
Abstract
Introduction:
Impulse control disorders (ICDs) represent a side effect of dopaminergic medication in Parkinson's disease (PD). Patients experience an excessive desire toward natural rewards paired with uncontrolled actions. Yet, the precise neural and behavioral mechanisms associated with ICDs and, importantly, each specific subdomain remain unclear. We aim to decipher resting-state and corticolimbic functional connectivity in PD patients with and without hypersexual ICD.
Materials and Methods:
Seventeen PD patients with hypersexuality (PD+HS) and 15 PD patients without hypersexuality (PD-HS) underwent two sessions (with and without medication) of resting-state functional magnetic resonance imaging and were compared with 17 healthy controls. Dual-regression independent component analyses extracted salience, sensorimotor, default-mode, and central executive networks. Seed-based functional connectivity with three striatal subdivisions (motor, associative, and limbic) was obtained and significant changes were correlated with key impulsivity and inhibitory measures.
Results:
Enhanced salience network (SN) activity represented by a significant rise in the right inferior frontal gyrus was found in PD+HS compared with PD-HS. Connectivity analyses revealed a functional disconnection between associative and limbic striatum with precuneus and superior parietal lobe in PD+HS, some connections explained by abnormal sexual behavior and inhibition in PD+HS.
Conclusions:
Hypersexual ICD is associated with enhanced SN signaling and corticolimbic disconnections, including striatal associative and limbic loops that contribute to altered control of sexually driven behavior and overall severity in PD and ICD.
Impact statement
We enlarge the neurobiological basis to one specific Parkinson's disease (PD) and impulse control disorder (ICD) (PD+ICD) subtype—that is, hypersexuality—and reveal its associated resting-state functional connectivity linked to altered behavior. We report enhanced salience network and right inferior frontal gyrus as part of the underlying resting-state functional networks in PD patients with hypersexuality (PD+HS). Corticolimbic changes were associated with sexual severity in PD+HS to hypoactive connectivity between associative-limbic striatum with precuneus and superior parietal lobe. The connectivity changes seen in PD+HS could explain baseline differences that engender aberrant control over sexual behavior in ICD.
Introduction
The mesolimbic system regulates the human tendency to approach natural stimuli with positive connotations and to avoid options that entail negative or harmful outcomes (Salamone and Correa, 2012). Dopamine and reward operate to maximize the best outcome for the individual by favoring positive actions and preventing negative ones. This equilibrium requires motivational control to select the best possible output and relies on a large brain network involving the orbitofrontal cortex (OFC), anterior cingulate cortex (ACC), ventral striatum, or the ventral tegmental area (VTA) among others (Haber and Knutson, 2010; Nieoullon and Coquerel, 2003).
In Parkinson's disease (PD), dopamine replacement therapies improve parkinsonism but relatively often induce abnormal responses such as dyskinesias and impulse control disorders (ICDs). The latter is expressed by excessive and compulsive overuse of reward items such as food, sex, hobbies, shopping, or gambling (Weintraub et al., 2010). A hyperdopaminergic state is caused in ICD patients along the mesolimbic pathway that directly alters the reward system, through which desire toward natural rewards increases and uncontrolled behaviors are observed (Weintraub and Claassen, 2017).
Patients with PD and ICD (PD+ICD) exhibit greater tendency for risk-taking behavior (Navalpotro-Gomez et al., 2020; Voon et al., 2011), increased subjective value of reward (Girard et al., 2019), higher novelty seeking (Djamshidian et al., 2011), low social cognition functions including poor empathy (Rossi et al., 2010), as well as poor learning from negative outcomes (Djamshidian et al., 2010; Martini et al., 2018). However, cognitive control functions such as flexibility (set shifting, verbal fluency, and attention) or in response inhibition, PD+ICD patients are relatively spared compared with PD patients without ICD (PD-ICD) (Claassen et al., 2015; Leroi et al., 2013; Santangelo et al., 2017), although contrasting evidence suggests that PD+ICD patients exhibit problems in the acquisition of stopping abilities (Leplow et al., 2017). Hence, the motivational control over natural rewards in PD+ICD is associated with certain mesolimbic functions that may guide problems in learning from negative outcomes and controlling behavior adequately.
The neural substrate part of PD+ICD shows changes in brain-oxygen-level-dependent (BOLD) activity along the mesocorticolimbic circuit. Several functional neuroimaging studies reported decreased activity along the OFC, ACC (Imperiale et al., 2018; Voon et al., 2011), and in the ventral striatum (Girard et al., 2019; Navalpotro-Gomez et al., 2020; Rao et al., 2010; Voon et al., 2011) during risky decision-making tasks while ON-medication. The motivational upstate part of PD+ICD pathophysiology is constrained to increased activity in the ventral striatum and OFC during reward outcome, suggested to represent a “better than expected” result as part of their continuous and compulsive search for positive reinforcers (Frosini et al., 2010; Girard et al., 2019; Stark et al., 2018; Voon et al., 2010). It is noteworthy that task-related BOLD activity underestimates the ongoing baseline activity in PD+ICD patients, a possible mechanism that could explain changes described above along the mesocorticolimbic circuits.
Functional connectivity using resting-state functional magnetic resonance imaging (rs-fMRI) has been informative in delineating baseline activity in PD+ICD. One study reported that PD+ICD patients exhibit increased connectivity within the salience network (SN) and default mode network (DMN), while connectivity within the central executive network (CEN) is reduced (Tessitore et al., 2017b). Using seed-based connectivity, reduced connectivity between the cortico/subcortical hubs and weaker frontostriatal connectivity are often reported in PD+ICD patients (Carriere et al., 2015; Petersen et al., 2018; Ruitenberg et al., 2018). A recent study using dynamic rs-fMRI showed that PD+ICD patients exhibit difficulties in switching between different brain networks and engage more frequently in brain states associated with SN activity (Navalpotro-Gomez et al., 2020).
The above studies using rs-fMRI have enrolled PD+ICD with mixed ICD subtypes and correlate general behavioral measures not directly related to each ICD subtype. As the neural circuitries for each reward type (sexual, food, money) differ (Sescousse et al., 2013), we hypothesize that different ICD subtypes might result in varying patterns of functional connectivity and associated behavioral dysregulation. We aim to decipher the neural state at rest in one specific PD+ICD subtype (i.e., hypersexuality) and the associated functional connectivity changes correlated with behavioral measures of sexual compulsivity and inhibition. To do so, we compare distinct brain networks (sensorimotor, SN, DMN, CEN), striatal seed-based connectivity based on its functional properties (sensorimotor, associative, and limbic), and extrastriatal regions including substantia nigra, VTA, subthalamic nucleus (STN), and amygdala to correlate with cognitive control (erotic stop signal task) and behavioral variables across medication and groups.
Materials and Methods
Participants
Forty-nine heterosexual male and cognitively preserved participants were enrolled. Hypersexuality is one of the most common problems involved in PD+ICD, with greater prevalence among men (Weintraub and Claassen, 2017). We recruited 17 PD patients with hypersexuality (PD+HS), 15 PD patients without hypersexuality (PD-HS), and 17 age-matched healthy controls. Patients met the U.K. Brain Bank diagnostic criteria for PD (Hughes et al., 1992). Because of the special characteristics of the sample, four hospitals located in Madrid's metropolitan area participated in the recruitment (HM CINAC - HM Puerta del Sur, Hospital Fundación Alcorcón, Hospital Clínico San Carlos, and Hospital 12 de Octubre). After referral from a neurologist, a second interview was conducted with the neuropsychologist (patient and family) to confirm the presence of hypersexual state and heterosexuality. Finally, PD+ICD was quantified using the Questionnaire for Impulsive-Compulsive Disorder in Parkinson's Disease (QUIP) and patients with scores ≥5 on the sexual subscale and clinical confirmation during interview joined the PD+HS group. Exclusion criteria were established based on the presence of (1) cognitive dysfunction as defined by a score of ≤24 on the Montreal cognitive assessment (MoCa), (2) depression (>18 in Beck depression inventory [BDI]), or (3) other severe comorbidities (e.g., history of substance abuse, hallucinations, or psychosis detected during the interview). The study was approved by the ethics committee (CEIm HM hospitales: 17.03.0852E2-GHM) and signed consents from every participant were obtained before their enrollment in the study.
Data acquisition and preprocessing
All subjects underwent MRI scans in ON- and OFF-medication conditions. MRI scans were performed in a 3T scanner (MMR Biograph, Siemens, Erlangen, Germany), with the following acquisition protocol: three-dimensional T1-weighted magnetization prepared-rapid gradient echo image with parameters TR/TE/TI 2300/3.34/900 ms, flip angle 8°, and isotropic spatial resolution 1 mm3 (FoV: 256 mm, matrix: 256 × 256, slice thickness: 1 mm); rs-fMRI using a single-shot gradient-echo-planar imaging (EPI) two-dimensional (2D) pulse sequence, with acquisition parameters TR/TE 2000/30 ms, flip angle 77°, and spatial resolution 3 × 3 × 4 mm3 (FoV: 192 mm, matrix: 64 × 64, slice thickness: 4 mm), the acquisition of this sequence lasted 10 min giving rise to 300 fMRI volumes. A field map was generated from two 2D gradient echo images, with acquisition parameters TR/TE1/TE2 455/4.92/7.38 ms, flip angle 60°, and the same spatial resolution as the fMRI acquisition. During fMRI acquisition, subjects were instructed to remain calm, motionless, without engaging in any cognitive or mental task, and to remain eyes-open with their eyes fixated on a white cross (black background).
T1-weighted MRI images were corrected for intensity bias (Tustison et al., 2010), denoised using an optimized nonlocal means filter (Coupe et al., 2008), skull-stripped using brain extraction tool-FMRIB software library (Smith, 2002), and segmented in white matter, gray matter, and cerebrospinal fluid (CSF) tissues using FMRIB's automated segmentation tool (Zhang et al., 2001). Skull-stripped images were nonlinearly registered to the MNI152 (1 mm3) space using an affine transformation followed by diffeomorphic symmetric normalization in advanced normalization tools (ANTs) (Avants et al., 2008). White matter and CSF masks were eroded to avoid partial volume contamination. Eroded CSF mask was confined to the ventricles using the mask provided in FSL.
During fMRI, preprocessing tools from analysis of functional neuroImages and FSL packages were used. The first five volumes of the rs-fMRI scan were discarded to allow signal stabilization. Then the fMRI series were despiked, corrected for slice-timing, realigned to the first volume, skull-stripped, and corrected for EPI distortion using the field map. A tagging process was then carried out (Power et al., 2012) taking three data-quality metrics: frame-wise displacement (FD), the derivative of the root mean square function across voxels (spatial root mean square of the data after temporal derivative [DVARS]), and the standard deviation (SD). Both DVARS and SD were computed for motion-corrected images using a gray matter mask. rs-fMRI volumes overpassing a conservative value of 0.3 for FD or a cutoff of 75%-percentile +1.5 × interquartile-range for DVARS and SD was tagged as noisy volumes. Although the noisy samples were not removed from the series during preprocessing, they were not considered for the estimation of functional connectivity (Power et al., 2012, 2014). Sessions with more the 30% of the volumes tagged as corrupted were discarded.
Motion induced artifacts were automatically removed using independent component analysis-based strategy for Automatic Removal of Motion Artifacts (ICA-AROMA) (Pruim et al., 2015). Noisy components identified with ICA-AROMA were removed from nonsmoothed fMRI using fsl_regfilt. The mean of the rs-fMRI series was subtracted, and linear trends were removed. A linear affine transformation was obtained between the average rs-fMRI volume and the T1-weighted image using ANTs. The inverse of this transformation was applied to the white matter and CSF masks to extract tissue average rs-fMRI activity. The nuisance regressor matrix contained the tissue average time series and their derivatives. The four regressors were z-scored and were regressed-out from the data set. The resulting residuals were band-pass filtered (0.009–0.1 Hz) using a second-order Butterworth filter. Preprocessed rs-fMRI was normalized to MNI152 (3 × 3 × 3 mm3) and smoothed using a 6-mm full-width-at-half-maximum Gaussian kernel.
Striatal seed-based functional connectivity analysis
Functional connectivity maps were computed as the z-transformed Pearson linear correlation between the average resting fMRI time-series of each region of interest (ROI) and all voxels in the whole brain. ROIs included were selected based on abnormal dopaminergic state in ICD literature along the striatal axes as well as those regions part of the mesolimbic route. We included striatal subdivisions from the striatal connectivity atlas from sensorimotor, associative, and limbic subdivisions (Tziortzi et al., 2014). The substantia nigra and STN were obtained from the 7T MRI atlas (Keuken et al., 2014); the amygdala from the Harvard/Oxford subcortical atlas; and the VTA parcellation by Murty and associates (2014). A total of 12 connectivity maps were obtained for each subject. Group functional connectivity maps for all ROIs were separately obtained by running a random-effect second-level analysis in SPM12, using a one-sample t-test. Statistical maps showing group connectivity were thresholded at the voxel level (p < 0.05, family-wise error [FWE] corrected for multiple comparisons). An implicit mask obtained from activations from the control group as per each network was used in comparisons between patients. The main effects for drug (ON- vs. OFF-medication) or disease (PD-HS vs. PD+HS) were tested within a two-way analysis of variance (ANOVA). Significance was considered for an uncorrected p-value of 0.001, with FWE correction at the cluster level (P FWE < 0.05).
Dual-regression analysis
Group-level resting-state networks were estimated using a group ICA with MELODIC and taking the preprocessed images (before nuisance regression and unfiltered) from all participants. The number of independent components was set to 15. Dual-regression was run in four resting-state networks of interest—that is, the sensorimotor, SN, DMN, and CEN—to extract individual-level functional networks, and the z-score map of each network was obtained. The main effects for drug (ON- vs. OFF-medication) or disease (PD-HS vs. PD+HS) were tested in a two-way ANOVA. Significance was considered for an uncorrected p-value of 0.001, with FWE correction at the cluster level (P FWE < 0.05).
Neuropsychological and behavioral assessments
Neuropsychological and behavioral tests were used as cognitive markers of brain connectivity changes as measured with Pearson correlation coefficients. Assessments were done ON-medication state before the scanning sessions. The tests included a general cognitive state using MoCa (Nasreddine et al., 2005), cognitive control using the Stroop interference test (word reading, color naming, and word-color interference) (Stroop, 1935), and executive screening using the frontal assessment battery (Dubois et al., 2000).
Behavioral measures of response inhibition were included. A modified stop-signal task based on the classic version developed by Logan and Cowan (1984) was used using visual erotic and nonerotic stimuli (erotic stop signal task; adapted from Prévost et al., 2010). Images were included and rated (valence and arousal levels per image) by independent male participants (n = 18) in a pre-experiment condition to determine a final database. The task provides accurate measures of the quality of response inhibition under the influence of highly appetitive erotic images and compared with nonerotic images. In brief, each trial started with a fixation cross (1000 ms), followed by an erotic or nonerotic stimulus (1000 ms). Subsequently, a go signal—a white arrow pointing either left or right—prompted participants to respond as fast and as accurately as possible with their index (for left arrows) or middle finger (for right arrows). On some trials, a white stop signal appeared immediately after the go signal in which case participants should withhold their already initiated response. The delay between the go and stop signal followed a staircase procedure termed stop signal delay (SSD; four independent SSD starting at 50, 100, 150, or 200 ms). The staircase adjusted for each trial based on accuracy of the previous stop signal: if the subject failed/succeeded at a given stop trial, the SSD would decrease/increase, respectively, one staircase for the next stop trial. A total of 192 erotic and 192 nonerotic trials were performed, including 128 go and 64 stop trials. The calculation of stopping ability was based on the stop signal reaction time (SSRT) using the integration method (iSSRT; Verbruggen et al., 2019).
Results
Table 1 provides results on the clinical, sociodemographic, and neuropsychological variables between groups. A total sample size of 11 PD+HS, 13 PD-HS, and 13 controls were included in the final analysis as some patients were discarded due to the excessive number of tagged images (>30%) as motion corrupted. No significant differences were observed between patient groups in terms of clinical condition (Unified Parkinson's Disease Rating Scale [UPDRS], levodopa-equivalent daily dose [LEDD], or disease duration), demographic variables (education or age), or neuropsychological assessments (Table 1). The presence of other PD+ICD subtypes was detected although the sexual subscale dominated for every patient. However, in the behavioral erotic stop signal task, worse performance was seen in PD-HS compared with PD+HS while ON-medication [t (27) = −3.25, p = 0.003] in the erotic condition and in the nonerotic condition [t (27) = −3.72, p < 0.001] of the iSSRT.
Sociodemographic, Clinical, and Behavioral Variables
Unpaired t-test is displayed between PD+HS and PD-HS.
(i)SSRT, (integrative) stop-signal reaction time; BAI, Beck's Anxiety Inventory; FAB, frontal assessment battery; GDS, geriatric depression scale; LEDD-DA, levodopa equivalent daily dose of dopamine agonist; LEDD-total, levodopa equivalent daily dose (total mg); MoCa, Montreal cognitive assessment; n.s., not statistically significant; PD, Parkinson's disease; PD+HS, PD patients with hypersexuality; PD-HS, PD patients without hypersexuality; QUIP, Questionnaire for Impulsive-Compulsive Disorder in Parkinson's disease; RT, reaction time; SAS, Starkstein Apathy Scale; UPDRS, unified Parkinson's disease rating scale (ON/OFF-medication total score).
Networks activated by hypersexual ICD
Dual-regression ICA showed a significant disease effect within the SN (Fig. 1), through which the PD+HS group showed significant hyperconnectivity of the right inferior frontal gyrus (IFG) (40, 20, 17; p < 0.005; Fig. 1) compared with PD-HS. Moreover, SN connectivity was lower for the PD-HS group compared with healthy controls. No significant interactions were found among the remaining networks of interest, including sensorimotor, DMN, or CEN, across groups and medication effects.
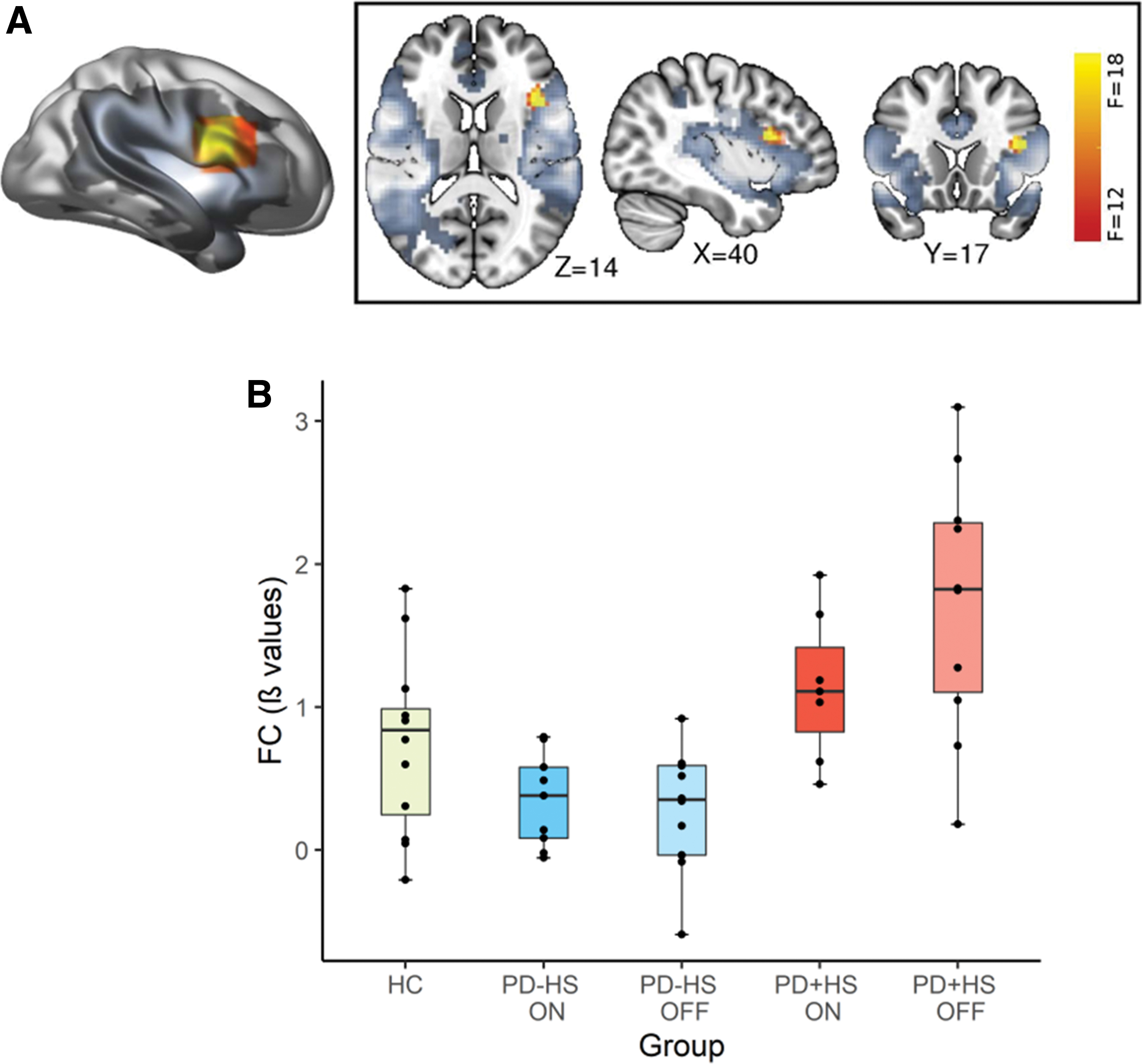
SN functional connectivity to IFG.
Striatal seed-based functional connectivity: disease and medication effects
Several significant drug- and disease-main effects at several striatal locations were found (Tables 2 and 3 and Fig. 2). Unless otherwise stated, all drug-main effects were driven by the ON-medication state, showing decreased connectivity compared with the OFF-medication state.
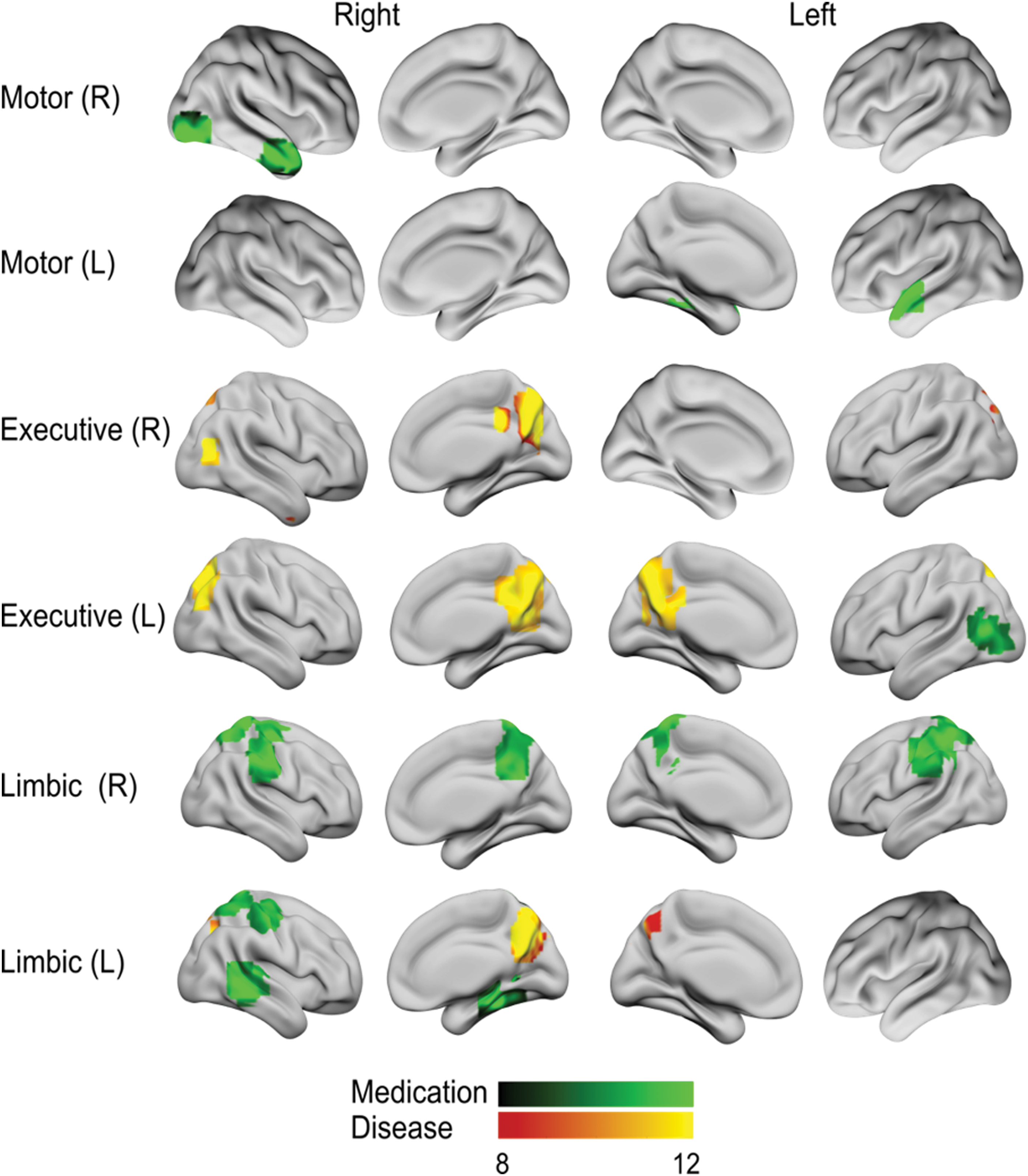
Striatal seed-based functional connectivity. Differences in functional connectivity maps for different striatal seeds in both hemispheres are shown in green-black for medication effects, while red-yellow tones indicate disease-group effects (p < 0.001; FWE correction cluster level). Color images are available online.
Striatal Seed-Based Functional Connectivity: Disease Effects
Post hoc comparisons between PD+HS and PD-HS (paired t-test) medication states shown at a significance level of p < 0.05. Secondary effect comparisons between ON and OFF (unpaired t-test) at a significance level of p < 0.05.
MNI, Montreal Neurological Institute.
Striatal Seed-Based Functional Connectivity: Medication Effects
Post hoc comparisons between ON and OFF (paired t-test) medication states shown at a significance level of p < 0.05. Secondary effect comparisons between PD+HS and PD-HS (unpaired t-test) at a significance level of p < 0.05.
Disease effects
Following our central hypothesis testing for baseline group effects, hypoactivation in PD+HS patients was seen between the left associative striatum and right precuneus extending to the posterior cingulate, as well as with the right superior parietal lobe (Table 2 and Figs. 2 and 3). Similarly, the right associative striatum showed a significant disease effect with the right precuneus (Table 2 and Figs. 2 and 3). In addition, a disease-effect was found between the left limbic striatum and the right precuneus, with decreased values in the PD+HS group (Table 2 and Figs. 2 and 3). Overall, PD+HS showed reduced connectivity between striatal seeds and cortical hubs representing a possible explanation to hypersexual excessive behaviors.
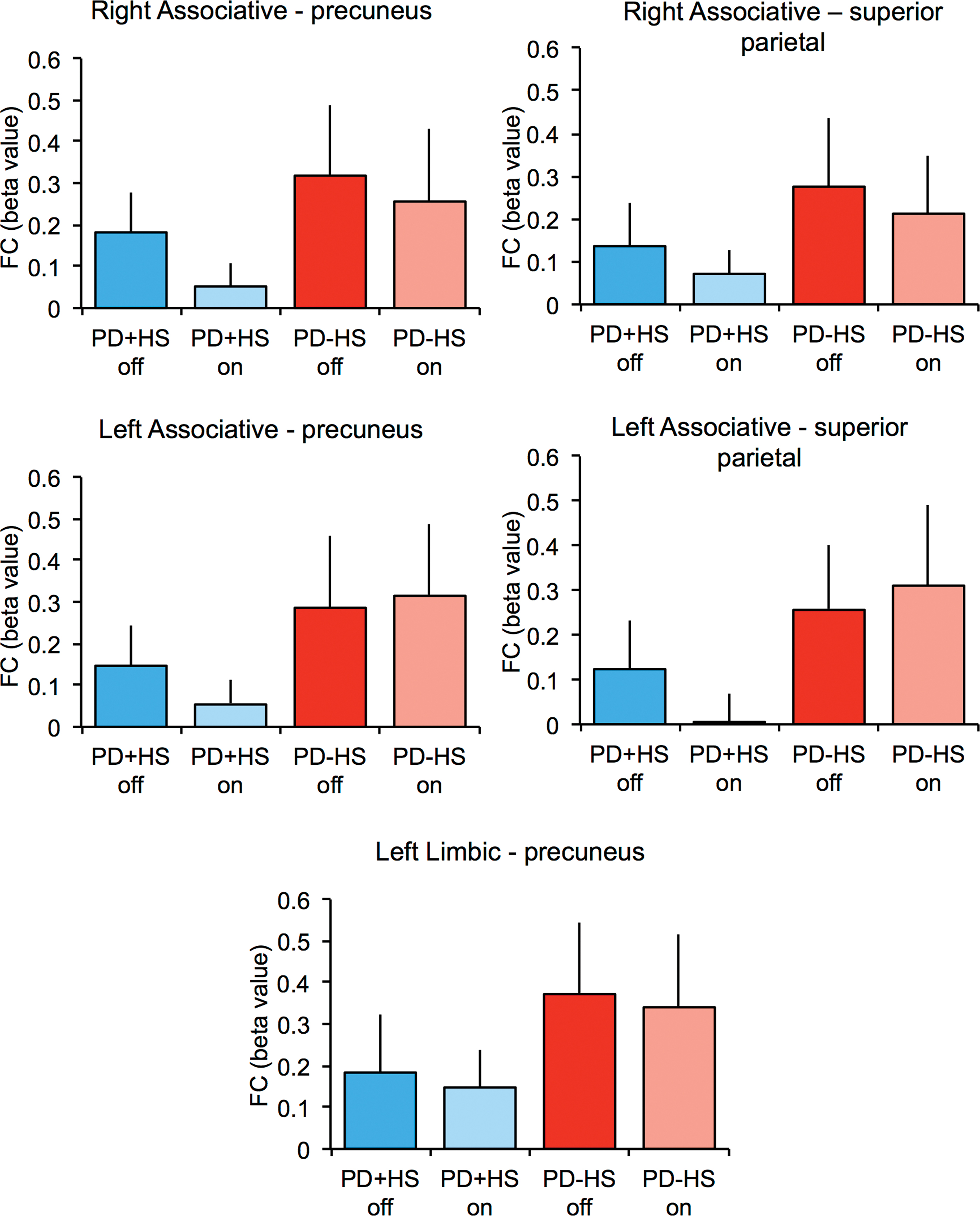
Functional connectivity beta values between several striatal and cortical regions. Functional connectivity beta values are shown for both patient groups in ON- and OFF-medication state. Color images are available online.
Medication effects
The right motor striatum seed showed a significant drug-effect in regions of the temporal lobe, including the temporal pole and the right lateral occipital cortex, whereas the left motor striatum only showed such effect on the left temporal pole (Table 3 and Fig. 2). In addition, a significant drug-effect was found between the left associative striatum and the left inferior lateral occipital cortex (Table 3 and Fig. 2). Moreover, the functional connectivity with the limbic striatum seed was influenced by drug-effects connecting with the bilateral postcentral gyri, the right precentral gyrus, and the right precentral gyrus (Table 3 and Fig. 2)—for this effect, hypoconnectivity was especially marked in PD-HS compared with the PD+HS (ON-medication), indicating that medication in PD-HS induces a reduction in the limbic striatum to precentral gyrus connectivity and the right superior parietal lobule when the seed was placed in the right hemisphere. Similarly, for the left limbic striatum, increased functional connectivity was observed with the following areas of the right hemisphere: the postcentral, middle temporal gyri, and parahippocampal gyri, as well as with the precuneus cortex (Table 3 and Fig. 2).
Extrastriatal seed-based functional connectivity: medication effects
To ascertain the mesolimbic contribution to networks associated with hypersexual ICD, seed-based connectivity was also conducted at several extrastriatal locations (substantia nigra, VTA, STN, and amygdala; Table 4). None of the reported effects are significant at the group level and medication effects were all showing reduced connectivity while ON-medication across patients groups, consistent with prior evidence (Kwak et al., 2012; Yang et al., 2016). The right substantia nigra showed significant drug-effects characterized by decreased activation in the right and left superior temporal gyrus, right IFG, left parahippocampal gyrus, and supplementary motor area for the ON-medication state compared with OFF-medication (Fig. 4 and Table 4). Moreover, the left substantia nigra showed significant drug-effect connectivity with bilateral supramarginal cortices resulting in decreased ON-medication state activity. Similarly, the right STN showed significant drug-effects showing hypoactivation (ON vs. OFF) with the parahippocampal gyrus, temporal pole, motor cortex, and the anterior part of the superior temporal gyrus (Fig. 4 and Table 4). The left VTA showed a significant drug-effect connecting to right superior parietal area the right precentral gyrus, an effect present while ON-medication for both groups (Fig. 4 and Table 4). Finally, significant drug effects in the ON-medication group were found between the left amygdala and the right precuneus—with stronger hypoconnectivity for the PD+HS group compared with the PD-HS group (Table 4)—as well as with the right supramarginal gyrus.
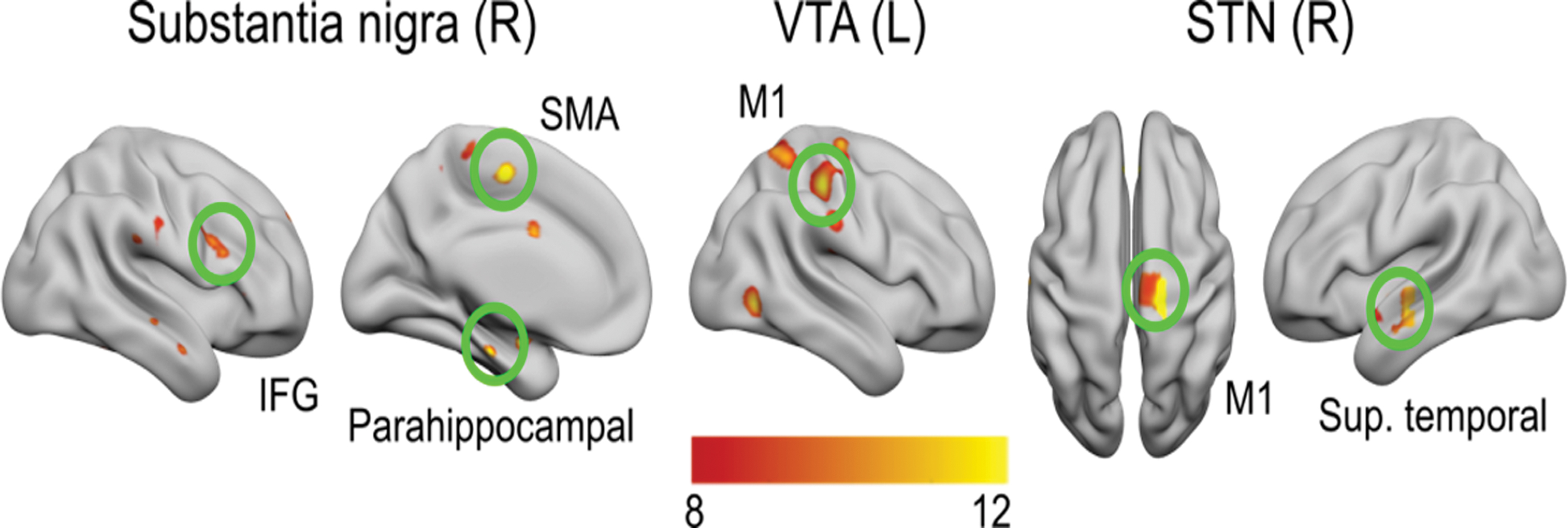
Seed-based functional connectivity from substantia nigra, VTA, and STN. Right substantia nigra connections to right superior temporal pole and left supplementary motor area; left VTA functional connectivity with right premotor cortex and right superior parietal gyrus; right STN connections with motor cortex and superior temporal lobe. STN, subthalamic nucleus; VTA, ventral tegmental area. Color images are available online.
Extrastriatal Seed-Based Functional Connectivity: Effect of Medication
Post hoc comparisons between ON and OFF (paired t-test) medication states shown at a significance level of p < 0.05. Secondary effect comparisons between PD+HS and PD-HS (unpaired t-test) at a significance level of p < 0.05.
Functional connectivity and behavioral correlates
To explain the group effects, the connectivity between the associative striatum and lateral superior parietal lobe was positively correlated to sexual severity as scored by the QUIP sex in PD+HS (r = 0.81, p = 0.02). Meanwhile, connectivity changes between limbic left and precuneus were positively related to reduced levels of QUIP sex scores in the PD-HS group (r = 0.76, p = 0.01).
Medication-driven correlations (while on medicated) were detected in the limbic striatum and parahippocampal and precentral regions, showing positive correlations with inhibitory control under erotic stimuli in PD-HS (r = 0.78, p = 0.01; r = 0.79, p = 0.01; respectively), but not found in PD+HS (p's > 0.05). Conversely, PD+HS patients showed a strong positive correlation with the erotic SSRT on the right substantia nigra to superior temporal gyrus (r = 0.91; p < 0.01) and the right STN to superior temporal pole connectivities (r = 0.82, p = 0.02). Finally, a double opposite correlation was found between VTA connectivity with superior frontal gyrus and QUIP sex scores, where PD+HS exhibited a positive relationship (r = 0.78, p = 0.03) but a negative one in the PD-HS (r = −0.78, p = 0.01) while medicated.
Discussion
The aim of the present study was to examine the differential patterns of functional connectivity in spontaneous fluctuations of BOLD in PD+HS compared with PD-HS and controls. The novelty of the study is to specifically test functional resting-state and mesocorticolimbic seed-based connectivity in one ICD subtype, that is, hypersexuality. Most of the existing resting-state evidence includes mixed cohorts of ICD subtypes in the context of PD (Carriere et al., 2015; Navalpotro-Gomez et al., 2020; Ruitenberg et al., 2018; Tessitore et al., 2017a,b). Given that different reward systems rely upon separable regions (Sescousse et al., 2013), our approach allowed increased control over potential confounders such as cue-dependent rewards and associated reward circuits that may emerge in heterogeneous PD+ICD samples.
Enhanced SN and right IFG in PD+HS
The prevalent function of the SN is to integrate sensory, cognitive, and emotional stimuli that are relevant to the individual as well as to switch between network states (Menon and Uddin, 2010). Specifically, the SN is dedicated to attention orienting toward rewarding cues by engaging cortical and subcortical hubs, including the anterior insula, the IFG, the ACC, substantia nigra, and ventral striatum (Menon and Uddin, 2010). We show specific enhancement in the SN in PD+HS with maximal representation over the right IFG compared with PD-HS. Since the SN monitors attention orienting toward rewarding cues (Anderson, 2017; Bucker and Theeuwes, 2014), our results may play a central role in the understanding of enhanced sexual desire in PD+HS, given patients enhanced cue detection system toward sexual cues.
The striatum and the SN diminish functional coupling as disease progresses in PD (Putcha et al., 2016), with impact over key nodes in the SN that become aberrant upon dopaminergic treatment in PD+ICD, such as the IFG (Ruitenberg et al., 2018). The right IFG is a key hub of the SN and CEN with established roles in inhibitory control (Aron et al., 2014; Maizey et al., 2020; Rae et al., 2015) and attentional orienting to relevant stop-related stimuli (Boehler et al., 2011; Hampshire et al., 2010; Sharp et al., 2010). Importantly, recent evidence pinpoints the IFG as essential for successfully inhibiting behavior upon the presence of erotic stimuli (Rodriguez-Nieto et al., 2019). Interestingly, our PD+HS patients outperformed in response inhibition ability (both under erotic and nonerotic influence) compared with PD-HS as reported elsewhere using a classic stop signal task (Claassen et al., 2015). Since our PD+HS sample showed good levels of response inhibition, a function associated with the IFG, the increased functional connectivity between the SN and the IFG could be rather associated with motivational mechanisms such as increased attention to reward-related stimuli (i.e., sexual cues). We suggest that the enhanced baseline activity of the SN and IFG in PD+HS is directly associated with excessive search of sexual cues as a consequence of their enhanced motivational state.
Connectivity fMRI studies (with mixed cohorts of PD+ICD subtypes) likewise report enhanced SN effects and general heightened functional connectivity compared with PD-ICD patients (Navalpotro-Gomez et al., 2020; Ruitenberg et al., 2018; Tessitore et al., 2017a,b). The analogous results reported by previous studies may be driven by the hypersexual weight of their sample, present in nearly half of their patients (Navalpotro-Gomez et al., 2020; Tessitore et al., 2017a,b). Isolated analyses of PD+ICD subtypes would be informative to provide distinct resting-state network weights as per each behavioral dysregulation. Yet, previous studies do not distinguish the differential contributions from different PD+ICD subtypes on SN-enhanced connectivity. Dividing PD+ICD across subtypes may enhance our understanding of each category as we know the reward system counts on separable neural circuitries for primary (food, sex) versus secondary rewards (money, social) (Sescousse et al., 2013).
Although the DMN did not reveal significant differences between groups, some central regions of the network did show hypoactive functional connectivity with striatal territories. Here, precuneus connectivity with the associative striatum (right and left) and left limbic striatum was hypoactive in the PD+HS group only. The precuneus is functionally connected with striatal areas (Di Martino et al., 2008) and both take part in some behaviors related to impulsivity, social cognition, and reward-based processes (Cox and Witten, 2019). The striatum and precuneus functionally engage in high- versus low-risk decisions, both showing a significantly higher activity for riskier choices (Roy et al., 2011). Other studies have shown that precuneus exhibits increased activation during several behaviors, including episodic memory retrieval (Mazzoni et al., 2019), empathy and perspective taking (Banissy et al., 2012), self-processing operations (Farrer and Frith, 2002), reward outcome monitoring (Neubert et al., 2015), and emotional stimulus processing (Maddock et al., 2003).
While few links have been assigned between precuneus and behavioral manifestations in PD+ICD, some studies do report precuneus connectivity with motor areas as hypoactive (Thibes et al., 2017) and associated with mild cognitive impairment in PD (Jia et al., 2019). Among the principal precuneus roles described above, first-person perspective taking and an experience of agency are unimpaired in PD+ICD (Mack et al., 2013). Nevertheless, PD patients with and without ICD show altered empathy as measured by classical mind-reading tests (Rossi et al., 2010). Hence, the known disconnection occurring between the precuneus and motor areas (cortical and striatal) in PD (Thibes et al., 2017), similar to our PD+HS sample, together with known empathetic problems in both PD and PD+ICD groups (Rossi et al., 2010), makes it plausible to suggest that the precuneus/striatal hypoactive connection may explain PD+HS problems in adjusting social cognition necessary during sexual activities. In fact, the PD-HS group showed greater connectivity between precuneus and striatum correlating with reduced sexual behaviors. Our interpretation goes in line with greater precuneus activation in PD+HS during passive viewing of erotic rewards (Politis et al., 2013) and valuation of costs required to obtain sexual rewards (Girard et al., 2019). Lost connectivity between the striatal areas and precuneus in PD+HS may represent a baseline deficit to generate adequate sexual behaviors.
Corticostriatal and the mesolimbic circuits in PD+HS
The striatum and the brain stem are part of the mesolimbic system and considered two of the most important subcortical hubs in the pathophysiology of PD. We dissected the mesocorticolimbic circuitry using the seed-based approach across three striatal functional territories (motor, associative, and limbic) and the extrastriatal regions (substantia nigra, VTA, STN, and amygdala). The selection of these regions is supported by beliefs that the risk of developing ICD lies on a balance between the preserved versus unpreserved mesocorticolimbic system (Stark et al., 2018). Here, we reveal that PD+HS severity (i.e., QUIP sex scores) correlates with associative striatum-superior parietal hypoconnectivity, a result not seen in the PD-HS group, while different extrastriatal subregions do not explain baseline differences between PD+HS and PD-HS (only expected medication effects).
An extensive network is needed while encoding sexual items, which includes: the ventral striatum, amygdala, insula, ACC, and superior parietal lobe (Georgiadis et al., 2012; Ruesink and Georgiadis, 2017). Meanwhile, volitional inhibition of sexual arousal in healthy men raised neural activity in the superior parietal, IFG, insula, and ventrolateral prefrontal cortex (Beauregard et al., 2001; Rodriguez-Nieto et al., 2019). These observations seem to suggest that sexual inhibition, be it intended or unintended, is related to the exaggerated activity of components of the sexual network (including the amygdala, superior parietal lobule, and ventral sections in the prefrontal cortex). Our result showing a group effect over the superior parietal lobe shows a disconnection with striatum in PD+HS and such disconnection correlates positively with sexual severity in PD+HS. In another study including only PD+HS during passive viewing of erotic stimuli (Politis et al., 2013), the right and left superior parietal lobes, as well as the limbic striatum (among others), were active when specifically viewing erotic versus neutral items. It all seems to indicate that striatal-superior parietal changes upon presentation of ICD can bias baseline functional connectivity and produce greater sexual demands as a by-product of corticostriatal disconnection in PD+HS patients.
Altogether, changes in precuneus and superior parietal functional connectivity could be partly related to DMN alterations that may act in isolation in PD+HS as opposed to the entire network, normally operating as a whole. Considering the established association of DMN with task disengagement, differences in hypersexuality between the functional core of DMN, that is, precuneus, and the broad network involve a new perspective into the possible mechanisms behind connectivity changes in PD+HS. This would represent an inefficient network usage taking “parts of a whole” to modulate baseline resting connectivity that later translates into impulsive acts in PD+HS.
Medication effects and the mesocorticolimbic circuit
The main medication findings of the present study do not contribute to expanding further possible PD+HS baseline mechanisms but to enhance our view of PD-related side effects of treatment. We report an association with PD+HS sexual severity and limbic striatum-parahippocampal hypoconnectivity, a result specific for PD-HS during ON-medication (not observed in the PD-HS group). The parahippocampal region has been linked to sexual approach tendencies in healthy adults (Rodriguez-Nieto et al., 2019), a cognitive control function normally adjusted according to a given personal and social context. This social function is relevant to the parahippocampus, ventral striatum, OFC, and ACC regions in healthy participants (Rodriguez-Nieto et al., 2019; Ruesink and Georgiadis, 2017). Our result gives a special foundation to the striatal and parahippocampal route that medication turns down the connectivity both in PD+HS and PD-HS.
To further understand the relative contribution of motor versus nonmotor ICD origin, we segregated motor-to-nonmotor territories of the midbrain (substantia nigra and VTA). Both structures are often used as separable neural seeds of whole-brain motor and nonmotor networks (Klein et al., 2012; Murty et al., 2014). Here, substantia nigra was significantly connected to the supplementary motor area, IFG, and supramarginal gyrus, an effect driven by medication. Importantly, VTA was connected with the motor cortex, a connection associated with sexual severity (i.e., QUIP sex scores) in the PD+HS group only (ON-medication). Hence, although these effects are not explained by group differences, medication seems to recruit a unified system, including motor and nonmotor loops in PD+HS from limbic to motor cortical areas.
Conclusions
The results reveal the underlying baseline neural connectivity behind one of the major and most problematic PD+ICDs, that is, hypersexuality, with special aberrant signals along salience, mesocorticolimbic, and striatal circuits. In particular, our results disclose aberrant connectivity in circuits, including SN connectivity (comprising the right IFG), together with hypoactive corticolimbic connectivity between the associative-limbic striatum with precuneus and superior parietal lobe in PD+HS. Impaired corticolimbic system may thus account for some neuropsychiatric features as striatal and extrastriatal seed-based connectivity explains some of the abnormal behavioral changes in PD+HS.
Footnotes
Authors' Contributions
D.M.-M: data analysis and article writing; J.A.P.-P.: study design, data collection, data analysis, and article writing; F.A.-F.: patient recruitment; J.A.M: patient recruitment; L.V: patient recruitment; I.O.: patient recruitment, study design, data collection, and article writing.
Acknowledgments
We are grateful to all the participants for their collaboration and the help from imaging technicians for assistance in data collection.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The work was conducted with help from Miguel Servet (CP18/00038), Instituto de Salud Carlos III, Spain (IO), and Fundación HM Hospitales.