
Abstract
Background:
Bilingualism is considered a cognitive reserve (CR) factor, due to the delay in the onset of dementia in bilinguals compared with monolinguals. Two neural mechanisms have been suggested to underlie CR: neural reserve and neural compensation. However, it is still unclear how bilingualism contributes to these mechanisms.
Methods:
In this study, we used cognitive tests, functional connectivity (FC), regional homogeneity, and fractional amplitude of low-frequency fluctuations (fALFF) measures to study resting-state brain patterns in a sample of bilingual and monolingual subjects with mild cognitive impairment.
Results:
We found no significant differences between the groups in age, sex, education, or cognitive level, but bilinguals showed higher FC than monolinguals between the posterior part of the superior temporal gyrus and the precuneus, positively correlated with Mini-Mental State Examination scores, and higher fALFF in the thalamus bilaterally.
Conclusions:
Our results suggest that bilingualism may act as a CR factor that protects against dementia through neural compensation.
Impact statement
Recent investigations suggest that neural compensation is one of the cognitive reserve mechanisms underlying the protection of bilingualism against dementia. Although brain changes in functional connectivity (FC) have been proposed as evidence of this mechanism, no study has directly used FC to study neural compensation in bilingualism. Our findings show that mild cognitive impairment bilinguals manifest higher resting-state FC than monolinguals between the language network and the precuneus, supporting the involvement of neural compensation in the protection of bilingualism against dementia. Moreover, we found bilingualism effects in the spontaneous activity of the thalamus, a region related to atrophy in dementia.
Introduction
Cognitive reserve (CR) refers to the mechanisms underlying the discrepancy between a person's level of brain pathology and his/her cognitive performance, which would be expected to match this pathology. Bilingualism has been proposed as one of the experience-based factors that contribute to CR, based on previous evidence showing that bilinguals exhibit the clinical manifestations of dementia 4–5 years later than monolinguals (Woumans et al., 2015).
To investigate the neural mechanisms of this supposed protective effect of bilingualism against dementia, some neuroimaging research has focused on patients with this condition. Thus, a first study found that bilinguals with the same cognitive level as monolinguals had more brain atrophy indicative of pathology in specific temporal areas that are normally used to distinguish dementia patients from controls (Schweizer et al., 2012).
Similar results have been found in subjects suffering from mild cognitive impairment (MCI) (Costumero et al., 2020; Marin-Marin et al., 2020). In a recent investigation, bilinguals showed reduced parenchymal volume and gray matter (GM) volume in areas related to atrophy in dementia (Costumero et al., 2020). Crucially, this study also found longitudinal differences: during a 7-month follow-up, monolinguals lost more parenchyma and had more cognitive deterioration than bilinguals.
Regarding white matter (WM) disintegration, bilinguals and monolinguals with MCI showed different patterns of atrophy in a diffusion tensor imaging study. Bilinguals showed higher WM integrity in the parahippocampal cingulum and uncinate fasciculus, but lower integrity in the fornix, all of which are fibers associated with language and memory (Marin-Marin et al., 2020).
When investigating the neural basis of CR, two brain mechanisms have been described: neural reserve and neural compensation. On the one hand, neural reserve refers to the efficiency and resilience of preexisting cognitive networks that may be capable of maintaining cognitive function despite brain pathology (Barulli and Stern, 2013; Stern, 2012). Several investigations suggest that this mechanism is related to bilingualism's contribution to CR because healthy older bilinguals show increased GM volume in the anterior cingulate cortex (Abutalebi et al., 2015) and higher WM integrity in the corpus callosum and superior longitudinal fasciculi (Anderson et al., 2018; Luk et al., 2011), compared with monolinguals.
On the other hand, neural compensation occurs when brain networks that are not normally used for a certain cognitive function acquire relevance and compensate for brain alterations in other regions (Barulli and Stern, 2013). Evidence supporting this mechanism in bilinguals comes from studies investigating brain connectivity using neuroimaging techniques (Perani and Abutalebi, 2015). A previous fluorodeoxyglucose-positron emission tomography study in patients with dementia found increased and positive metabolic connectivity in bilinguals compared with monolinguals between the default mode network (DMN) and the executive control network (ECN) and brain areas related to language control, such as the cingulate cortex and the inferior frontal gyrus (Perani et al., 2017).
In fMRI investigations carried out with samples of healthy older adults, bilinguals showed more frontoparietal and frontal/occipital functional connectivity (FC) (Luk et al., 2011) and stronger intrinsic connectivity than monolinguals in the frontoparietal control network and DMN, as well as stronger correlations between intrinsic connectivity of this control network and task-related increases in activity in prefrontal and parietal regions (Grady et al., 2015).
Neuroplastic effects in circuits linked to the executive and attentional demands of language processing have been proposed as the neural mechanism through which bilingualism compensates for brain damage in dementia (Perani and Abutalebi, 2015; Perani et al., 2017). However, the mechanism of neural compensation due to bilingualism has not been explored in pre-clinical stages of the disease, such as MCI. Moreover, the previous evidence regarding this mechanism is inconclusive because of the use of healthy samples (Grady et al., 2015; Luk et al., 2011) or neuroimaging techniques that do not directly investigate the temporal synchronization between brain regions (Perani et al., 2017).
Therefore, the aim of our research was to study the role of bilingualism in CR by comparing FC at rest in bilingual and monolingual subjects suffering from MCI. Based on the previous study on this topic (Perani et al., 2017), bilinguals were expected to show higher FC in regions included in the language network, ECN, and DMN, when compared with monolinguals. To comprehensively investigate brain patterns in MCI bilinguals and monolinguals during resting state, we also explored possible bilingualism effects on the synchronization and amplitude of regional spontaneous activity.
Methods
Participants
Eighty-one MCI individuals were recruited for this study (40 women; mean age = 73.83 ± 5.69). All of the participants were selected from dementia units of the Valencian Community public health care system. The diagnostic and inclusion criteria were the following: (1) subjective memory complaints (self-reported or confirmed by an informant), (2) objective memory impairment assessed with the logical memory subtest II of the Wechsler Memory Scale-III (Wechsler, 1997), (3) essentially intact activities in daily living, (4) no evidence of dementia, and (5) a Clinical Dementia Rating score of 0.5.
Participants were excluded if they had: (1) other nervous system diseases, such as a brain tumor, cerebrovascular disease, encephalitis, or epilepsy, or met the criteria for dementia; (2) a Geriatric Depression Scale (Martínez de la Iglesia et al., 2002; Yesavage et al., 1982) score >6; (3) visible cerebral abnormalities, such as leukoaraiosis and infarction, reported by a radiologist with experience in magnetic resonance images; and (4) a current psychiatric disorder or use of psychoactive medication.
All the participants were born in Spain and resided permanently in the Spanish region of Valencia. During a clinical interview, they were asked about their use of languages. Participants who reported only speaking Spanish were considered monolinguals (n = 50), whereas those who reported Catalan as their native language, Spanish as a second language, and active daily use of both were considered bilinguals (n = 31). The two groups shared similar everyday life settings and circumstances, such as neighborhood of residence and school/workplace environment.
It should be noted that, for the sake of simplicity, we use the term monolingual to refer to the participants who only speak Spanish. Nevertheless, they could also be referred to as passive bilinguals because monolinguals who permanently reside in Valencia and do not speak Catalan are generally able to understand it.
All the participants were informed of the nature of the research, and they provided written informed consent before their participation in the study. All the study procedures were approved by the Clinical Research Ethics Committee of the Bellvitge University Hospital (Clinical Trial Registration No. PR020/15) and conformed to the Code of Ethics of the World Medical Association (Declaration of Helsinki).
Neuropsychological assessment
All the participants underwent a structured clinical interview and a neuropsychological assessment, including the following tests: Mini-Mental State Examination (MMSE) (Folstein et al., 1975), Functional Activities Questionnaire (Pfeffer et al., 1982), Boston Naming Test (Serrano et al., 2001), a Word List Acquisition and Recall test, two fluency tests (phonetic and semantic), a remote memory test, and the clock-drawing test (Cacho et al., 1996). Descriptive statistics of the sociodemographic variables and results of a two-sample t-test for each of the neuropsychological tests are reported in Table 1. There were no significant differences between the groups on any neuropsychological or sociodemographic variables.
Sociodemographic and Neuropsychological Variables for Bilinguals and Monolinguals with Mild Cognitive Impairment
χ2, chi-squared test; FAQ, Functional Activities Questionnaire; M, mean; M/F, males/females; MMSE, Mini-Mental State Examination; N, sample size; SD, standard deviation; t, t-value for two-sample t-test.
Functional MRI acquisition
Images were acquired on a 3T MRI scanner (Siemens Magnetom Trio, Erlangen, Germany). Participants were placed in a supine position inside the scanner, and their heads were immobilized with cushions to reduce motion. During the acquisition, they were instructed to simply rest with their eyes closed, trying to let their minds go blank and not to fall asleep. A total of 270 volumes were collected over 9 min using a gradient-echo T2*-weighted echo-planar imaging sequence (TR = 2000 ms; TE = 30 ms; matrix, 64 × 64; FOV, 224 × 224 cm; flip angle, 90°; 33 slices, parallel to the hippocampus; slice thickness, 3.5 mm; slice gap, 0.5 mm).
Image preprocessing and statistical analyses
We used Data Processing and Analysis for Brain Imaging (DPABI V4.2_190919) to carry out resting-state fMRI data processing. Preprocessing included the following: (1) removal of the first 10 volumes of each raw fMRI data set; (2) slice timing correction; (3) realignment using a six-parameter (rigid body) linear transformation; (4) spatial normalization to the Montreal Neurological Institute (MNI) space (voxel size 3 × 3 × 3 mm); (5) removal of spurious variance through linear regression: 24 parameters from the head motion correction, linear, and quadratic trends, and the first 5 principal components associated with WM and cerebrospinal fluid (Behzadi et al., 2007); (6) spatial smoothing with a 4-mm full-width at half-maximum (FWHM) Gaussian kernel; and (7) band-pass temporal filtering (0.01–0.1 Hz).
Participants with more than 1 mm/degree of movement in any of the 6 directions or fewer than 120 volumes with framewise displacement (FD) <0.5 mm (Jenkinson et al., 2002) (ensuring at least 4 min of rest with low FD) were excluded from the analyses.
Resting-state FC analysis
A seed-based correlation analysis was performed using a priori regions of interest (ROIs). ROIs were defined from the CONN network cortical ROI atlas (Human Connectome Project-Independent Component Analysis) included in the CONN toolbox. Following the previous study on this topic (Perani et al., 2017), we focused our analysis on the following networks: DMN, ECN, and the language network. To avoid the introduction of different amounts of noise derived from the signal average of regions with different sizes, we used the representative local maxima provided on the atlas to create spherical masks (5 mm radius) as our seeds.
Specifically, the medial prefrontal cortex (MNI: 1, 55, −3), left (MNI: −39, −77, 33) and right parietal gyri (MNI: 47, −67, 29), and posterior cingulate cortex (MNI: 1, −61, 38) were considered seeds for the DMN. Left (MNI: −43, 33, 28) and right (MNI: 41, 38, 30) prefrontal cortices and left (MNI: −46, −58, 49) and right (MNI: 52, −52, 45) posterior parietal cortices were considered ROIs for the ECN. Finally, for the language network, left (MNI: −51, 26, 2) and right (MNI: 54, 28, 1) inferior frontal gyri and the left (MNI: −57, −47, 15) and right (MNI: 59, −42, 13) posterior parts of the superior temporal gyri (pSTG) were used.
After the estimation of individual correlation maps, group-level voxel-wise analyses restricted to an inclusive mask comprising the brain networks under study (i.e., the DMN, ECN, and language network) were performed.
Regional homogeneity analysis
We used the regional homogeneity (ReHo) method to explore group differences in regional synchronization of fMRI time courses between neighboring voxels. For this analysis, Steps 6 and 7 of preprocessing were modified. After spatial normalization, band-pass temporal filtering (0.01–0.1 Hz) was applied, and Kendall's coefficient of concordance was calculated between each voxel's BOLD time series and those of its 19 neighbors. The ReHo value of each voxel was divided by the global mean ReHo of the whole-brain mask, and the resulting ReHo maps were smoothed with a 4-mm FWHM Gaussian kernel (Chao-Gan and Yu-Feng, 2010). Then, group-level, whole-brain, voxel-wise analyses were performed.
Fractional amplitude of low-frequency fluctuations analysis
Differences in the amplitude of regional spontaneous activity between groups were explored using the fractional amplitude of low-frequency fluctuations (fALFF) method. For this analysis, Step 7 of preprocessing was modified. After performing spatial smoothing, the time series of each voxel was transformed into the frequency domain and band-pass filtered (0.01–0.08 Hz). Then, the square root was calculated at each frequency of the power spectrum, the averaged square root (i.e., ALFF) was obtained at each voxel, and a ratio of total amplitude within the low-frequency range to the total amplitude of the detectable frequency range was calculated (i.e., fALFF). Lastly, group-level, whole-brain, voxel-wise analyses were performed.
Statistical analyses
Group differences in FC, ReHo, and fALFF values were investigated using a two-sample t-test, with FD values as a covariate, as implemented in SPM12 (Statistical Parametric Mapping 12; Wellcome Trust Centre for Neuroimaging, University College, London, United Kingdom). The statistical significance was determined using cluster-based inference at a threshold of p < 0.05, family-wise error corrected, with a primary voxel-level threshold of p < 0.001 uncorrected.
Correlation with cognitive status
We explored the relationship between the differences in resting-state measures found in our sample and their cognitive status. To do so, for each brain region showing significant differences in group analyses of any of the measures, a 5 mm3 sphere mask centered in its local maxima was defined (see Table 2 for MNI coordinates). Then, the specific values of the voxels within the mask were averaged in each subject separately, and these averaged values were used to perform a correlation analysis with the MMSE scores, using p < 0.05 as a statistically significant threshold, for the sample as a whole and for each group separately.
List of Regions with Higher Functional Connectivity, and Thalamic Nuclei with Higher Fractional Amplitude of Low-Frequency Fluctuations in Bilinguals Compared to Monolinguals
Brain regions based on the AAL atlas (Tzourio-Mazoyer et al., 2002) and subcortical nuclei based on a recently proposed thalamic segmentation (Najdenovska et al., 2018).
AAL, automated anatomical labelling; AN, anterior nucleus; BA, Brodmann's area; CMPN, central-medial pulvinar nucleus; fALFF, fractional amplitude of low-frequency fluctuations; K, cluster size in voxels; LN, language network; MDN, mediodorsal nucleus; MNI, Montreal Neurological Institute; pSTG, posterior superior temporal gyrus; t, t-value for two-sample t-test; VLVN, ventral-lateroventral nucleus.
Results
Seed-based connectivity analysis
We found significant differences between bilinguals and monolinguals in the FC of the left pSTG seed of the language network. Specifically, the connectivity of this region with the precuneus was higher in bilinguals than in monolinguals (Fig. 1 and Table 2). The opposite contrast did not show any significant results (monolinguals>bilinguals). No other significant differences were found in any other seed.

Differences in FC of the left pSTG between bilinguals and monolinguals. In blue, the location of the left pSTG seed. The color bar represents the t-value of areas with significantly higher FC with this seed in bilinguals compared with monolinguals. FC, functional connectivity. Color images are available online.
ReHo analysis
We found no significant differences in regional synchronization in bilinguals compared with monolinguals.
fALFF analysis
We found a higher amplitude of regional spontaneous activity in bilinguals compared with monolinguals in the thalamus bilaterally (Fig. 2 and Table 2). Specifically, the differences appeared in the left mediodorsal and central-medial pulvinar and right anterior and lateroventral nuclei of the thalamus (Najdenovska et al., 2018). The opposite contrast did not show any significant results.
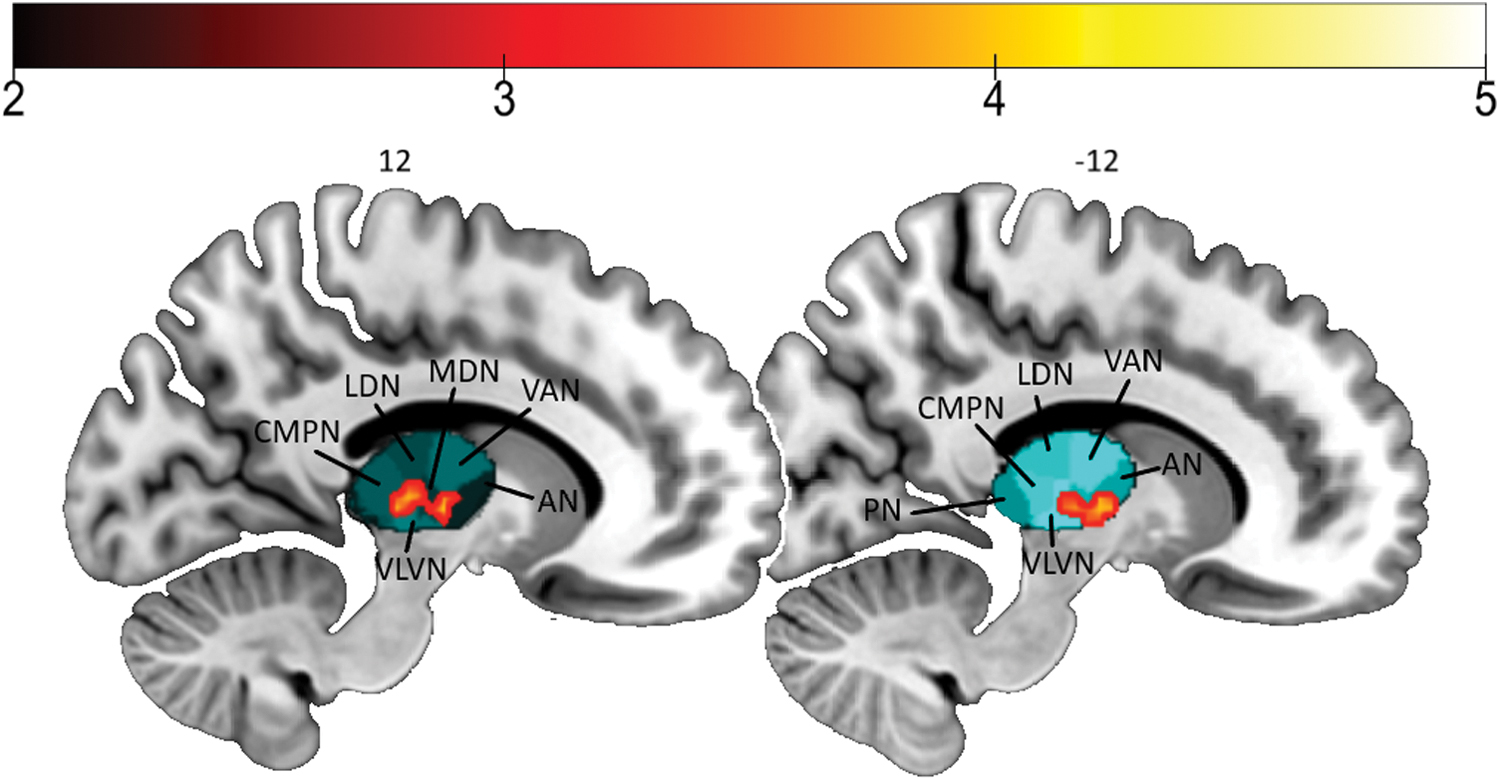
Differences between bilinguals and monolinguals in fALFF. The color bar represents the t-value of areas with significantly higher fALFF in bilinguals compared with monolinguals. Thalamus nuclei are shortened as follows: AN, anterior; CMPN, central-medial pulvinar; fALFF, fractional amplitude of low-frequency fluctuations; LDN, laterodorsal; MDN, mediodorsal; PN, pulvinar; VAN, ventral anterior; VLVN, ventral-lateroventral. Color images are available online.
Correlation with cognitive status
We found a significant correlation between our sample's performance on MMSE and FC between the left pSTG seed of the language network and the precuneus (R = 0.272, p = 0.014). We found no significant correlations for the groups separately or for the differences in fALFF.
Discussion
In this study, we investigated the FC, ReHo, and fALFF differences between bilingual and monolingual MCI subjects who had no significant differences in age, years of schooling, proportion of men and women, or performance on neuropsychological testing. Bilinguals showed higher FC than monolinguals between the left pSTG of the language network and the precuneus, and higher fALFF in several nuclei of the thalamus. Moreover, FC values between the pSTG and precuneus correlated with the MMSE scores in the whole sample. These results suggest that the experience of bilingualism promotes CR through neural compensation.
Our results are consistent with previous evidence suggesting that bilingualism may contribute to neural compensation in dementia (Luk et al., 2011; Perani et al., 2017). A previous study showed enhanced WM integrity and more functional connections involving frontal, parietal, and occipital lobes in healthy older bilinguals compared with monolinguals (Luk et al., 2011). These results were subsequently interpreted as a possible compensatory mechanism that would provide reserve and compensate for GM deterioration (Guzmán-Vélez and Tranel, 2015).
In a later study analyzing metabolic connectivity in a sample of patients with dementia, bilinguals showed increased connectivity compared with monolinguals between the precuneus/posterior cingulum and the anterior cingulum, orbitofrontal cortex, thalamus, and caudate nucleus, all described as crucial brain regions for language control in bilinguals (Perani et al., 2017). Since bilinguals also showed more hypometabolism than monolinguals, the increased connectivity was also interpreted as a compensatory mechanism by which the bilingual brain would be able to cope better with neurodegeneration (Perani et al., 2017).
Along the same lines, our study showed that, in a sample of subjects with MCI, bilinguals exhibited higher levels of FC than monolinguals between the left pSTG, an area within the language network, and the precuneus, a region typically affected in dementia. Therefore, we interpret our findings as neural compensation and not neural reserve, validating previous evidence using indirect measures of interregional connectivity (Perani et al., 2017). Moreover, we also found a positive correlation between MMSE performance and FC between the precuneus and pSTG for the whole sample, further supporting the relationship between cognitive status and FC between these brain regions.
Perani and colleagues (2017) also found increased anterior/posterior metabolic connectivity in the ECN in bilinguals compared with monolinguals. However, we found no significant FC differences between bilinguals and monolinguals in the ECN seeds. This may be due to the different characteristics of our samples: in their investigation, participants were dementia patients, whereas our work involved MCI subjects.
In the first stages of Alzheimer's disease, which is the most common form of dementia in elder populations, brain pathology, such as β-amyloid accumulation (Palmqvist et al., 2017), tau deposition (Hall et al., 2017), and hypometabolism (Sperling et al., 2011), is mainly limited to the DMN areas, and especially to the precuneus/posterior cingulum. Therefore, the fact that the differences in our sample are restricted to the FC of this region may be due to the early stage of the disease in our sample. Along the same lines as previous investigations (Grady et al., 2015; Luk et al., 2011; Perani et al., 2017), our results suggest that bilingualism may be acting as a CR factor through neural compensation mechanisms.
We also found higher fALFF in bilinguals compared with monolinguals in multiple nuclei of the thalamus. Previous studies show that fALFF values tend to decrease in prodromal AD and MCI (Cha et al., 2015; Zeng et al., 2019). Regarding the thalamus and its role in dementia, a review based on postmortem studies, animal models, and noninvasive imaging investigations suggests that the loss of episodic memory in early stages of dementia is not mainly related to hippocampal dysfunction, but rather to broader neurodegeneration of the Papez circuit, an extended memory system that involves the limbic thalamus (Aggleton et al., 2016).
Thus, previous studies found reduced thalamic volumes in amnestic MCI subjects (Pedro et al., 2012; Sorg et al., 2007; Yi et al., 2016) and correlations between thalamic volume and cognitive status in MCI (Pedro et al., 2012; Yi et al., 2016). Moreover, bilateral atrophy in the dorsomedial thalamus, reductions in WM integrity in the anterodorsal nucleus, and smaller internal medullary lamina were found in dementia patients compared with controls (Zarei et al., 2010). In our study, the differences found in fALFF in the thalamus were restricted to the left mediodorsal and central-medial pulvinar and right anterior and lateroventral nuclei, based on the human thalamic segmentation proposed in a recent investigation (Najdenovska et al., 2018).
Although the relationship between memory impairment and thalamic lesions has been described for years (Carlesimo et al., 2011; Danet et al., 2017; Fedio and Van Buren, 1975; Harding et al., 2000), there is a lack of agreement about the specific nuclei responsible for this relationship: early investigations found verbal memory deficits due to left pulvinar nucleus stimulation (Fedio and Van Buren, 1975), neuronal loss in the anterior nuclei due to Korsakoff's syndrome was associated with amnesia (Harding et al., 2000), and mediodorsal nucleus lesions due to left thalamic stroke were related to impaired recollection (Danet et al., 2017).
Based on these studies, we tested if the differences in fALFF in the thalamus found in our sample were correlated with the scores of any of the memory tests used, but we found no significant results. A possible explanation for this lack of relationship between memory status and the amplitude of regional spontaneous activity in the thalamus could be the low variability in the memory scores of the participants in our sample. Moreover, previous studies state that functional alterations in fMRI can be detected before the manifestation of cognitive decline and clinical deterioration (Sheline and Raichle, 2013; Sperling et al., 2011). Thus, another possibility is that the differences found in our sample in fALFF in the thalamus might not have manifested behaviorally yet.
Also importantly, other investigations suggest that the thalamus is related to language processing and bilingualism, based on evidence showing that corticothalamo-cortical connections have a pivotal impact on language processing through feedback mechanisms (Crosson, 2013) and that the thalamus is expanded in young simultaneous bilinguals compared with monolinguals (Burgaleta et al., 2016). Thus, our results suggest that bilingualism may act as a CR factor by means of a higher amplitude of regional spontaneous activity in the thalamus, specifically in nuclei that show atrophy in dementia (Zarei et al., 2010) and are related to memory impairment (Danet et al., 2017; Fedio and Van Buren, 1975; Harding et al., 2000).
As a general conclusion, our results show that, in a sample of MCI subjects with the same disease severity, proportion of men and women, years of schooling, and sociocultural characteristics, bilinguals manifest higher FC than monolinguals between the pSTG of the language network and the precuneus, and higher fALFF in the thalamus. These results expand our knowledge about the effects of the active use of two languages on brain function, and they support the role of bilingualism as a CR factor that protects against dementia through neural compensation.
Footnotes
Authors' Contributions
V.C. and L.M.-M. conceptualized the study and were responsible for implementation of data analyses. V.C., L.M.-M., M.-Á.P.-G., A.M.-P., N.A., J.A.-V., and E.V.-R. were involved in interpreting the findings, and drafting and revising the article.
Author Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This work was supported by the project (201410-30-31) provided by Fundació Marató TV3. Lidón Marin-Marin, Naiara Aguirre, and Jesús Adrián-Ventura were supported by an FPU grant from the Spanish Ministry of Education, Culture and Sports, and Esteban Villar-Rodríguez, by an FPU grant from the Ministry of Science, Innovation and Universities. María-Ángeles Palomar-García and Anna Miró-Padilla were supported by a postdoctoral graduate program grant (University Jaume I) and Victor Costumero was supported by a Juan de la Cierva postdoctoral graduate program grant from the Spanish Ministry of Economy, Industry and Competitiveness (IJCI-2016-29247).