
Abstract
Background:
The endogenous allocation of spatial attention to selected environmental stimuli is controlled by prefrontal (frontal eye fields [FEFs]) and parietal (superior parietal lobe [SPL] and intraparietal sulcus [IPS]) regions belonging to the dorsal attention network (DAN) with a subdivision in subsystems devoted to reorienting (or shifting) of attention between locations (SPL) or maintaining attention at contralateral versus ipsilateral locations (ventral IPS [vIPS]). Although previous studies suggested a leading role of prefrontal regions over parietal sites in orienting attention, the spectral signature of communication flow within the DAN for different attention processes is still debated.
Methods:
We used the directed transfer function (DTF) on magnetoencephalography (MEG) data to examine the causal interaction between prefrontal and parietal regions of the DAN when subjects shifted versus maintained attention to a stream of cued visual stimuli.
Results:
In the beta band, we found that shift versus stay cues induced stronger connectivity (DTF values) from right FEF to right SPL, in the early phase of reorienting. Conversely, when considering stay versus shift cues, an increase of DTF values and stronger directionality was observed between bilateral vIPS and from right vIPS to FEF. Similar analyses carried out in theta, alpha, and gamma showed no significant frontoparietal increases of DTF for shift versus stay cues, whereas the stay-related increase of DTF observed in beta between ventral parietal areas was preserved in the alpha band.
Conclusions:
These findings suggest that control processes in DAN regions (in particular between FEF and SPL) can be associated to a beta frequency channel during shift of attention.
Impact statement
In the present study, we compared the reorienting response to novel stimuli with respect to maintaining response. Results provided new insights into understanding the neural mechanisms of control attention processes by identifying the frequency-specific causal interactions between frontal and parietal regions belonging to the dorsal attention network supporting spatial reorienting response.
Introduction
Humans can rapidly process a visual scene focusing on the most relevant aspects. It has been widely reported that the allocation of spatial attention to selected environmental stimuli is controlled by prefrontal (i.e., frontal eye fields [FEFs]) and parietal (i.e., superior parietal lobe [SPL] and intraparietal sulcus [IPS]) regions forming the so-called dorsal attention network (DAN) (Corbetta and Shulman, 2002). Interestingly, within the parietal regions of the DAN, a functional-anatomical segregation has been demonstrated between medial regions in the SPL, which encode transient signals for shifting attention between spatial locations, and regions of the posterior ventral IPS (vIPS), which encode sustained spatially selective signals for maintaining attention at contralateral versus ipsilateral locations (Shulman et al., 2009; Tosoni et al., 2013). Consistently, transcranial magnetic stimulation (TMS) studies provided causal evidence for this subdivision of the DAN into subsystems dedicated to either shifting or maintaining covert visuospatial attention (Capotosto et al., 2013, 2015). Specifically, interference of the right vIPS, but not right SPL, was shown to impair target discrimination at contralateral locations, whereas stimulation over the right SPL, but not vIPS, affected target discrimination following a shift of attention irrespective of its location. In addition, a key role of the FEF has been observed in nonhuman primates during shifts of covert attention (Buschman and Miller, 2009), and in humans during the preparatory period of a visuospatial attention task (Siegel et al., 2008), specifically through a mechanism of increased coherence in the beta frequency band. Taken together, the above results suggest a crucial role of prefrontal (FEF) and medial parietal (SPL) regions of the DAN for shifting attention, and of more ventral parietal regions (vIPS) of the DAN for the maintenance of attention. A recent magnetoencephalography (MEG) study provided evidence that frontoparietal regions interact in alpha and beta bands during top-down (task-driven) attention (D'Andrea et al., 2019). This interaction may be directional, for example, from frontal to parietal areas or vice versa. fMRI findings showed an increased communication characterized by time series of bilateral FEF that predict signals of posterior parietal and visual occipital regions during top-down attention (Bressler et al., 2008; Spadone et al., 2015). In the same framework, animal studies suggested a spectral specificity of the directional interaction related to respective task demands. Specifically, stronger top-down attention involves increased beta coherence from frontal to parietal regions, whereas bottom-up (sensory driven) attention involves increased gamma coherence from parietal to frontal regions (Buschman and Miller, 2007). A similar association of lower frequency bands with top-down and higher frequency bands with bottom-up signaling has also been found in the visual occipital system (Bastos et al., 2015; van Kerkoerle et al., 2014). Notably, a consistent interareal directional influence within the visual system has been also reported in humans (Michalareas et al., 2016), although to date a directional investigation between frontoparietal regions during top-down attention is still to be elucidated.
In the present study, we assume that based on animal versus human consistency observed in the alpha/beta band in the visual system during visuospatial attention, beta band specificity of top-down communication reported in primates will be found also in humans. At this aim, we focused on different top-down attentional processes (i.e., maintaining or shifting) using a paradigm that emphasized the endogenous signals. We explored the directionality in the beta band (15–30 Hz) while controlling for directionality in the other frequency bands in different functional subdivisions of the DAN, specifically between subsystems involved in shifting (dorsal [d]FEF, SPL) versus maintaining (vIPS) attention. To this aim, we applied the directed transfer function (DTF), a directional connectivity method based on Granger causality approach, to estimate the causal influences between MEG source activities of selected brain regions. Based on the above observations in animal studies (Buschman and Miller, 2007) and in humans (Bressler et al., 2008; Capotosto et al., 2013), we hypothesize a stronger directed functional connectivity in the beta band from the dFEF to the SPL compared with vIPS for the shifting of attention. In addition, we predict a stronger information flow involving more ventral parietal nodes (i.e., bilateral vIPS) during the maintenance of attention.
Materials and Methods
Subject and stimuli
Twenty healthy subjects participated in the study after providing written informed consent according to the Ethics Committee at the University of Chieti (July 22, 2008; HEALTH-F2-2008-200728). All the subjects participated in our previous combined fMRI study, and a subset participated in the EEG-TMS study (Capotosto et al., 2013, 2015; Spadone et al., 2015). During the procedure, they were asked to maintain fixation on a central cross while covertly directing attention to one of the two drifting Gabor patches constantly presented on the left and right peripheral locations, with the following parameters: 2 cycles/° spatial frequency, 0.7°/sec drift rate, 3° diameter, 5.5° eccentricity from central fixation. The to-be-attended location was indicated by the appearance of a cue consisting of a 300 ms isoluminant color change of both patches (pink, cyan), of which only one indicated the attended stream (e.g., cyan, Fig. 1A) that was indicated at the beginning of the run and counterbalanced across runs. Relevant color could appear in the same location with respect to the previous one, indicating to maintain attention to the currently attended stream of stimuli (stay cue), or in the opposite stream, indicating to reorient attention (shift cue). Cue location correctly predicted with 80% probability where (but not when) targets would appear, consisting of a brief clockwise or counterclockwise change in the orientation of one patch. Nevertheless, a target could not occur earlier than 1 sec after a cue, and cues could not occur earlier than 2 s after a target. Subjects were instructed to detect and discriminate orientation changes as fast as possible by pressing a button on a response box with their right hand.
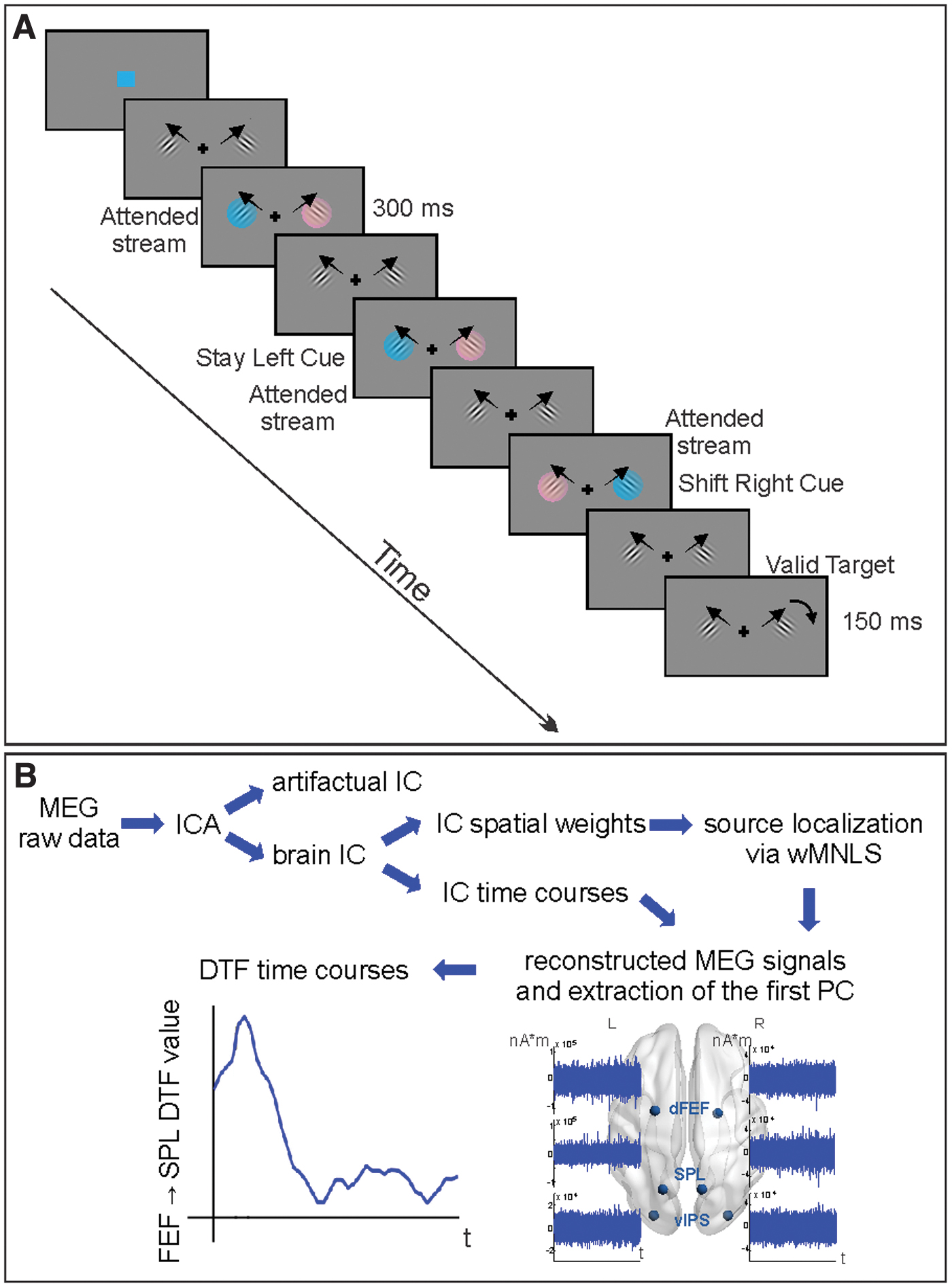
Example of the display sequence of the experimental paradigm
Subjects were selected through a preliminary behavioral session in which performance and eye position (Iscan etl-400, RK-826 PCI) were evaluated. The selected subjects showed a significant validity effect (valid>invalid) on target discrimination accuracy and were able to maintain central fixation after both stay and shift cues [detail can be found in Spadone and colleagues (2015); supplementary materials and methods section].
MEG procedure, preprocessing, and regions of interest
MEG signals were recorded using the 153-channel MEG system installed in a magnetically shielded room at the University of Chieti, Italy (Della Penna et al., 2000). Magnetic fields were continuously acquired at 1025 Hz sampling rate while participants performed the visuospatial attention tasks and the original signals were filtered in the band 1–150 Hz. Electrocardiogram and electrooculogram were simultaneously recorded for offline artifact rejection and to monitor horizontal eye movements. Participants underwent an MEG session including two runs of about 16 min of the visuospatial attention task. Individual high-resolution anatomical images (M-PRAGE T1-weighted sequence) recorded during the fMRI study (Spadone et al., 2015) were used for MEG localization. Two subjects were excluded from further analyses in the preprocessing step, due to abnormal artifacts and to technical problems. Then, the magnetic signals were analyzed using independent component analysis to separate artifact components from the ones containing the brain activity (Mantini et al., 2011; Spadone et al., 2012). The sensor maps of the latter ones were projected into the source space through a weighted minimum-norm least-squares procedure implemented in Curry 6.0 (Neuroscan). This step was performed using the coregistered individual anatomical images to obtain a realistic model of the source space and volume conductor sampled by a Cartesian 3D grid with 4 mm voxel side. Source space signals were reconstructed by combining time courses of non-artifactual components weighted by the related source maps, normalized to the Montreal Neurological Institute brain template (MNI152). Specifically, we analyzed source-space signals of the voxels from the regular grid corresponding to centroids of fMRI regions of interest (ROIs). In this work, the source space for both modalities is represented by a 3D grid with different voxel sizes (BOLD—3 mm, MEG—4 mm). While this size reflects the effective resolution of the fMRI method, the effective MEG spatial resolution is larger than 4 mm. However, these methodologies observe distinct phenomena (i.e., the vascular response in fMRI, the electrophysiological activity in MEG) and the location of the vascular response could be displaced with respect to the true activity. Moreover, possible effects of modeling approximations in the MEG source estimation increase the inherent spread of MEG sources. Taking into account all these effects, when seeding MEG sources from BOLD results, spheres with radius of 3 mm (and even larger) centered in the centroid of the fMRI ROI can be associated with MEG sources (Torquati et al., 2005), representing a compromise accounting for the different spatial resolutions of the two techniques and the inherent source displacement in fMRI. These considerations suggest that using the MEG voxel activities with the closest positions to the centroid of the fMRI ROIs is consistent with previous works associating MEG activity with fMRI ROIs (Del Gratta et al., 2002; Torquati et al., 2005). Source-space signals were extracted for ROIs belonging to the DAN, selected from shift-related (shift>stay) and spatially selective (contralateral>ipsilateral) evoked responses during cue in our previous fMRI study (Spadone et al., 2015) run on the same subjects using the same stimuli. Briefly, this selection was obtained through a whole-brain ANOVA with cue type (shift and stay), cue location (right and left), and time (seven time points) as factors. Stronger shift-related responses, regardless of cue location, were identified from the Cue Type by Time map in the SPL and in the dorsal aspect of the FEF (Fig. 2). Stronger preference for cue stimuli presented in the contralateral hemifield, regardless of cue type, was identified from the Cue Location by Time map in the vIPS (Fig. 2). Importantly, we selected, among the other visual and DAN regions, the dFEF, which showed a leading role in visuospatial attention (Bressler et al., 2008; Spadone et al., 2015) and two parietal regions, which, respectively, showed a selective causal role for shifting attention between locations and attending to contralateral locations, as confirmed by our repetitive transcranial magnetic stimulation studies (Capotosto et al., 2013, 2015). For further analyses, we considered the first principal component of the vector activity of each region. For each cue type, the phase-locked activity was rejected through an adaptive algorithm, regressing out the average evoked signal from each trial (Della Penna et al., 2004).
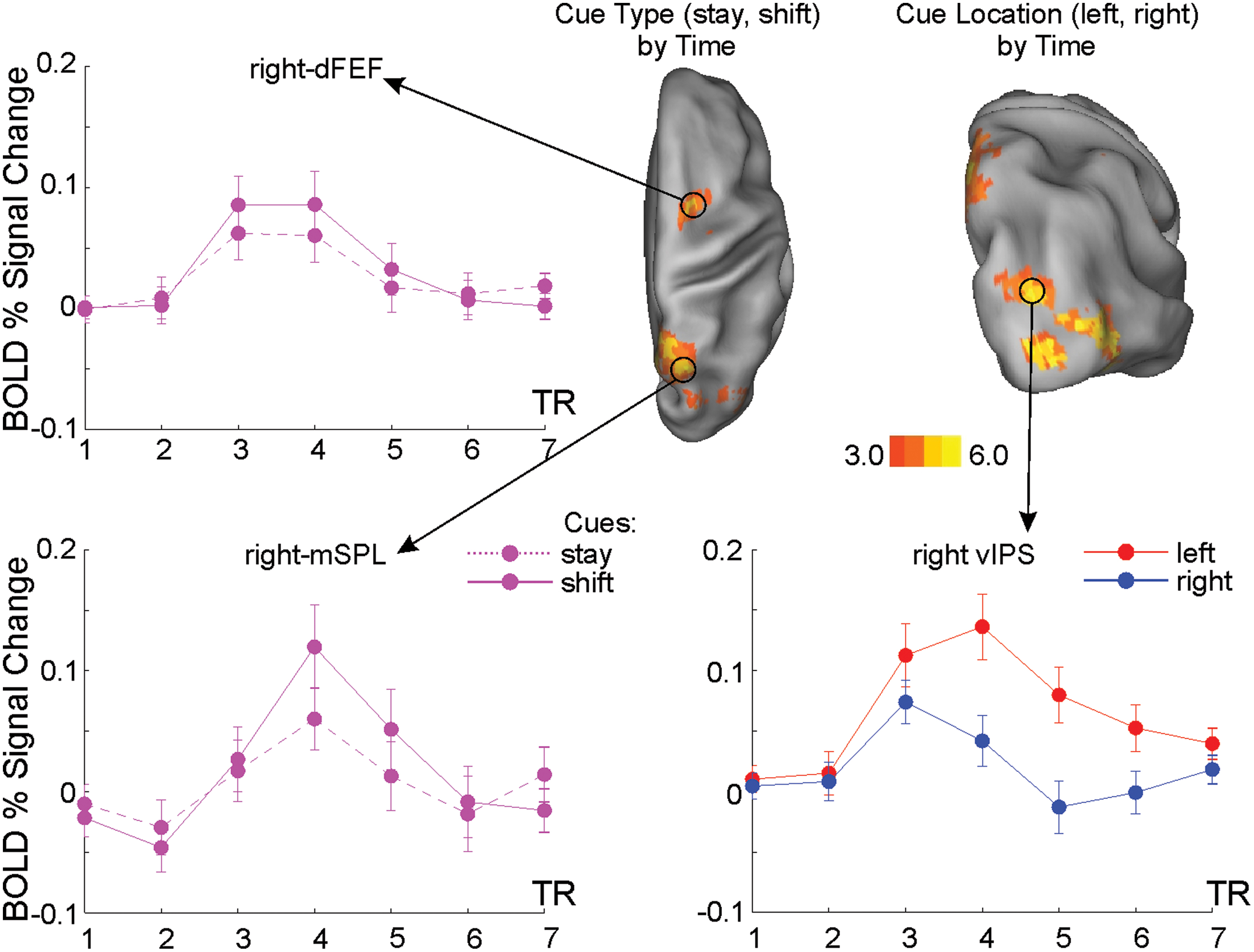
fMRI results. Voxels showing significantly group-wise different fMRI activation after shift versus stay cues (cue type by time map), and left versus right cues (cue location by time map), respectively (top-right part of the figure). Time courses of the BOLD response extracted from peak regions in right dFEF (top left), mSPL (bottom left), and vIPS (bottom right) where error bars represent SE across subjects. In figure were only reported fMRI activations and relative time courses for the right hemisphere. Figure is derived from the results of our previous fMRI publication (Spadone et al., 2015). dFEF, dorsal frontal eye field; mSPL, medial superior parietal lobe; vIPS, ventral intraparietal sulcus. Color images are available online.
DTF analysis and statistical analysis
The task-induced modulations of directed functional connectivity between the selected ROIs were analyzed in postcue periods by means of the DTF (Blinowska et al., 2004; Kaminski et al., 2001). Based on the Granger concept, this method provides a measure of causal relationships between signals in the frequency domain using multivariate autoregressive modeling (MVAR).
Before fitting the MVAR model and improving its performance, we filtered frequencies outside the bands of interest using a separate Chebyshev type II filter (4–50 Hz). Furthermore, we also compensated for spatial leakage confounds using a symmetric multivariate orthogonalization procedure (Colclough et al., 2015). Then the signal was resampled to 128 Hz and epoched to segments 0–800 ms relative to cue onset. Following this step, an average number of trials (N = 175 ± 6) per condition (i.e., stay and shift) were available for the connectivity analysis.
All DTF calculations were carried out with Multar toolbox (University of Warsaw). The MVAR model order was set to 8, according to the Akaike information criterion. DTF was estimated in the beta band (15–30 Hz) according to our hypotheses. Moreover, the theta (4–8 Hz), alpha (8–13 Hz), and gamma (30–50 Hz) information transfers were used as a control for frequency specificity. Non-normalized DTF values were calculated for selected directions in sliding windows of 150 ms duration shifted by 20 ms, thus resulting in N = 33 bins with the last starting from 640 to 790 ms (of note, the last part of the trial intervals that could be contaminated by target occurrence was not analyzed). The distributions of resulting individual subject connectivity values were checked for possible extremes, defined using boxplot 1.5 IQR (interquartile range). Then, individual DTF time series were smoothed by symmetrical moving of the mean with the window width set to three data points, which was a good compromise between high temporal resolution and reduction of random variations (see Fig. 1B for the flowchart of the method).
Statistical analysis was performed using linear mixed models with the lme4 R toolbox (Bates et al., 2015). For each direction, a single statistical model was built, which included two fixed factors and their possible interactions (shift/stay condition, time window) and subjects as a random factor. p Values of condition effects at each sliding window were determined using the Satterthwaite's method for approximating degrees of freedom by means of the lmerTest library (Kuznetsova et al., 2017), and then corrected by the number of pairs of interest (N = 12 as showed in Fig. 3) with the Bonferroni method.
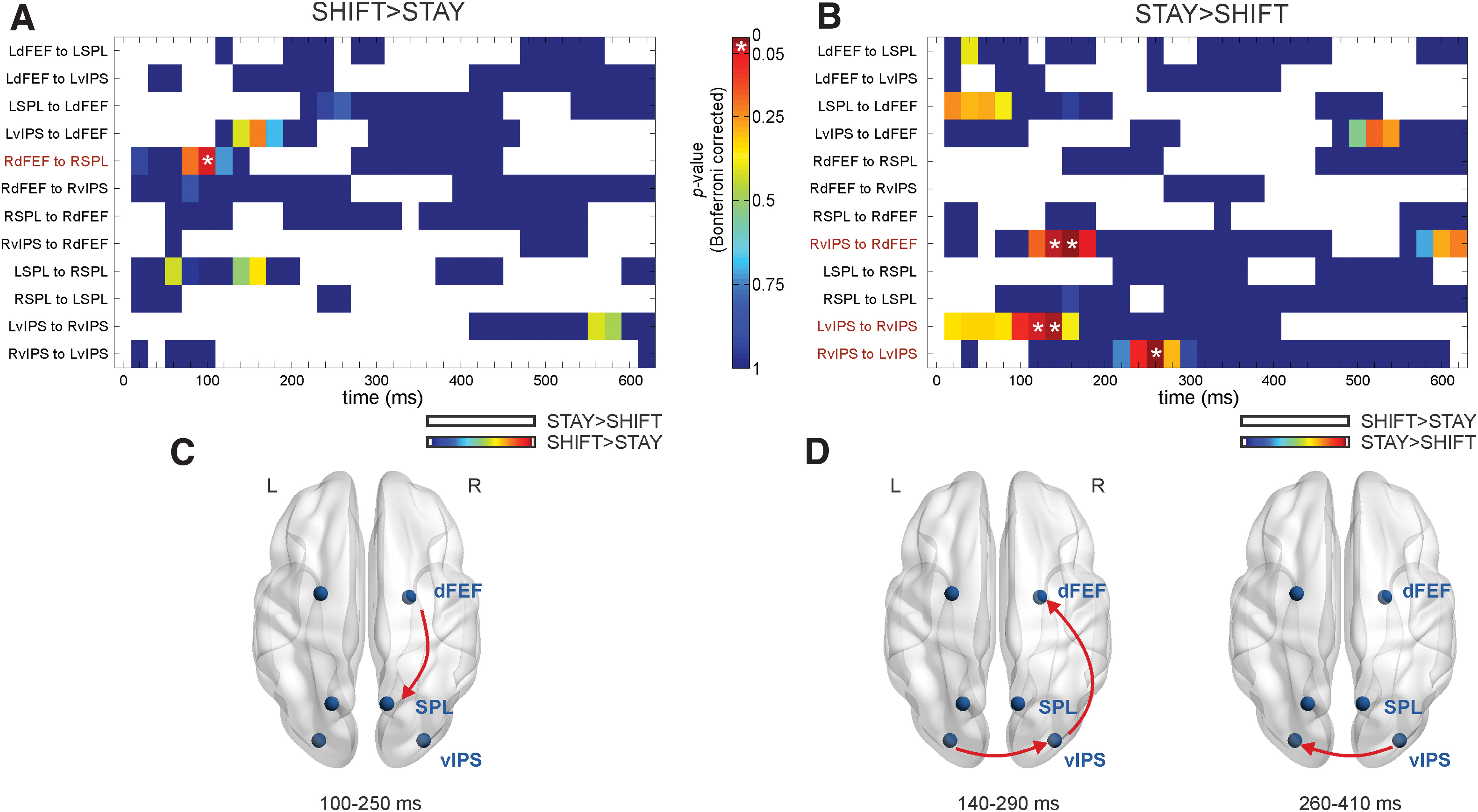
DTF results. p Values of DTF analysis for shift>stay
Eye movement
To further verify that the directional connectivity results were not guided by eye movements, we checked the correlation between eye movement parameters and DTF results during epochs showing significant modulations of directional communication. Specifically, we analyzed eye movements using vertical and horizontal electro-oculography (EOG) signals simultaneously recorded with MEG signals. First, EOG signals were converted in degree of visual angle using the calibration factor obtained from calibration runs recorded at the beginning and at the end of each visuospatial attention run. Then we obtained event-related time courses of horizontal eye position time-locked to the presentation of stay and shift cues, averaged across trials within each subject. For a better estimation of eye movements from the EOG, we removed the baseline drift and the high-frequency noise (Bulling et al., 2011) and the intervals across eye-blinks, which were identified from the vertical EOG. For each subject, the horizontal eye position time courses, locked to the presentation of cues, were estimated during four consecutive 200 ms time bins (800 ms total) following the cue onset. From these signals, the averaged values and the peaks of eye movements were estimated and correlated with DTF results during epochs showing significant modulations of directional communication across subjects.
Results
Shift- versus stay-related modulations of DTF in beta band
We examined whether different attentional processes are mediated by selective dynamics of directional interactions between prefrontal (dFEF) and parietal (SPL and vIPS) regions of the DAN, in the beta band. This hypothesis was examined by contrasting, for all possible ROI pairs, modulations of directed functional connectivity, over time bins during epochs of shifting versus maintaining of attention and vice versa. Epochs showing significant increases of directional communication following shift versus stay cues and vice versa, after Bonferroni correction, are displayed in Figure 3A and B, respectively. At these epochs, both stay- and shift-related increases of DTF values in the beta band were represented as arrows on standard brain in axial plane (visualized with BrainNet Viewer) (Xia et al., 2013) (Fig. 3C, D).
We observed a significant shift-related increase of DTF (shift vs. stay, Fig. 3A–C) only for the right dFEF-right SPL pair. This increase was observed in the time window 100 ÷ 250 ms following spatial cues (p < 0.05, Bonferroni corrected). In contrast, when considering the stay-related increase of DTF values (i.e., stay versus shift, Fig. 3B–D), the frontoparietal connection was replaced by a stronger link between bilateral vIPS. The initial higher directional connectivity reported from left to right vIPS in the time windows 120 ÷ 270 and 140 ÷ 290 ms (ps < 0.05, Bonferroni corrected) was followed by the reversed connection in 260 ÷ 410 ms (p < 0.05, Bonferroni corrected). Moreover, we reported a strong directionality from right vIPS to right dFEF in two subsequent periods, 140 ÷ 290 and 160 ÷ 310 ms (ps < 0.05, Bonferroni corrected).
Control analyses on frequency specificity of shift- and stay-related increases of DTF and eye movements
We further tested whether these selective modulations of directional interactions were specific for the beta band. To this aim, we repeated the analysis in the other frequency bands (theta, alpha, and gamma) on the ROI pairs showing significant modulations in the beta band (i.e., from right dFEF to right SPL, between bilateral vIPS in both directions, and from right vIPS to right dFEF). Both the shift- and stay-related increases of DTF are displayed in Figure 4 as a function of time and frequency. Results showed no significant increases of DTF between prefrontal and parietal regions after shift compared with stay cues in the other frequency bands (Fig. 4A), whereas communication between ventral parietal areas was partially preserved in the alpha band, that is, only from left to right vIPS in the time window 120 ÷ 270 ms (p = 0.05, Bonferroni corrected), when contrasting stay versus shift cues (Fig. 4B–D). Furthermore, to verify that the directional connectivity results were not guided by eye movements, we checked the correlation between eye movement parameters and DTF results for region pairs showing significant beta effects during the corresponding time epochs. Importantly, our DTF results were not significantly (or close to be significantly) associated with eye movements (all ps > 0.4).
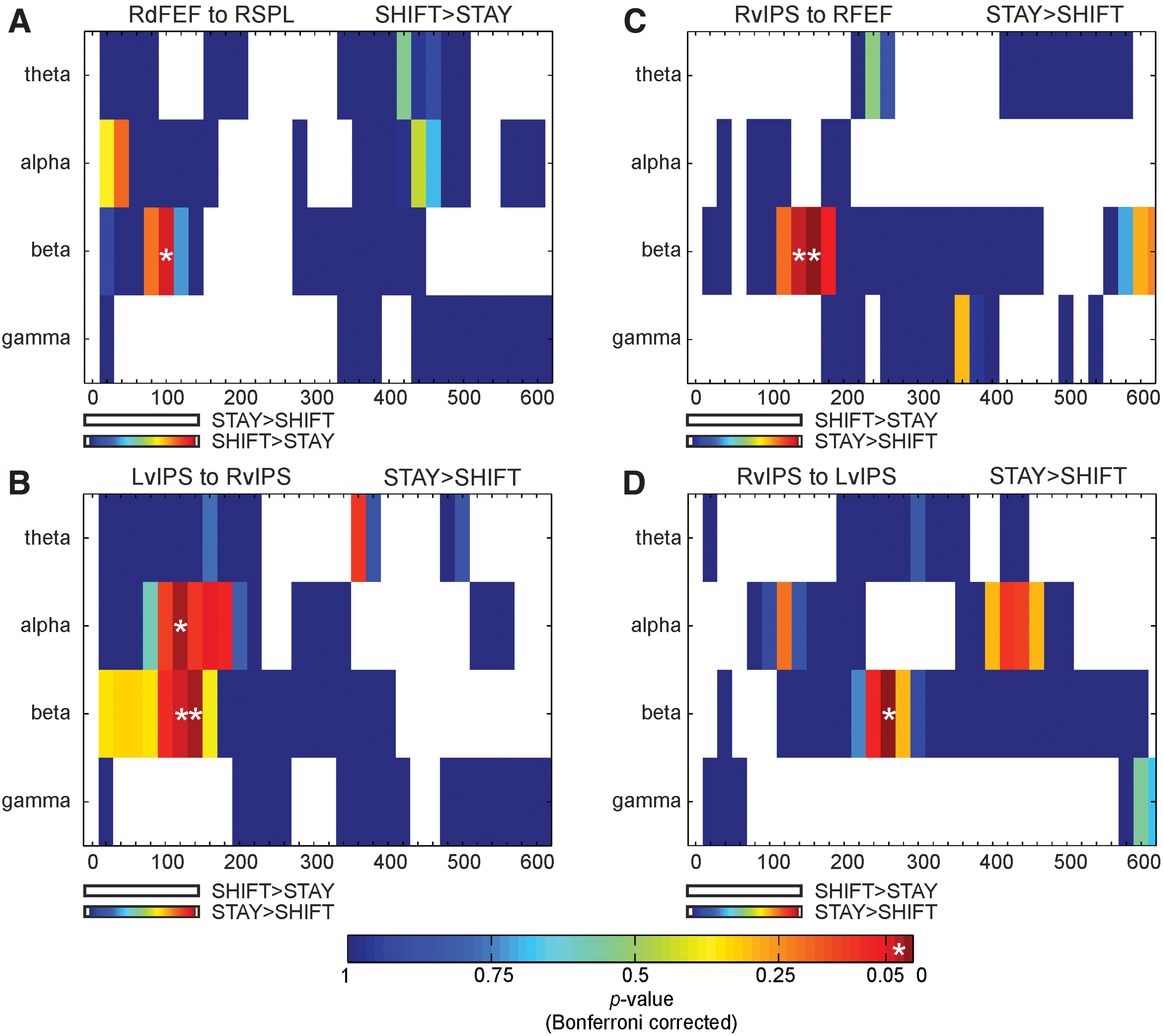
p Values of DTF analysis as a function of time and frequency for ROI pairs showing significant modulations in the beta band [from right dFEF to right SPL
Discussion
The present MEG study examined the modulation of the directed flow of the beta band communication between prefrontal (FEF) and parietal (vIPS and SPL) regions belonging to the DAN when comparing shifting versus maintaining of attention. The results indicate a stronger information flow from the right dorsal FEF to right SPL after shift with respect to stay in the early phase of the attentional process. On the contrary, when contrasting stay with shift cues, a strong directionality was reported between bilateral vIPS and from the right vIPS to right dFEF. Notably, while the shift-related increase of DTF from dFEF to SPL was specific for the beta band, the stay-related increase of DTF involving vIPS was observed in the beta and, partially, in the alpha band.
This is the first electrophysiological evidence showing that following a shift cue, the top-down control of reorienting of attention was exerted by the prefrontal DAN region and directed toward (medial) parietal regions using the beta band communication. As a matter of fact, during voluntary attentional control, two contrasting models have been proposed. Specifically, one model suggested that activities in parietal regions precede activity in frontal area (Green and McDonald, 2008). On the contrary, an alternative model argued that frontal regions initiate the sequence of attentional operations (Bressler et al., 2008; Spadone et al., 2015). Our results partially support the latter one. In fact, the frontal-to-parietal control was restricted to the medial part of the parietal cortex (i.e., SPL), did not involve the ventral part of the parietal cortex, and was exerted mainly for the shift of attention. Such regional selectivity is in line with several neuroimaging studies associating a direct involvement of both FEF and SPL for the shifting of attention (Capotosto et al., 2013, 2015; Shulman et al., 2009; Spadone et al., 2015; Tosoni et al., 2013). Accordingly, TMS studies assigned a causal role for attention shifting between locations to both SPL (Capotosto et al., 2013, 2015) and FEF (Grosbras and Paus, 2002). Interestingly, Grosbras and Paus (2002) showed that TMS interfered with shift of attention only in the case of right hemisphere stimulation, consistent with the present results showing an asymmetry of the directional connectivity modulations during shift versus maintenance of attention, thus providing further evidence for the dominance of the right hemisphere during this process (Heinen et al., 2017).
Here, we focused on the beta band since in literature it is associated with top-down attention (Bastos et al., 2015). Importantly, the control analysis of frequency specificity revealed that the present shift-related frontoparietal (i.e., from dFEF to SPL) connectivity was observed in beta but not in the other frequency bands. The key role of the beta band within the FEF is in line with the idea that top-down control of spatial attention is generally mediated by beta band synchronization (Buschman and Miller, 2007; Richter et al., 2017). Taking together these findings, we argue that the beta frequency is the communication channel between the prefrontal and medial parietal DAN regions. It is used for the actual orienting of the current focus of attention toward the new stream, promoting novel (sensory) representations before the target onset.
Another interesting result of the present study is the key role of ventral IPS during the maintenance of attention. As a matter of fact, we showed a stronger information flow from vIPS to FEF (in the beta band) and between bilateral vIPS (in both alpha and beta bands), when considering the stay-related increase of DTF. This is in accordance with several EEG/MEG studies suggesting that alpha/beta modulations are observed in the parieto-occipital cortex during the anticipation of forthcoming stimuli (Bauer et al., 2014; Capotosto et al., 2017; Spadone et al., 2017, 2020), for example, in the delay period before target onset (Bauer et al., 2014; Thut et al., 2006). Furthermore, at least for the alpha band, the present finding is in line with our previous EEG-TMS study, using the same paradigm (Capotosto et al., 2015), showing that interference with vIPS altered the behavioral performance and the peak latency of desynchronization in the alpha band in parieto-occipital regions, following stay compared with shift cues. In contrast to shift modulations, the stay-related increases of DTF between ventral parietal regions of the DAN show a bidirectional boost of information flow, especially in the beta band.
Notably, since we were explicitly interested in modulations of directional connectivity induced by top-down attentional processes within DAN, we restricted the analysis to few fMRI-defined ROIs, which showed a selective modulation of BOLD activity and a distinct causal role for shifting and contralateral maintaining of visuospatial attention. However, we suggest that future studies should investigate whether a third region, which was not considered in our analysis, may control the observed phenomena.
Conclusions
Overall, these results indicate that reorienting versus maintaining of attention increases the top-down frontoparietal directional connectivity in beta, but not in alpha band. These results corroborate the view that control processes in the DAN, exerted by the prefrontal to parietal region during attentional reorienting, are to be conveyed through a beta frequency channel. Conversely, both alpha and beta bands showed an increase of directional interaction between ventral parietal nodes of the DAN during maintaining versus reorienting of attention.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the “Departments of Excellence 2018–2022” initiative of the Italian Ministry of Education, University and Research for the Department of Neuroscience, Imaging and Clinical Sciences (DNISC) of the University of Chieti-Pescara and by the National Science Center, Poland (Grant No. 2016/21/B/HS6/03723).