produce, the phonological word form of the target item during the semantic facilitation experiment. In the phonological experiment, pictures presented to participants during the facilitation phase were accompanied by the spoken name of the picture. Spoken names were recorded in the same manner as those for the semantic questions. After hearing the spoken name, participants were required to repeat the name of the picture into a microphone.
Both experiments were coded using E-prime 2 (Psychology software tools), and the specific sequence of events for each trial was as follows (Fig. 1). During the facilitation phase, a fixation point (+) appeared in the center of the computer screen for 500 msec. The picture then appeared in the center of the screen and either a semantic question (for semantic facilitation) or the picture name (for phonological facilitation) was simultaneously presented via speakers. The picture remained on screen for 3000 msec or until the participant provided a response (yes/no button press for the semantic condition; spoken response for the phonological condition). The next trial was then initiated automatically after 1500 msec.
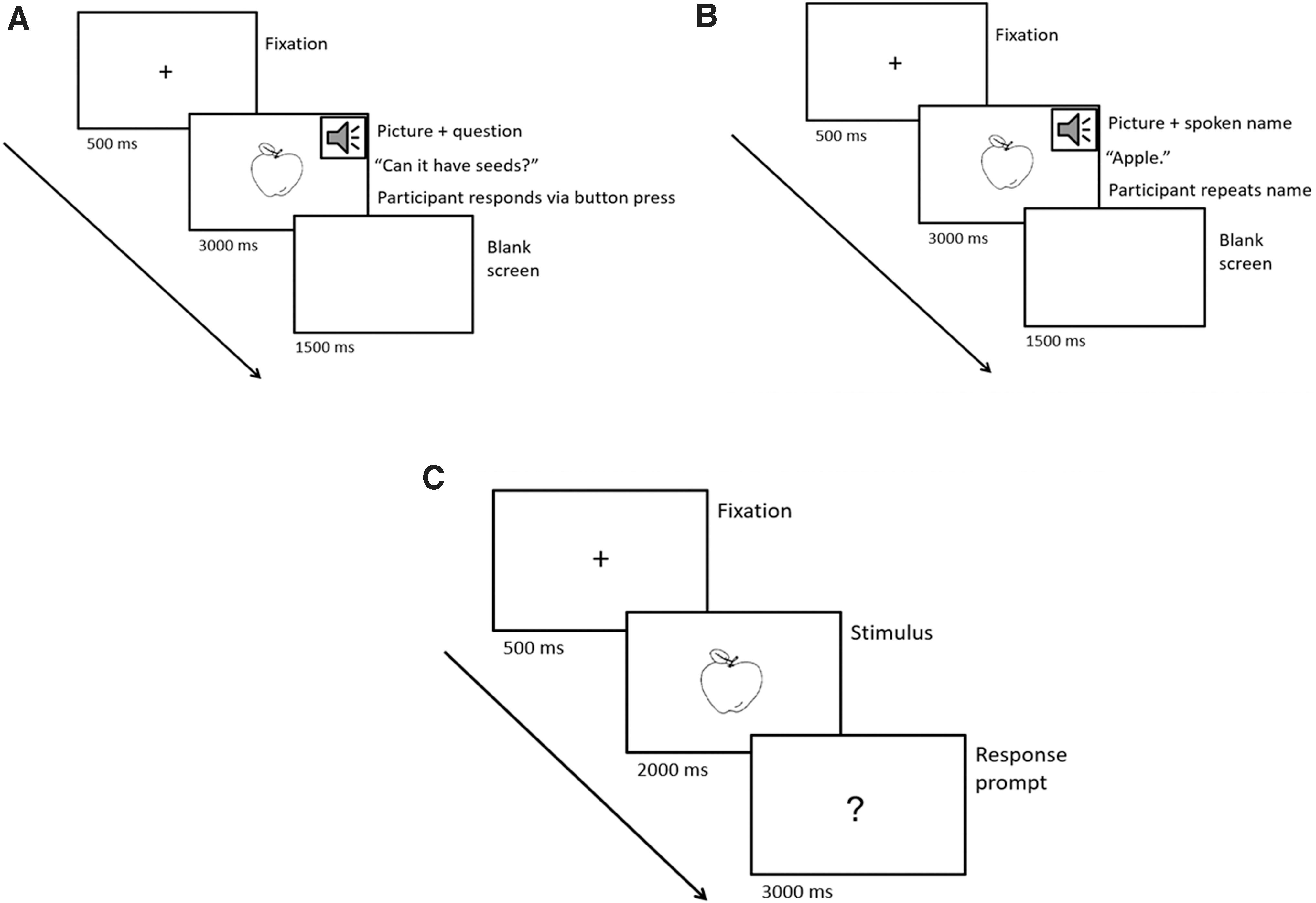
An illustration of the trial events for
Immediately following completion of each facilitation phase, the subsequent naming phase commenced. At the beginning of each trial, a fixation point (+) appeared in the center of the screen for 500 msec. The target picture then appeared in the center of the screen for 2000 msec, followed by a question mark. As per Laganaro et al. (2008), participants were asked to prepare the word but to wait until the question mark appeared before providing a response. The question mark remained on screen for 3000 msec or until the participant provided a spoken response, and the next trial was then manually initiated by the researcher after ∼2 sec. The researcher maintained manual control over the trials in this manner so that participants could be given reminders (if necessary) to wait for the question mark before responding and/or to speak more loudly into the microphone. Participant's verbal responses during the naming phase were recorded on an Olympus (DS-40) digital voice recorder to allow later transcription of responses for verification of naming accuracy. Before each facilitation and naming phase, participants were reminded of the task instructions both verbally and via written instructions on the computer screen, and participants were provided with a short rest break after every four blocks of trials.
EEG acquisition and preprocessing
EEG data were recorded continuously during task performance with the EGI 300 Geodesic EEG system (Electrical Geodesics, Inc.) using a 128-channel HydroCel Geodesic Sensor net at a sampling rate of 250 Hz. Electrode impedance was kept below 50 kΩ, which is acceptable when using high-impedance amplifiers (Ferree et al., 2001). Following acquisition, EEG data were band-pass filtered between 0.1 and 20 Hz (Butterworth filter). The EEG data from the naming phases (i.e., postfacilitation) were subsequently segmented from −100 to 600 msec relative to picture onset (i.e., when participants were asked to mentally prepare the word). Ocular artifacts and noisy channel data were interpolated from the remaining channels using spherical splines. Any subsequent trials that still consisted of ocular artifacts or that contained more than 20 bad channels (defined as reaching amplitudes greater than 200 mV) were excluded from analysis. In addition, trials where participants named an item before the appearance of the question mark (2.19% of trials) were also excluded, as were a small proportion of trials relating to experiment error. Overall, the average number of trials available for analysis for each condition totaled to the following: 78.7 for phonological facilitation, 85.3 for phonological unfacilitated; and 76.7 for semantic facilitated and 80.2 for semantic unfacilitated. The data were rereferenced to an average reference and baseline corrected to the 100 msec period before picture onset.
Source imaging
SPM12 was next used to source reconstruct ERP data (Litvak et al., 2011). Cortical source densities were obtained from semantic and phonological condition ERPs using the in-built three-dimensional source reconstruction routines, with electrode positions coregistered to a canonical head model composed of 8196 vertices based on the MNI brain (corresponding structural image
Dynamic causal modeling
Effective connectivity between cortical sources was examined via DCM for ERP in SPM12. Briefly, the DCM approach uses a neurobiologically generated neural mass model, which consists of distributed, laminar neural architectures that denote excitatory pyramidal cells, and spiny stellate and inhibitory interneurons. The ERP model for DCM is a modified version of the canonical Jansen-Rit equations (David et al., 2006; Kiebel et al., 2007) for cortical activity, which enables us to examine excitatory changes in ERPs during task states. As a hypothesis-driven approach, the DCM ERP approach relies on several connectivity rules that allow us to model reciprocal connections between cortical sources, providing a template for which an optimal model can best explain cortical dynamics according to a given task state. Within this model, forward connections ostensibly model “bottom-up” processes, in which connections originate in agranular layers of the cortex and terminate in granular layers (e.g., layer IV). Backward connections for this modeling approach reflect “top-down” modulatory processes, where connections target agranular layers. Self-inhibiting (also known as intrinsic) connections can also be included in this model, in which regions that typically demonstrate recurrent cortical activity have self-inhibiting (or intrinsic) connections, which provides insight into neuronal adaptation processes occurring from repeated exposure to task-related information. For this study, forward, backward, and self-inhibiting connections were modeled within an ERP window of 0–600 msec, to capture N200 and N400 components.
Statistical analyses of source activity
For source analysis of N200 and N400 components, stringent multiple comparison testing was applied via cluster-level thresholding (activations within a 10 mm radius), in which cluster thresholds were set at greater than 500 voxels. Family-wise error correction (FWE) of source images was then applied to ascertain statistical significance, set at p FWE<0.05. A one-sample t-test was performed over the group for each of the semantic and phonological tasks, respectively, so that statistically significant source-activated regions were identified. For DCM analysis, Bayesian model selection, via a random-effects analysis, was used to assess evidence of all DCM model hypothesis tests, for the semantic and phonological conditions. Using this statistical inference, the “winning” model was determined by calculating the model with the highest exceedance probability, ensuring that model-generated data were optimally fit for any given (randomly chosen) subject. Variations in the model space were also subsequently examined for highest model exceedance probabilities to determine which models of long feedback and self-inhibiting connections best explained the data.
Results
Behavioral results
As expected for healthy adults, naming accuracy was near ceiling, exceeding 97% accuracy for both facilitated and unfacilitated items in both experiments (semantic facilitation experiment, 97.74% and 99.31% accuracy for facilitated and unfacilitated items, respectively; phonological facilitation experiment, 99.25% and 98.55% accuracy for facilitated and unfacilitated items, respectively). Due to the high accuracy of performance, no statistical analysis of behavioral data was undertaken. Similarly, correct performance during the facilitation tasks (i.e., auditory repetition or response to semantic questions) was also high. During the semantic facilitation phases, participants responded incorrectly to only 2.01% of the questions. During auditory repetition, only one error was made by one participant.
Source imaging results
Group-level, one-sample t-tests on semantic and phonological EEG task conditions revealed significant clusters of cortical activity at N200 and N400 ERP windows. Figure 2 indicates statistically significant source-activated regions during facilitation of semantic and phonological items. Cluster-level thresholds were enforced via FWE (p FWE<0.05). These activations formulate our ROI for DCM analysis. Based on our source imaging approach, our DCM framework used six ROIs (Table 1 and Fig. 1). These ROIs included the following: (1) occipitotemporal junction (OT), (2) inferior parietal lobule (IPL) covering the angular gyrus, (3) posterior superior temporal and (4) anterior middle temporal gyri, (5) inferior and (6) MFG.
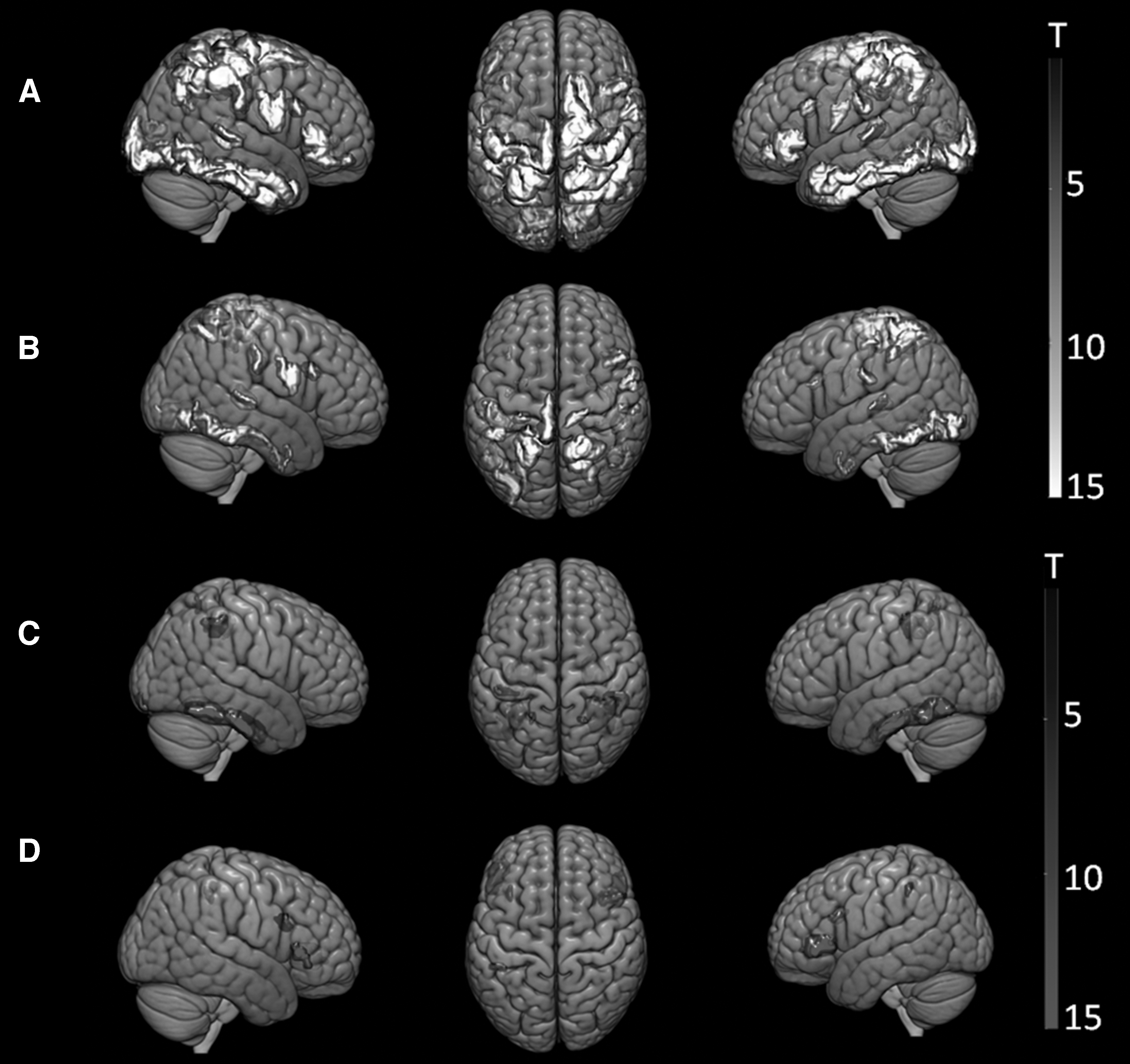
Cortical source activity during facilitation of semantic and phonological tasks at two time windows: N200 (150–250 msec) and N400 (300–500 msec). A one-sample t-test of cortical sources at these windows indicates significant regional activity for the
Regions of Interest for Dynamic Causal Modeling Analysis Indicated by Cortical Source Imaging (p FWE < 0.05)
ROIs include: OT, IPL, pSTG, aMTG, IFG, and MFG.
BA, Brodmann's area; FWE, family-wise error correction; IFG, inferior frontal gyrus; IPL, inferior parietal lobule; MFG, middle frontal gyrus; MNI, Montreal Neurological Institute; OT, occipitotemporal junction; pSTG, posterior superior temporal gyrus; ROIs, regions of interest.
Dynamic causal modeling
Using a Bayesian statistical inference approach, our DCM analysis considered 48 models to represent the most plausible variations of dorsal and ventral routes of cortical language processing. These 48 models included 3 bilateral variations of modeled connectivity pathways of regions identified (Table 1). These model variants include the following: (Variant 1) dorsal and ventral pathways, that is, forward and backward connections between ROIs; (Variant 2) a more dorsal route of connections; and (Variant 3) more ventrally connected components. We further delineated our model space to include (1) no long-range feedback connections, (2) feedback of long-range connections from IFG to IPL, (3) feedback of long-range connections from IFG to OT, and (4) long-range connections from IFG to both IPL and OT. In addition, we included self-inhibiting connections modeled over the OT, IPL, and IFG individually and a separate model variant with OT, IPL, and IFG together, for each model variant represented in Figure 3.
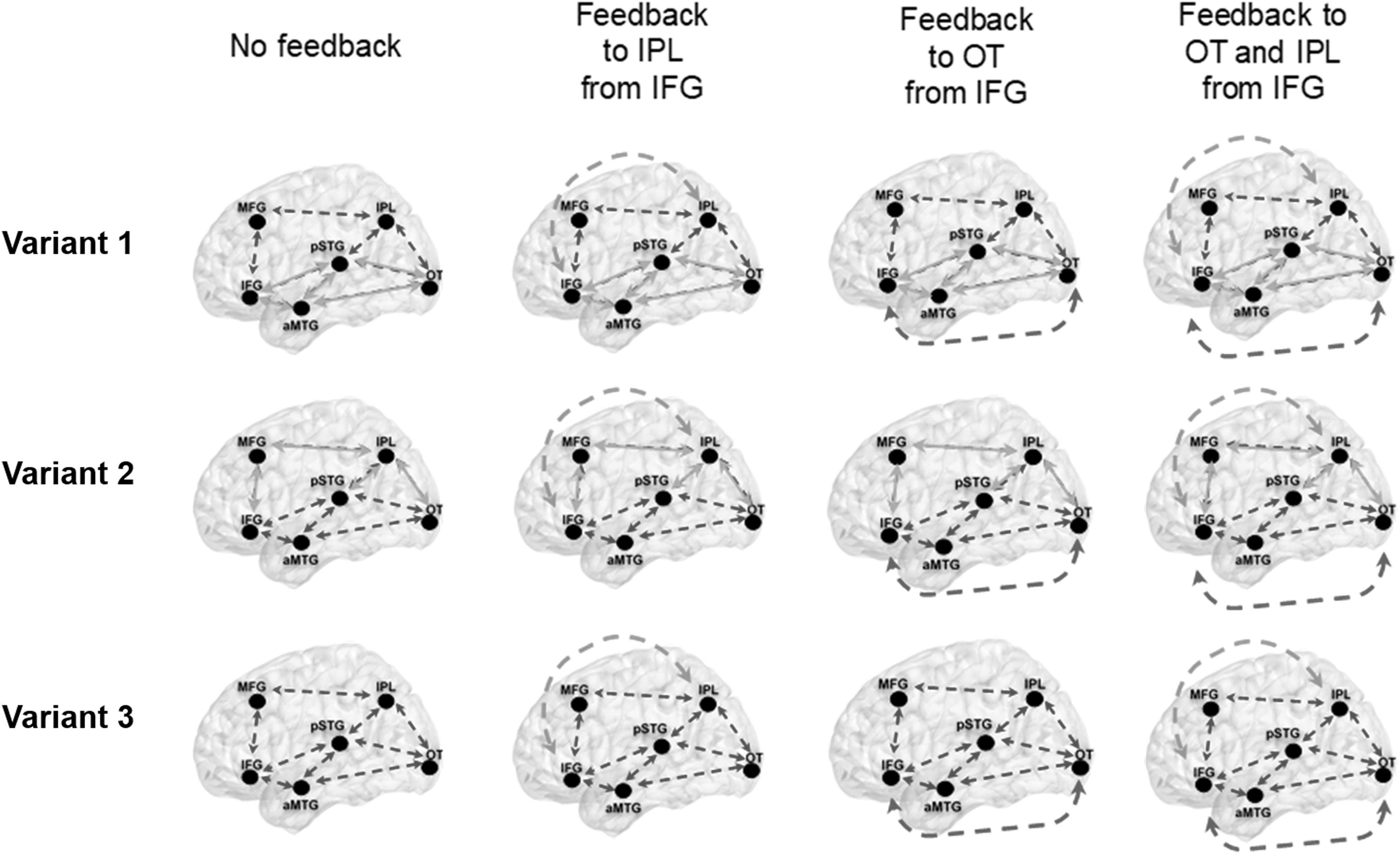
The model space tested for this study via DCM analysis. In total 48 models are tested, in which the above 12 models have three different model variants. These model variants included testing of self-inhibiting connections on each OT, IPL, and IFG, with one set of models consisting of all three regions. Blue connections represent forward and backward connections. Green and red forward and backward connections represent long-range cortical connections. DCM, dynamic causal modeling; IFG, inferior frontal gyrus; IPL, inferior parietal lobule; OT, occipitotemporal junction.
DCM results
Our results indicated that the highest model evidence was present for a fully connected dorsal and ventral cortical model (Fig. 4), in which (1) phonologically facilitated conditions were denoted by a long-range connection from IFG to OT with a self-inhibiting (intrinsic) connection over the IPL and (2) semantically facilitated conditions were plausibly evidenced by an intrinsic connection over the IFG and no long-range connections.
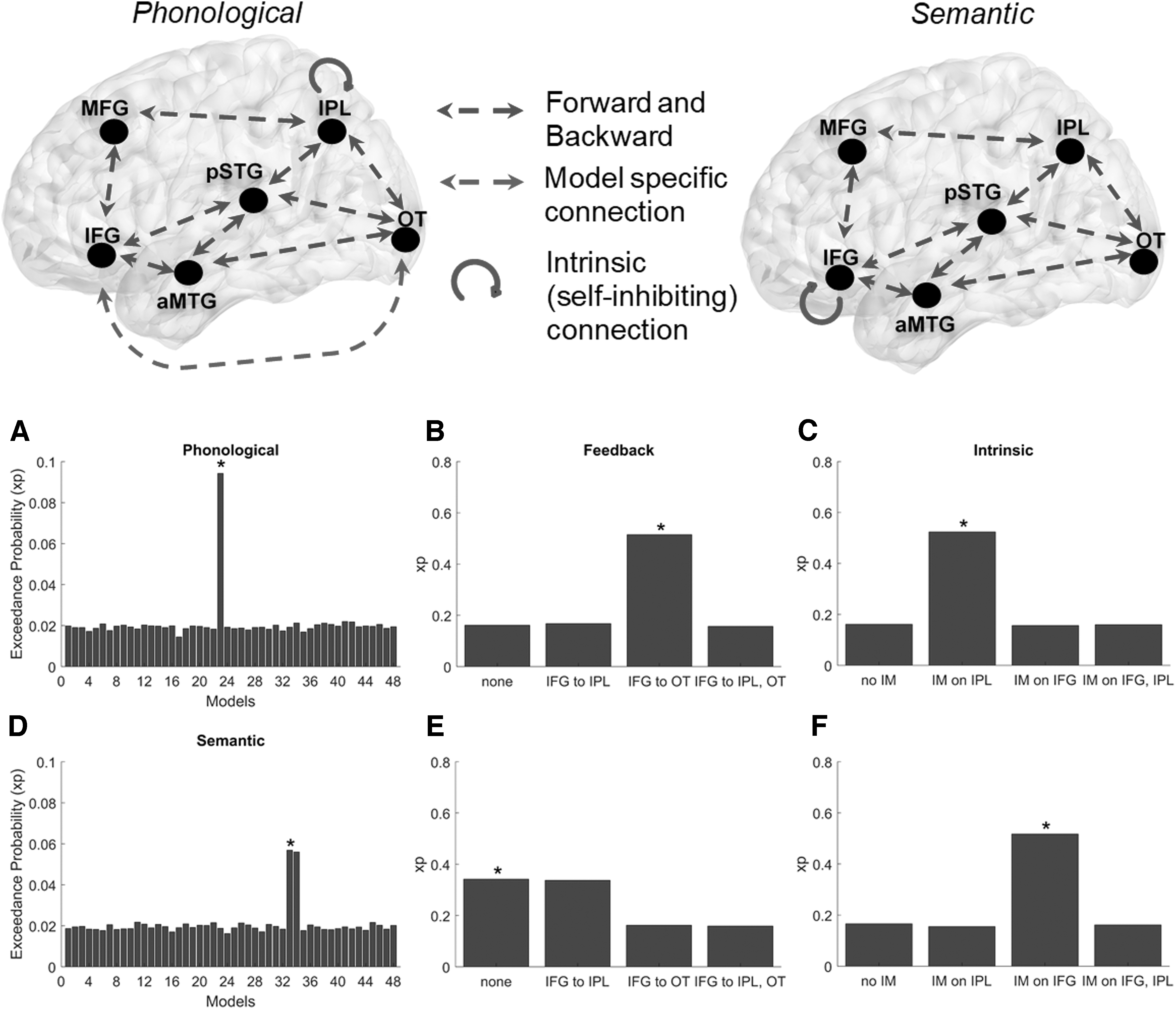
Winning DCM models (asterisked) based on Bayesian statistical inferences, in which model exceedance probabilities indicate that during facilitated learning, phonological models consist of
Discussion
Findings from the present study suggest that during short-term naming facilitation, the effective connectivity of cortical connections, as revealed by DCM analysis, is modulated by common cortical pathways comprising the anterior and posterior temporal gyrus, inferior and MFG, IPL, and occipitotemporal areas. During phonological facilitation, this combined dorsal and ventral cortical model included a long-range, modulatory feedback connection between the occipitotemporal region and IFG, with a self-inhibiting connection over the IPL. This model slightly differed during semantic facilitation, where the most plausible model (based on highest model exceedance probability) consisted only of a self-inhibiting connection over the IFG within a combined dorsal and ventral cortical model. These results provide insights into key cortical connectivity pathways involved in naming-based ERP paradigms, and the potential for monitoring key language mediating regions in older, healthy and/or language-impaired cohorts.
Similar to Iyer et al. (2020), the results indicated that a cortical model with fully connected (forward and backward) dorsal and ventral network pathways explained the results for both semantic and phonological facilitation, though with an additional ROI in the MFG. This fits with recent DCM work showing a crucial role for the MFG in both picture naming (Meier et al., 2016) and semantic judgement (Meier et al., 2018) in healthy older adults, possibly related to its role in domain general cognitive control as opposed to a language-specific contribution.
Crucially, there were also distinct differences in connectivity associated with each form of facilitation. During phonological facilitation, the winning model had the presence of a strong modulatory effect between the occipitotemporal region and IFG and a self-inhibiting connection over the IPL, a model quite similar to that observed for the controls in Iyer et al. (2020). The results are, at least partially, consistent with Heath et al. (2012a) who found that short term phonological facilitation was associated with a repetition suppression effect within the inferior occipito-temporal region, suggesting that these short term facilitation effects are driven by semantic and object recognition processes. Van Turennout et al. (2003) also found that repeated naming of objects led to decreased activity within occipital-temporal and inferior frontal regions. Thus, the modulation between the IFG and OT region observed in the present study suggests the likely presence of strong, top-down modulatory processes occurring in response to phonological facilitation.
The self-inhibitory connection with the IPL for the winning model of phonological facilitation may also reflect its involvement in phonological operations. The IPL is often activated during word production (Indefrey and Levelt, 2004) although its precise role is uncertain (Indefrey, 2011). It has been argued that the IPL serves as a phonological store (Baddeley, 2003) with Vigneau et al. (2006) suggesting its involvement in a fronto-parietal loop for phonological working memory. Accordingly, intrinsic IPL connections may represent bottom-up support for word production and phonological code retrieval stemming from hearing and/or producing the spoken word form during facilitation.
While there was a clear winning model for phonological facilitation, the results were less clear for semantic facilitation. In particular, while the winning model contained a strong self-inhibiting connection over the IFG, the expected probability of this model (model 33) was close to that of a model with an additional connection from the IPL to IFG (model 34). Heath et al. (2012b) found that short-term semantic facilitation effects were associated with repetition enhancement effects in the left inferior occipital gyrus (extending to the left fusiform gyrus), suggesting that such facilitation effects may be driven by episodic memory and visual object recognition enhancement effects. While the current results are not incompatible with that view, the self-inhibiting connection within the IFG points to the possibility that semantic information is exerting top-down control, which is consistent with IFG involvement in controlled semantic processing (Binder et al., 2009; Klaus and Hartwigsen, 2019).
Of note, Iyer et al. (2020) found that improvements in naming following semantic treatment in patients with chronic aphasia were not predicted solely by ventral stream components. Specifically, post-therapy outcomes from semantic naming therapy covaried with pretherapy effective connectivity between IPL and IFG. What is particularly interesting in this case is that while the naming treatment delivered to patients in Iyer et al. still necessitated overt production of the target name during treatment, the facilitation task utilized in the present study avoided any production of the target. Notwithstanding the obvious differences in population and study design, the present results are still somewhat consistent with prior studies in that they suggest that the effects of semantic facilitation techniques may be mediated, at least in part, by changes to engagement of both dorsal and ventral pathways.
Finally, there are a number of limitations that should be considered. First, the present study only examined the short-term facilitation effects that spanned a few minutes in length, and was conducted exclusively on healthy adults. Similar to previous fMRI research, further work should consider the comparison of short-term versus longer term effects of facilitation (Heath et al., 2012b, 2012c), as well as investigating how these facilitation techniques compare in people with aphasia who have different loci of naming difficulties (Heath et al., 2012c, 2013). While the current study focused upon older adults as a precursor to future work in people with aphasia, recent findings of functional connectivity differences between younger and older adults during overt picture naming (Ferré et al., 2020) suggest that analyses of how age impacts the neural response to naming facilitation are also warranted. Another limitation of our study was that although high-density EEG greatly improves the spatial resolution of cortical activity, the precise location of cortical nodes studied within this experiment is likely to vary for fMRI and/or MEG. It would be advantageous for future studies to cross-examine concurrent and/or sequential fMRI/EEG naming paradigms to gauge the spatial accuracy of cortical regions studied. Lastly, it is acknowledged that the current study utilized a delayed rather than an immediate naming paradigm, a decision designed to eliminate the impact of overt articulatory processes on the EEG signal.
Conclusions
The present study provides strong support for a common network comprising cortical pathways that traverse dorsal and ventral pathways during phonological and semantic naming facilitation. The key differences observed across this common network between these tasks highlight that the frontal gyrus plays an important role in mediating top-down feedback to lower order processes. The current findings could be repurposed for future investigation of naming facilitation in people with poststroke aphasia and factors that predict optimal responsiveness to such techniques.
Footnotes
Authors' Contributions
All authors contributed to the study's conception, experimental design, and data analysis. A.J.A. and D.A.C. contributed to the design and execution of the short-term facilitation naming paradigm. K.K.I. designed and implemented the source activation and DCM analysis. All authors were involved in evaluating the study's results, write-up, and drafting of the article. All authors read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was conducted with support from the ARC Centre of Excellence for the Dynamics of Language (Project ID: CE140100041). D.A.C. was supported by a University of Queensland Vice-Chancellor's Fellowship.