
Abstract
Background:
It is becoming increasingly recognized that there is significant interneuron degeneration in Alzheimer's disease. As the hippocampus is integral for learning and memory, we performed a systematic review of primary literature focused on the relationship between Alzheimer's and hippocampal interneurons. In this study, we summarize the experimental work performed to date and identify opportunities for future experiments.
Objectives:
This PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses)-style systematic review seeks to summarize the findings of all accessible research focused on cholecystokinin (CCK), neuropeptide Y (NPY), parvalbumin (PV), and somatostatin (SOM) interneurons in the hippocampal formation.
Results:
One thousand five hundred ninety-three articles were pulled from PubMed, PsycInfo, and Web of Science, based on three blocks of search terms. There were 45 articles that met all the predetermined inclusion/exclusion criteria. There is strong evidence that PV interneurons are affected early in the disease by toxic amyloid beta (Aβ) fragments; SOM interneurons are affected indirectly while the SOM neuropeptide may act to slowly worsen toxic Aβ fragment accumulation, whereas NPY- and CCK-positive interneurons are affected later in the progression of the disease.
Conclusions:
Fewer studies have been performed on NPY and CCK interneurons, and there is room for further investigations regarding the role of PV interneurons in Alzheimer's to help resolve contradictory findings. This review found that PV interneurons are affected early in the disease, but only in Alzheimer's precursor protein but not tau models. NPY and CCK interneurons were found to be affected later in the disease, and SOM interneurons vary greatly. Future studies may consider reporting immunohistochemical studies inclusive of either cell location or morphology—as well as marker to give a more robust picture of the disease.
Impact statement
In the past decade, the contribution of hippocampal interneurons to the pathology of Alzheimer's disease has been increasingly recognized. The goal of this systematic review was to summarize findings to better determine the focus of the work to date, to help identify where there is consensus in the research, and to elucidate future avenues of inquiry.
Introduction
Alzheimer's disease (AD) is a debilitating neuropathology that is becoming increasingly prevalent in modern societies of aging population (Ballard et al., 2011; Patterson, 2018). According to the 2018 World Alzheimer's Report, more than 50 million people are affected, and the global cost exceeds 1 trillion dollars (Patterson, 2018). Our understanding of AD pathology has advanced recently to shift from a focus on tau and amyloid beta (Aβ) in principal neurons to the roles they may play in altering interneurons (Najm et al., 2019; Rozycka and Liguz-Lecznar, 2017; Solarski et al., 2018; Verret et al., 2012; Vico Varela et al., 2019). The systematic nature of this review will add to the field by summarizing knowledge about how different interneuron subtypes contribute to the pathology of AD and, more importantly, by determining areas that could benefit from replication, where there is a lack of consensus, as well as identifying knowledge gaps in the current literature. This review includes work performed in mouse, rat, and human studies that focus on questions directly related to the relationship between interneurons and AD in the hippocampal formation.
It is well established that dysfunction in the structure and metabolism of tau and Alzheimer's precursor protein (APP) are strongly associated with AD, and this has led to the theory that an initial dysregulation of APP metabolism causes toxic Aβ to accumulate and leads to the aggregation of phosphorylated tau neurofibrillary tangles and neurodegeneration (Bloom, 2014). Recently, it has been discovered that APP dysregulation seems to affect interneurons initially, leading to speculation that the loss of interneurons could then cause aberrant neural activity and excitotoxicity in principal cells (Kiss et al., 2016; Palop et al., 2007). As this degeneration progresses regionally, the circuit changes can also affect cortical and basal brain regions differently (Fewster et al., 1991). Interneuron subtypes can be segregated immunohistochemically in the hippocampus, with antibodies specific for parvalbumin (PV), somatostatin (SOM), cholecystokinin (CCK), or neuropeptide Y (NPY) being the main ways used to differentiate these populations (Pelkey et al., 2017). While calretinin and calbindin are also markers assessed in interneurons, these markers are expressed across a wider variety of interneurons and not considered as their own distinct subclasses (Rogers, 1987). For the purpose of this review, hippocampal interneuron subtypes were simplified as immunohistochemically defined. A strength of immunohistochemical studies is that studies in postmortem human tissue may be included. This allows for parallels to be drawn between mice and rats exposed to specific homogeneous disease conditions and the clinical population. Immunohistochemical studies, however, are often limited by the lack of ability to view changes over time or in three dimensions. In rodent studies, optogenetics and whole-brain clearing with sheet fluorescence confocal microscopy can be used to overcome this. From a technical standpoint, immunohistochemical studies must be carefully validated; antibody cross-reactivity and specificity can make cells erroneously appear positive for a marker while darkness of staining between replicates must be kept constant for accurate evaluation.
CCK was first discovered as a digestive hormone; however, it is also a specific marker for interneurons in the hippocampus. CCK has two main G-protein-coupled receptors: CCKAR and CCKBR (Nishimura et al., 2015). CCKBR is the predominant receptor in the brain, and the activation of this signal cascade has been linked to anxiety-like behavior (Nishimura et al., 2015; Singh et al., 1991). CCK-positive interneurons colocalize with the cannabinoid receptor type 1 (CB1) and likely play an important role in the interaction between delta-9-tetrahydrocannabinol (THC) consumption and AD (Lee and Soltesz, 2011). There is an abundance of articles that assess cannabis and cannabis-derived compounds as a potential treatment for AD (Aso et al., 2015; Brown University, 2019; Janefjord et al., 2014; Li et al., 2020; Shelef, et al., 2016). Two main potential mechanisms have been identified. The first is through Gi/o-linked CB1 receptors that are primarily located in the hippocampus on CCK-positive interneurons. Effects have also been associated with cannabinoid receptor type 2 (CB2) receptors, which are more prevalent on astrocytes and in the peripheral nervous system (Scheyer et al., 2019). To date, there is a lack of consensus on whether cannabis treatment is helpful or harmful, with studies varying greatly across exposure paradigms; see Charernboon and colleagues (2021) for a systematic review on treatment outcomes. Regardless of whether cannabis is effective for treating AD, CB1 receptor activation causes a signal cascade that inhibits neurotransmitter release (i.e., GABA in CCK interneurons) (Scheyer et al., 2019). CCK-positive interneurons are therefore necessary to consider when assessing inhibition/excitation imbalance in AD.
NPY also has functions as a gastric hormone as well as a neuro-signaling molecule (Sundler et al., 1983). In the brain, NPY acts upon a host of postsynaptic G-protein-coupled receptors (Duarte-Neves et al., 2016). NPY-positive interneurons have been implicated in adult neurogenesis, a mechanism that is thought to occur in the dentate gyrus (DG) (Decressac et al., 2009). Adult neurogenesis is the process of new granule cell growth from a pool of neural stem cells, and this is thought to be integral to memory consolidation (Snyder, 2019). NPY has been shown to increase neural precursor proliferation, and the number of NPY-positive interneurons has been shown to decline from young adulthood to middle age (Hattiangady et al., 2005; Spencer et al., 2016). NPY has also been implicated in post-traumatic stress disorder, Huntington's disease, and epilepsy (Schmeltzer et al., 2016; Wagner et al., 2016; Wickham et al., 2019).
PV is a calcium binding protein, and changes in PV affect presynaptic calcium signaling (Collin et al., 2005). Calcium signaling is a driving force behind neurotransmitter release, and the amount of time that calcium is able to bind to its ligands helps determine neuronal signal fidelity (Branch et al., 2014). PV-containing basket interneurons make up 15% of all interneurons in the CA1 hippocampal subregion (Pelkey et al., 2017). PV-containing basket cells are associated with both theta and gamma waves in the hippocampus, although they preferentially act in gamma oscillations (Chung et al., 2020). They are thought to be integral to lateral inhibition, memory consolidation, and made up the bulk of the results in the articles that we identified (Espinoza et al., 2018; Ognjanovski et al., 2017). PV interneurons are implicated in the seizure component of AD, and some studies have found these cells to decrease in number over time (Albuquerque et al., 2015; Hijazi et al., 2019; Mahar et al., 2017).
SOM was first discovered in the pituitary, and while it is expressed on GABAergic neurons, there was debate as to the function of SOM itself (Boehm and Betz, 1997; Brazeau et al., 1973). There are five SOM receptors, four of which have been found in the hippocampus (Cammalleri et al., 2019). SOM receptors are all G-protein-coupled receptors and can be found pre- and postsynaptic (Cammalleri et al., 2019; Hoyer et al., 1995). SOM signaling acts as an inhibitory molecule through action on potassium channels, leading to hyperpolarization of the postsynaptic neuron (Schweitzer et al., 1998). SOM-positive interneurons act in the creation of theta oscillations, have been found to be decreased in AD brains, and tend to be found in the stratum oriens lacunosum moleculare layer in the hippocampus (Chung et al., 2020; Schmid et al., 2016). Recent studies have shown that they accumulate phosphorylated tau, and new evidence suggests that they facilitate hippocampal–prefrontal synchrony and spatial encoding (Abbas et al., 2018; Zheng et al., 2020).
Here, we use a modified Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guideline to review the literature published on SOM-, PV positive, CCK-, and NPY-positive interneurons in the hippocampus in relation to the cellular pathogenesis of AD in the hippocampus.
Methods
This review was completed under the stylistic criteria of the PRISMA. Minor revisions were made to the traditional process due to the nature of cellular studies (e.g., exclusion of bias scoring) as reported by Moher and colleagues (2009). In lieu of bias scoring, articles that recommended a treatment were excluded. PRISMA search terms were designed to capture all articles that discussed hippocampal CCK, SOM, NPY, or PV interneurons and AD, and then, articles were selected based on the inclusion and exclusion criteria (Supplementary Table S2). Within search blocks, similar terms were grouped using “OR” as an operator, and between search blocks an “AND” operator was used. Three blocks of search terms were used. Blocks of search terms required articles to (1) mention interneurons, (2) mention the hippocampus, and (3) specify Alzheimer's Disease or Dementia (Supplementary Table S3). The search was completed in three databases, PubMed, PsycINFO, and Web of Science, by two authors individually using the same terms. Dates of coverage for the search included January 1, 2000–June 15, 2020. Citations were all exported to Mendeley, duplicates were removed, and articles were screened using the inclusion/exclusion criteria (Supplementary Table S2). Initially, titles and abstracts were screened, and if the article showed potential relevance, the article was read in full. Final articles were compared between the two authors, and discrepancies were resolved through discussion. A forward and reverse search was completed from the selected articles' reference lists to identify any additional relevant articles.
Results
Figure 1 summarizes the selection process. A total of 1598 records were identified through database searching and additional sources. We identified 73 records after review of title and abstract at an initial screening. Twenty-eight of these records were then excluded after full-text article screening as 10 records did not study solely AD model or include an AD model, 12 did not study CCK, PV, SOM, or NPY interneurons, 3 were not in the hippocampus, 2 were not research articles, and 1 used a treatment. These excluded articles are referenced in Supplementary Table S1. Forty-five articles met the inclusion criteria and were included in the qualitative analysis. The number of studies found to analyze CCK, NPY, PV, and SOM hippocampal interneurons was 5, 10, 37, and 20, respectively. Of these 45 articles, 5 studied human AD subjects, 6 studied rat AD models, and 36 studied mice AD models. All the five human AD studies used both male and female subjects. Of the 36 mice studies, 15 studies were on male mice, 1 study was on females, 8 studies included both sexes, and 12 studies did not specify sex. Four of the AD rat studies included only male rats, whereas two studies included only female rats (Supplementary Table S4).
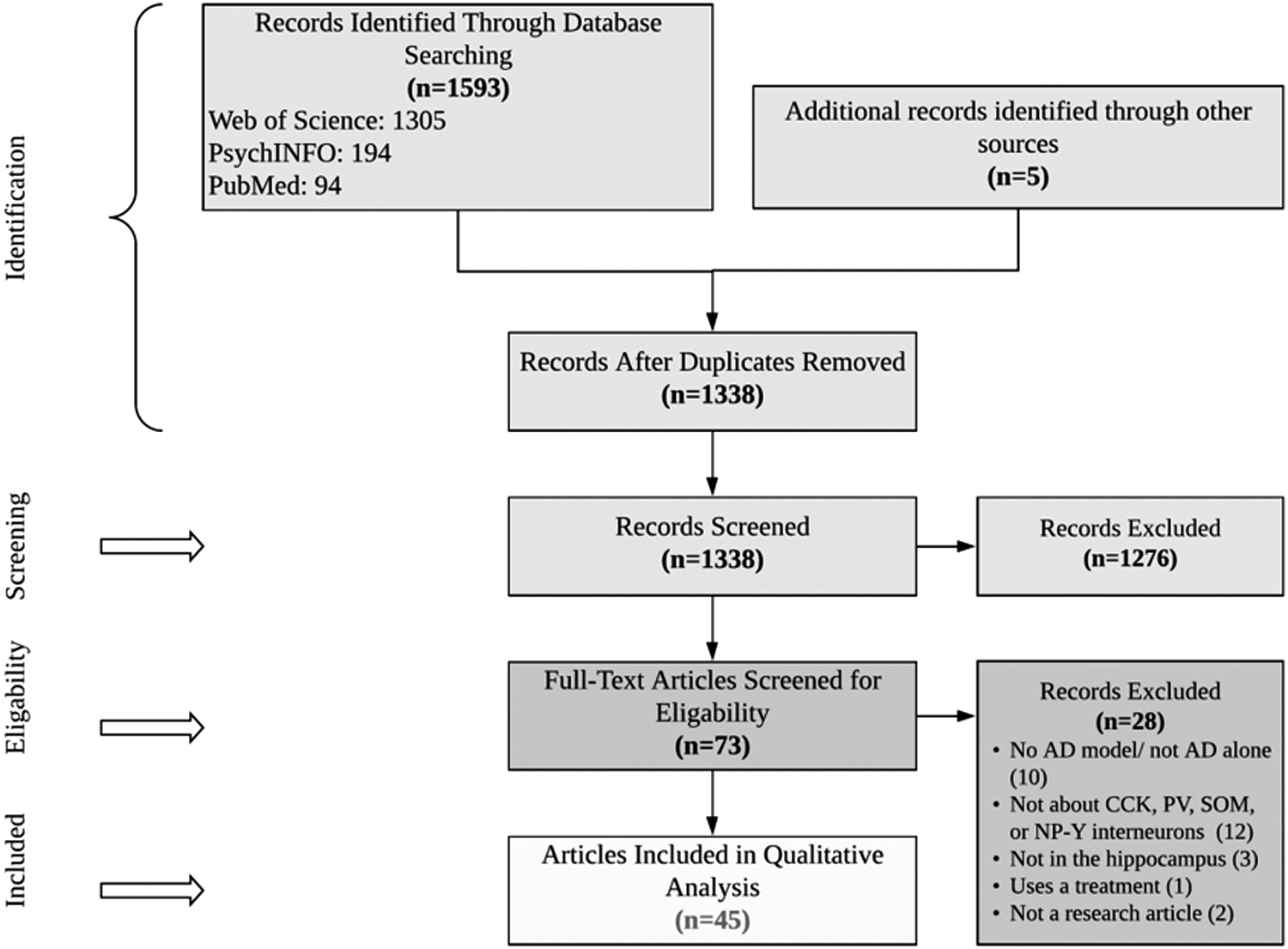
PRISMA flow diagram for the selection of articles included in this systematic review. PRISMA, Preferred Reporting Items for Systematic Reviews and Meta-Analyses.
Figure 2 shows that most studies identified were performed in mice, and that most studies were sex-balanced. Studies with “Unspecified” as sex were almost completely mouse studies and were likely sex-balanced as well. Figure 3 panels show the types of assay performed in each cell type, and the colour of the bar indicates if the finding was an increase, decrease, or null finding from baseline. For the purpose of bucketing results, experiments that used multiple markers (of the main four investigated), experiments that validated a method, and duplicate experiments between a hippocampal region and subfield were excluded from this figure.
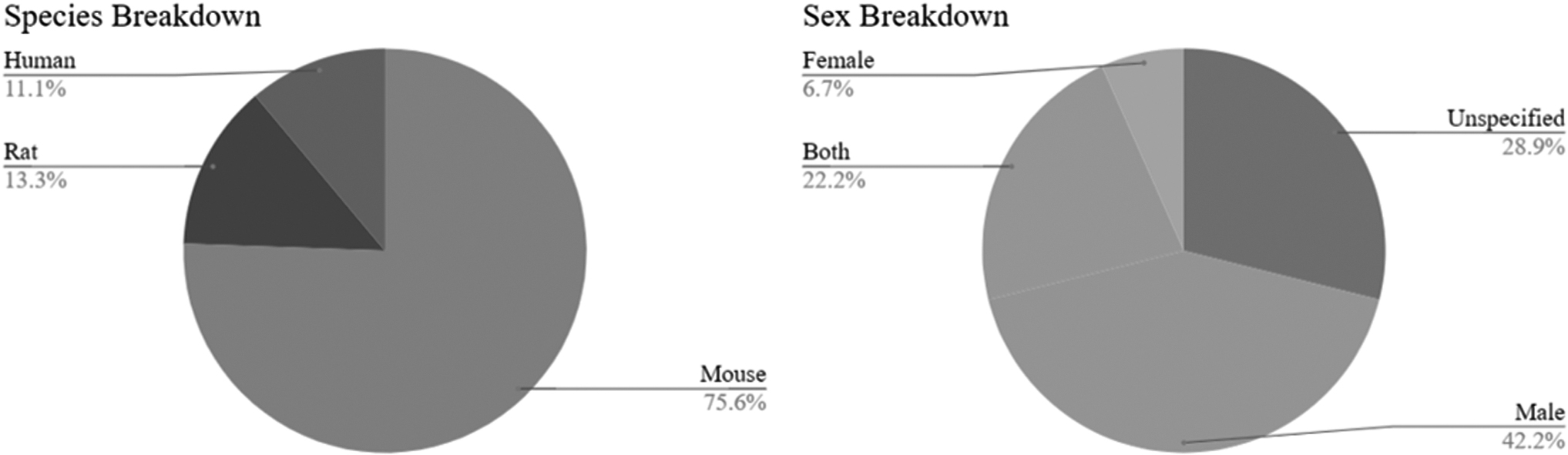
Breakdown of sex and species used in the 45 articles included in this systematic review.
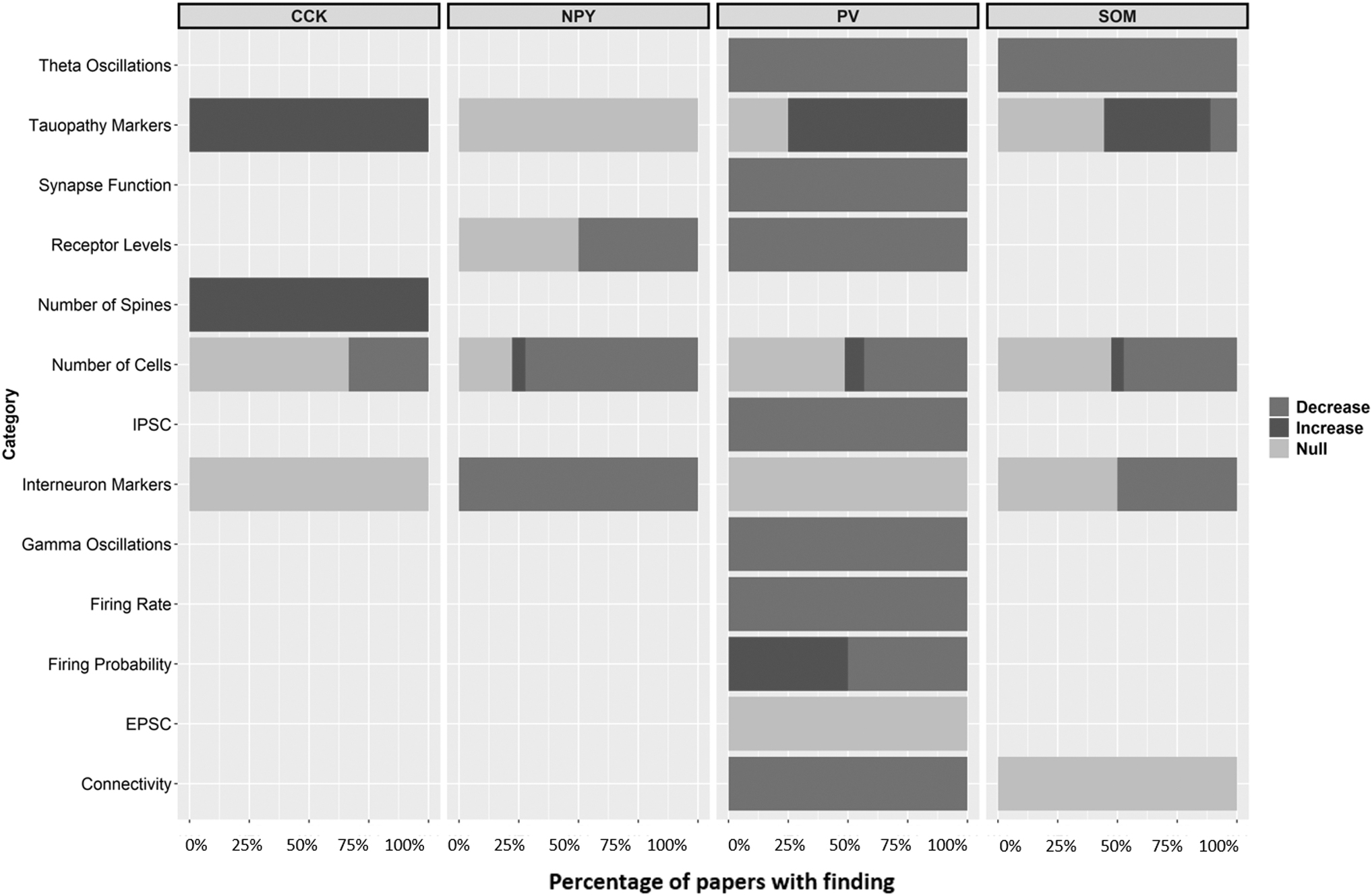
Summary of experimental findings across all articles included in the study showing agreement or disagreement in the literature.
Cholecystokinin
Five articles were found to focus on CCK hippocampal interneurons. The distribution of species and sex studied is listed in Table 1. Three articles assessed CCK immunoreactivity in the hippocampus. Two of these articles showed no change in CCK-positive cells after Aβ injections (Aguado-Llera et al., 2018; Villette et al., 2012), whereas the other article showed a decrease in CCK-positive cells only at 4 months and 9–18 months (Shi et al., 2020). Similarly, two articles assessed CCK mRNA content in the hippocampus and found no change in PS1xAPP mice (Ramos et al., 2006) or in rats after a 14 day infusion of Aβ25–35 (Aguado-Llera et al., 2018). One study assessed Aβ accumulation in CCK interneurons and found an age-dependent increase from 9 to 18 months in APP NL-F/NL-F mice (Shi et al., 2020). Finally, one study found that CCK-8S administration into an AD mouse model increased the spine density of hippocampal neurons (Zhang et al., 2013). This work showed that three of the five articles found evidence that hippocampal interneurons are not affected until older age in animal models.
Summary of Cholecystokinin Experiments in Identified Literature
Aβ, amyloid beta; APP, Alzheimer's precursor protein; CCK, cholecystokinin; H, hippocampus; M, male; RT-PCR, reverse-transcriptase PCR; U, unspecified; WT, wild type.
Neuropeptide Y
To better understand the role of NPY interneurons in AD, we assessed articles that examined hippocampal NPY interneurons in rat and mice AD models. No studies were found to assess human AD tissue. The distribution of species and sex studied is listed in Table 2. Seven articles assessed the immunoreactivity of NPY in various layers and regions of the hippocampus in AD, and six of these articles showed decreased NPY immunoreactivity in AD models (Albuquerque et al., 2015; Leung et al., 2012; Loreth et al., 2012; Mahar et al., 2017; Moreno-Gonzalez et al., 2009; Ramos et al., 2006). Specifically, three articles discovered decreased NPY immunoreactivity in the CA1, CA2, and DG regions of mouse AD models at 1 month (Mahar et al., 2017), 6 months (Albuquerque et al., 2015), and 12 and 18 months (Loreth et al., 2012). One article did not observe any change to NPY immunoreactivity in the CA1 region 24 days after microinjection of Aβ40 and Aβ45 compared with controls (Villette et al., 2012), whereas another article did not observe any change in the subiculum region at 6 months in a mouse AD model (Albuquerque et al., 2015). One study focused on the entorhinal cortex region and found decreased NPY immunoreactivity in PS1xAPP mice at 6 months compared with controls (Moreno-Gonzalez et al., 2009).
Summary of Neuropeptide Y Experiments in Identified Literature
AICD, amyloid precursor protein intracellular domain; DG, dentate gyrus; EC, entorhinal cortex; F, female; g.r., granule cell layer; Hi, hilus; m.o., molecular layer; NPY, neuropeptide Y; p.o.; p.y.; s.o., stratum oriens; SOM, somatostatin; s.r, stratum radiatum.
Corresponding to decreased NPY immunoreactivity, two articles analyzed NPY mRNA levels in the hippocampus of AD mice and found that these were decreased compared with controls after a 14 day Aβ25–35 infusion into the hippocampus (Aguado-Llera et al., 2018) or in 6-month-old PS1xAPP mice (Ramos et al., 2006). However, one of these articles found that co-administration of IGF-1 with Aβ25–35 restored the reduction of NPY mRNA (Aguado-Llera et al., 2018). There were two studies that assessed NPY receptor levels in AD mice, and one of these articles found no change in NPY receptor levels compared with controls (Ghosal and Pimplikar, 2011), whereas the other study found that NPY-Y1 receptor mRNA levels were decreased, but NPY-Y2 receptor mRNA levels were increased in hAPP-J20 mice compared with wild-type (WT) controls (Palop et al., 2007). Interestingly, one article found that AD mice that expressed higher levels of activity-related cytoskeleton protein (Arc) also had higher NPY levels (Palop, et al., 2007). The majority of studies here support the conclusion that NPY levels are reduced with AD pathology.
Parvalbumin
A total of 37 articles that assessed PV hippocampal interneurons in AD were evaluated. The distribution of species and sex studied is listed in Table 3. Twenty-one articles used immunohistochemistry to quantify the changes seen in PV interneuron levels of various AD models in various regions and layers of the hippocampus. Nine of these articles found a decrease in PV immunoreactivity compared with controls (Ahnaou et al., 2017; Aso et al., 2018; Hoefling et al., 2019; Huh et al., 2016; Levenga et al., 2013; Popovic et al., 2008; Soler et al., 2017; Takahashi et al., 2010; Zallo et al., 2018), two articles found an increase in PV immunoreactivity (Hollnagel et al., 2019; Verdaguer et al., 2015), and five articles found no change compared with controls (Albuquerque et al., 2015; Moreno-Gonzalez et al., 2009; Ramoset al., 2006; Villette et al., 2012; Waller et al., 2020); however, this was not taking into account the specific region within the hippocampus. Additionally, five articles found increases, decreases, or null effects in PV positive cell density depending on the particular hippocampal region studied or the time period assessed (Arevalo-Serrano et al., 2008; Hijazi et al., 2019; Loreth et al., 2012; Mahar et al., 2017; Petrache et al., 2019). For example, one study found no change in the CA1 region of a mouse AD model but found decreased PV positive cell density in the dorsal lateral entorhinal cortex (Petrache et al., 2019). Another article reported decreased PV positive cell density in the CA1 but no change in the CA2 after injection of Aβ1–40 into rats compared with control rats (Arevalo-Serrano et al., 2008). Additionally, Mahar and colleagues (2017) reported that in 1-month-old AD mice, there was decreased PV positive immunoreactivity in the CA1, CA2, and subiculum regions but no change in the DG. Two articles assessed PV positive cell density at multiple time points in an animal model. One article found that the CA1 region had increased PV positive cell density compared with WT controls at 15–17 weeks but had significantly decreased PV positive cell density at 23–25 weeks (Hijazi et al., 2019), whereas the other article found no changes in PV positive cell density in the DG and hilus of a mouse AD model at 12 months but at 18 months found a significant decrease (Loreth et al., 2012).
Summary of Parvalbumin Experiments in Identified Literature
Overall: apoE4 has no effect on PV positive interneuron profiles.
Overall: ApoE4-mediated impairment of GABAergic interneurons in F apoE-KI mice occurs predominantly in the hilus.
AD, Alzheimer's disease; BDA, biotinylated dextran-amine; CB, cannabinoid; CTF, β-secretase-derived fragment; eEPSCs, evoked excitatory post synaptic current; CNO, nitric oxide; ELISA, enzyme-linked immunosorbent assay; GCL, granule cell layer; H-MS, hippocamposeptal; LEC, lateral entorhinal cortex; MF, male and female; NTG, non-transgenic; PARV, parvalbumin; PBS, phosphate-buffered saline; PC, parvalbumin cell; PCL, polymorphic cell layer; PV, parvalbumin; PV-IR, parvalbumin immunoreactivity; sEPSCs, spontaneous excitatory postsynaptic currents; SG, stratum granulare (granule cell layer); SHP, septohippocampal pathway; sIPSC, spontaneous inhibitory postsynaptic currents; s.m., stratum moleculare; s.l., stratum lacunosum; s.r., stratum radiatum; SST, somatostatin; TBS, theta burst stimulation; TG, transgenic; VLW, double-transgenic APP(SW)/Tau.
Five experiments were completed on human AD postmortem hippocampal tissue. Three experiments assessed PV positive cell density using immunohistochemistry, and all the three experiments found no change in PV positive cell density in the DG-SG and hilus (Takahashi et al., 2010) or the CA1 and entorhinal cortex (EC) (Waller et al., 2020). However, all the three experiments were limited to a small sample size (n = 10). Another experiment found that human AD postmortem tissue had decreased levels of a PV-specific voltage-gated sodium channel subunit, Nav1.1 (Verret et al., 2012). Finally, one article found increased pTau accumulation compared with controls (Zheng et al., 2020).
In animal AD models, the most commonly studied hippocampal region, CA1, had nine experiments that show decreased PV positive cell density (Ahnaou et al., 2017; Arevalo-Serrano et al., 2008; Hijazi et al., 2019; Hoefling et al., 2019; Levenga et al., 2013; Mahar et al., 2017; Soler et al., 2017; Takahashi et al., 2010; Zallo et al., 2018), two experiments that show an increase (Hijazi et al., 2019; Hollnagel et al., 2019), and four experiments that show no change compared with controls (Albuquerque et al., 2015; Loreth et al., 2012; Petrache et al., 2019; Villette et al., 2012).
In both the CA2 and CA3 regions of animal AD models, four experiments observed no change in PV positive cell density (Albuquerque et al., 2015; Arevalo-Serrano et al., 2008; Loreth et al., 2012; Mahar et al., 2017), whereas two experiments showed a decrease in the CA2 region compared with controls (Mahar et al., 2017; Takahashi et al., 2010).
In the whole DG, 12 experiments were performed across 9 articles. One experiment showed an increase in mossy inputs in an hAPP-J20 model (Palop et al., 2007). Six experiments concerned the amount of PV positive cells or PV immunoreactivity. Three experiments found no change in AD mice at 1 month (Mahar et al., 2017), 6 months (Albuquerque et al., 2015), and 12 months (Loreth et al., 2012); however, one experiment found decreased PV immunoreactivity at 18 months in AD mice compared with controls (Loreth et al., 2012), and one experiment found a decrease in PV positive cells at 12 months primarily in the dendritic projections of hilar and granular layer neurons. Additionally, one experiment found decreased PV immunoreactivity in 50% of brain slices at 14 months old in APPxPS1 mice compared with controls; however, the sample size was limited (n = 6). Three studies assessed tauopathy in PV interneurons. Levenga and colleagues (2013) found no colocalization with early and late tauopathy markers, Wang and colleagues (2014) found APP immunoreactivity in PV interneurons, and Soler and colleagues (2017) found that some PV positive cells co-labeled with a marker for tau phosphorylated at threonine 231.
Three experiments assessed the entorhinal cortex across four articles in AD animal models. Leung and colleagues (2012) found that male ApoE3 knock-in mice had more PV interneurons than male ApoE4 knock-in mice at 1 month old. Another experiment found no change in the number of PV positive cells in the EC at 6 months in an Aβ accumulation model (Moreno-Gonzalez et al., 2009). Concerning tauopathy, one experiment found slight pTau accumulation in the EC, but less than in the CA1 region, and Mondragon-Rodriguez and colleagues (2018) found slight pTau accumulation in the EC, but less than in CA1.
Two experiments across two articles looked at PV interneurons in the subiculum in animal AD models. Maher and colleagues (2017) found that there were less PV positive interneurons in the subiculum of mice in a tau AD model, and Albuquerque and colleagues (2015) found no change in 6-month-old mice of the same model.
Somatostatin
A total of 20 studies that analyzed the role of SOM interneurons in AD were identified. The distribution of species and sex studied is listed in Table 4. In total, we assessed 13 articles that used immunohistochemistry to assess SOM cell density in a total of 49 experiments in various regions and layers in the hippocampus. A total of 17 experiments exhibited decreased SOM immunoreactivity in AD humans and animal models compared with controls (Aguado-Llera et al., 2018; Albuquerque et al., 2015; Levenga et al., 2013; Moreno-Gonzalez et al., 2009; Ramos et al., 2006; Sekiguchi et al., 2009; Shi et al., 2020), whereas 17 experiments found no effect in the particular region studied (Albuquerque et al., 2015; Hoefling et al., 2019; Leung et al., 2012; Loreth et al., 2012; Palop et al., 2007; Villette et al., 2012; Waller et al., 2020).
Summary of Somatostatin Experiments in Identified Literature
i.c.v., intracerebroventricular injection.
Six experiments studied SOM interneurons in postmortem human AD tissue. Five of these experiments used immunohistochemistry to quantify changes in SOM+ cell density in human AD tissue of Braak stage 4 or higher. Three experiments found a significant loss of SOM+ neurons in the CA1, DG, and subiculum (Sekiguchi et al., 2009), whereas two experiments found no change in SOM+ cell number in the CA1 and EC (Waller et al., 2020), compared with healthy controls postmortem. However, both these studies had small sample sizes of 6 (Sekiguchi, Habuchi, et al., 2009) and 10 (Waller et al., 2020). Finally, one experiment found increased pTau accumulation in SOM+ cells in postmortem human AD compared with controls (Zheng et al., 2020).
In studies that analyzed animal AD models, the majority of the experiments were in the CA1 region. Of these experiments, seven showed decreased SOM+ cells at time points between 6- and 18-months old in AD animals compared with controls (Aguado-Llera et al., 2018; Albuquerque et al., 2015; Levenga et al., 2013; Ramos et al., 2006; Shi et al., 2020; Villette et al., 2012), whereas two experiments showed no effect in AD animals compared with controls (Leung et al., 2012; Villette et al., 2012). Greater heterogeneity of results was found when assessing studies that analyzed other regions of the hippocampus. For example, four experiments assessed SOM immunoreactivity in the CA2 region of AD mice, two of which found decreased SOM immunoreactivity at 6 months (Albuquerque et al., 2015; Ramos et al., 2006), whereas two others observed no effect at various time points at 12 and 18 months, compared with controls; however, these two particular experiments assessed CA1–3 as a group and did not specifically assess CA2 separately (Loreth et al., 2012). Four experiments observed no change in SOM immunoreactivity in the CA3 region at 6, 12, and 18 months of AD animals compared with controls (Albuquerque et al., 2015; Leung et al., 2012; Loreth et al., 2012). One experiment found decreased SOM immunoreactivity in the CA1–3 regions (Ramos et al., 2006); however, the experiment did not specifically assess SOM+ levels in the CA3. In the hilus, four experiments found no change in SOM+ cell levels at various points between 3 and 18 months (Hoefling et al., 2019; Loreth et al., 2012; Palop et al., 2007), and one study found that only female apoE4-KI mice showed a decrease in SOM+ cell immunoreactivity compared with males and controls (Leung et al., 2012). In the subiculum, one article found no change in SOM immunoreactivity in a mouse AD model at 6 months compared with WT controls (Albuquerque et al., 2015). Two experiments assessed SOM+ levels in the EC, and one article found decreased levels in 4- to 7-month-old AD mice (Ramos et al., 2006), whereas the other found no effect at various points between 1- and 16-month-old apoE-knock-in mice compared with controls (Leung et al., 2012).
One article assessed SOM mRNA levels in APPxPS1 mice and found decreased levels starting at 4 months and onward. The same study found that APP mice had a nonsignificant reduction in SOM mRNA compared with WT controls (Ramos et al., 2006).
To assess the effect of tauopathy on SOM interneurons, 11 experiments that co-labeled SOM+ neurons with tauopathy markers in AD animal models were assessed. Five of these experiments demonstrated colocalization, or at least an increase in colocalization compared with controls (Levenga et al., 2013; Najm et al., 2020; Shi et al., 2020), two of which were specifically in the CA1 region (Levenga et al., 2013; Shi et al., 2020). Correspondingly, two experiments reported that this tau accumulation resulted in loss of SOM+ cells or SOM mRNA (Burgos-Ramos et al., 2007; Ramos et al., 2006). Four experiments found no colocalization in SOM+ interneurons in AD mice; one in the CA1 region (Soler et al., 2017), one in the CA3 region (Soler et al., 2017), and two in the DG (Levenga et al., 2013; Soler et al., 2017). Thus, the results demonstrate evidence that tauopathy does occur in SOM interneurons in AD (See also Supplementary Fig. S1).
We assessed one study evaluating the role of SOM interneurons in the genesis of hippocampal theta oscillations and found that they were impaired in three experiments for mice injected with soluble AβO at 28–49 days postnatal. However, these impairments were restored when optogenetically activated at theta frequency (Chunget al., 2020).
Finally, one article assessed the activity of SOM+ interneurons following chronic and acute doses of Aβ25–35 and found that there was a decrease of activity with a chronic 14 day infusion but no effect with a single dose of Aβ25–35 in male Wistar rats compared with controls (Burgos-Ramos et al., 2007).
Discussion
Cholecystokinin
Notably, few CCK studies were identified by this systematic review. In an APPNL-F/NL-F model, Aβ accumulation was seen at 9–18 months of age and that degeneration that cannot be attributed to age occurs at 9–18 months but not in 1- to 3-month-old mice (Shi et al., 2020). Another study in an APP knock-in model found that 32% of CCK-positive interneurons in the CA1 subfield of the hippocampus contained APP buildup and that 95% of APP was found in CCK-positive interneurons (Rice et al., 2020). An amyloid injection model shows no change in amyloid cell deposits or CCK-positive cell numbers in the CA1 at either 5 or 24 days postinjection (Villette et al., 2012). These data, taken with the other articles found, suggest that CCK interneurons are a site of APP accumulation later in disease pathogenesis but are heavily impacted and require more study. Interestingly, CCK-positive interneurons are located where many type 1 cannabinoid (CB1) receptors are found, and so these cells may mediate the effects of THC, the psychoactive component of cannabis, and AD (Pelkey et al., 2017). Also given the fact that CCK-positive interneurons degenerate later in the disease progression, treatments targeting these cells may be helpful in cases of early-onset AD.
Neuropeptide Y
Most studies found a decrease in the number of NPY-positive interneurons in AD models compared with controls (Aguado-Llera and Canelles, 2018; Albuquerque et al., 2015; Ramos Baglietto-Vargas, et al., 2006; Loreth et al., 2012; Mahar et al., 2017). Mahar and colleagues (2017) found a decrease in the numbers of these cells in mice as young as 1 month old. Two models that did not see a change in the number of NPY neurons were the amyloid precursor protein intracellular domain model used by Ghosal and Pimplikar (2011) and the Aβ injection model used by Villette and colleagues (2012). Villette and colleagues (2012) used an acute injection model, with the latest time point studied being 24 days postinjection. Figure 2 shows that the number of NPY-positive cells in the hilus decreases in all studies but one. The study that showed an increase was due to an ApoE4 knock in (Leung et al., 2012). Together, the time courses and robust agreement of these studies suggest that NPY-positive interneurons are heavily impacted by AD, but that they may not be the first interneuron type impacted. There is a paucity of studies that assess the impact of AD-related NPY neuron degeneration and NPYs affect on adult neurogenesis in the DG, and so this is an area ripe for future study. A subset of NPY-positive interneurons, called neurogliaform cells (caveat: these cells also express other interneuron markers), would be a particularly interesting subtype to study. This is because neurogliaform cells have a significant number of en passant boutons. In these boutons, GABA is released into the extracellular space—not the synaptic cleft—to activate metabotropic GABAA receptors and maintain a basal level of inhibition (Pelkey et al., 2017).
Parvalbumin
In every subsection of the hippocampal formation, there are conflicting results about whether the number of PV-positive interneuron is altered (Albuquerque et al., 2015; Aso et al., 2018; Loreth et al., 2012; Mahar et al., 2017; Petrache et al., 2019; Takahashi et al., 2010). There is evidence that AD affects inhibitory post-synaptic currents and gamma oscillations through PV interneurons and that inputs to PV interneurons in CA1 degenerate in AD (Chung et al., 2020; Park et al., 2020; Rubioet al., 2012).
In an hAPP-J20 mouse model, Palop and colleagues (2007) found aberrant innervation of PV interneurons by mossy fiber collaterals. Rice and colleagues (2020) found that in the stratum oriens of CA1, 41% of APP accumulation occurs in PV-positive interneurons. Villette and colleagues (2012) found that in their 24 day Aβ injection paradigm, a subset of PV/SOM interneurons was greatly affected by Aβ. Chung and colleagues (2020) also used an injection paradigm and found deficits in PV-positive interneurons that were rescued with optogenetic stimulation of PV interneurons. Increased PV-specific GABAergic input was found in 3-month-old APP/PS1 mice by Holnagel and colleagues (2019), whereas Hjazi and colleagues (2019) found an early increase in excitability in PV interneurons using whole-cell patch clamping in the same model. Verret and colleagues (2012) found a decrease in NaV.1 channels on PV interneurons.
In light of these findings, it would be pertinent for future research to assess PV interneurons as a potential early target for toxic APP products. This could lead to a loss of lateral inhibition, which could cause first early excitability and later, death, via toxic APP buildup and excitotoxicity. Further studies could be carried out to delineate if the decrease in NaV.1 levels found by Verret and colleagues (2012) is the cell's attempt to control aberrant activity, or a target of APP toxicity through affects on gephyrin. Interestingly, the articles that found no change in PV interneuron numbers were also models of tau and not APP accumulation. This agrees with findings that PV interneurons are affected early, and not late in AD. It would also be interesting for future studies to assess SOM and PV concurrently to see if the finding can be corroborated that the cell type that expresses both SOM and PV suffer more acutely. PV-positive cells in the DG are thought to participate in neurogenesis by controlling the integration of adult-born granule cells (abGCs) into the circuit; abGCs that do not receive inputs form PV-positive cells are much more likely to die (Song et al., 2013). As adult neurogenesis is a possible mechanism of memory, targeting these cells early in disease could be beneficial. Indeed, one group found that increasing the excitability of PV interneurons in early Alzheimer's in a mouse APP/PS1 model was beneficial (Hijazi et al., 2019).
Somatostatin
SOM-positive interneurons showed both APP and tau accumulation, although Rice and colleagues (2020) found minimal APP accumulation (Hoefling et al., 2019; Shi et al., 2020). There is disagreement as to whether the number of SOM-positive interneurons is lost or not; Albuquerque and colleagues (2015) found a decrease in the number of PV-positive cells in 6-month-old mice in the CA1/2 region and the stratum radium of CA3, whereas Loreth and colleagues (2012) found no change in the pooled CA1–3 regions, the DG, or the hilus in 12- or 18-month-old mice, although both articles used a tau accumulation model (Albuquerque et al., 2015; Sekiguchi et al., 2009). Similar phosphotau accumulation was found by Zheng and colleagues (2020) finding a robust accumulation in two separate mouse models, as well as human tissue, whereas Soler and colleagues (2017) found no co-labeling in any hippocampal subfield of tau phosphorylated at threonine 231. Villette and colleagues (2012) found no change in hippocampal amyloid deposits or in the number of SOM-positive interneurons after acute Aβ injections. Theta oscillations were shown to be affected by SOM-positive interneuron loss, and this phenomenon was confirmed with an optogenetic rescue experiment (Chung et al., 2020). SOM interneurons are integral in the production of theta oscillations and have more recently been implicated in lateral inhibition to maintain the fidelity of neuronal signals during memory retrieval (Stefanelli et al., 2016). The heterogeneity of findings about whether the number of SOM-positive interneurons increases, decreases, or remains unaffected suggests that more studies in this area should be performed and also that SOM-positive interneurons may themselves be heterogeneous and should be studied in conjunction with other intraneuronal markers. There is emerging evidence that the SST14 neuropeptide acts to exacerbate Aβ aggregation, and so the heterogeneity of results could be due to the time it takes for this event to happen in different AD models (Solarski et al., 2018). SOM-positive interneuron degeneration in the hilus has previously been shown to be implicated in nonpathological age-related memory changes, and further treatments with a low dose of the antiepileptic drug levetiracetam were able to upregulate SOM expression (Spiegel et al., 2013). It is important to note that SOM is a peptide hormone that binds to a host of G-protein-coupled receptors in the brain and so changes in the level of this peptide hormone as well as loss of GABA signaling by interneurons positive for this marker must be considered in future studies (Pelkey et al., 2017).
Conclusions
This systematic review found that there is strong evidence that PV interneurons are affected early in the disease by toxic Aβ fragments. This leads to a loss of lateral inhibition and increased excitotoxicity. The loss of lateral inhibition will likewise affect SOM interneurons indirectly while the SOM neuropeptide may act to slowly worsen toxic Aβ fragment accumulation. The literature suggests that NPY- and CCK-positive interneurons are affected later in the progression of the disease and lead to loss of regulation of adult neurogenesis and the cannabinoid system, respectively. In the future, treatments for AD may fall into two categories: treatment for those who are diagnosed early and those who are diagnosed only when symptomatic. Treatments that upregulate PV and SOM activity may buy patients some time before lateral inhibition breaks down and memory fidelity is lost. Treatments that target CCK- and NPY-positive interneurons may be helpful in the late disease where the SOM and PV interneurons have already degraded. There is also a possibility that targeting CCK interneurons may increase overall inhibition through the CB1 pathway, by disinhibiting GABA release. This systematic review aims to identify gaps in the literature and opportunities for new experiments to strengthen our understanding in the field.
Footnotes
Authors' Contributions
H.M.O.R. and N.C.-M. were responsible for the conceptualization, reference selection, and writing of this article. T.S. was responsible for checking the applicability of each included reference, as well as editing the text and designing some figures. B.R.C. was responsible for conceptualization, writing of the article, and editing of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by a CIHR catalyst grant (FRN:163015).
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.