
Abstract
Background:
Neuroimaging studies suggest that aged brains show altered connectivity within and across functional networks. Similar changes in functional network integrity are also linked to the accumulation of pathological proteins in the brain, such as amyloid-beta plaques and neurofibrillary tau tangles seen in Alzheimer's disease. However, less is known about the specific impacts of amyloid and tau on functional network connectivity in cognitively normal older adults who harbor these proteins.
Methods:
We briefly summarize recent neuroimaging studies of aging and then thoroughly review positron emission tomography and functional magnetic resonance imaging studies measuring the relationship between amyloid–tau pathology and functional connectivity in cognitively normal older individuals.
Results:
The literature overall suggests that amyloid-positive older individuals show minor cognitive dysfunction and aberrant default mode network connectivity compared with amyloid-negative individuals. Tau, however, is more closely associated with network hypoconnectivity and poorer cognition. Those with substantial amyloid and tau experience even greater cognitive decline compared with those with primarily amyloid or tau, suggesting a potential interaction. Multimodal neuroimaging studies suggest that older adults with pathological protein deposits show amyloid-related hyperconnectivity and tau-related hypoconnectivity in multiple functional networks, including the default mode and frontoparietal networks.
Discussion:
We propose an updated model considering the effects of amyloid and tau on functional connectivity in older individuals. Large, longitudinal neuroimaging studies with multiple levels of analysis are required to obtain a deeper understanding of the dynamic relationship between pathological protein accumulation and functional connectivity changes, as amyloid- and tau-induced connectivity alterations may have critical and time-varying effects on neurodegeneration and cognitive decline.
Impact statement
Amyloid and tau accumulation have been linked with altered functional connectivity in cognitively normal older adults. This review synthesized recent functional imaging literatures in a discussion of how amyloid and tau can interactively affect functional connectivity in nonlinear ways, which can explain previous conflicting findings. Changes in connectivity strength may depend on the accumulation of both amyloid and tau, and their integrative effects seem to have critical consequences on cognition. Elucidating the effects of these pathological proteins on brain functioning is paramount to understand the etiology of Alzheimer's disease and the aging process overall.
Introduction
The aging brain displays progressive changes over time that can lead to functional and cognitive decline, ranging from cellular and molecular changes (Freeman et al., 2008; Pakkenberg et al., 2003; Scheibel et al., 1975) to gross anatomical atrophy (Dekaban, 1978; Hartmann et al., 1994) to large-scale functional network disruption (Geerligs et al., 2015; Grady et al., 2016). The prefrontal cortices and medial temporal lobe (MTL) are especially susceptible to age-related changes, as they show marked atrophy with age as well as altered functional connectivity, which are closely linked to cognitive decline (Damoiseaux et al., 2016; Raz et al., 2004).
While those with advanced neurodegenerative diseases show marked reductions in brain function, the border separating normal and pathological brain aging is blurred; the accumulation of pathological proteins, such as amyloid-beta and tau, can cause abnormal brain function even among older adults with seemingly normal cognition. The accumulation of amyloid-beta and tau protein deposits, accompanied by age-related neuroanatomical and cognitive decline, is thought to have important consequences in the development of dementia (see reviews by Heutink, 2000; Rowe et al., 2007).
Functional magnetic resonance imaging (fMRI) has emerged as a useful tool to measure regional brain activity and functional network connectivity, and it is becoming more commonly applied to study the aging brain and its relation to cognitive decline. Older adults develop cognitive deficits with advancing age, presenting difficulties with executive function, processing speed, and episodic memory formation, while semantic and verbal memory are relatively preserved with age (Hedden and Gabriel, 2004; Warner Schaie, 1996). Both decreases and increases in brain activation have been observed and respectively related to impaired cognitive ability and compensatory effects.
The relationship between age-related changes in brain function and cognition is clearly complex and is likely affected by neuropathologies, such as amyloid and tau accumulation. Critically, older adults can harbor amyloid and/or tau and remain cognitively normal (Crary et al., 2014; Jansen et al., 2015; Oh et al., 2014), although the effects of these pathological proteins on brain function during the normal course of aging are not well understood. Previous models have suggested that increasing amyloid accumulation is associated with increasing and then decreasing brain activity (Foster et al., 2018; Jagust and Mormino, 2011), although recent evidence suggests that decreasing brain activity and connectivity may be more closely associated with tau levels.
In this review, we first briefly summarize the recent literature on age-related changes in brain activation and functional connectivity and then explore the effects of amyloid and tau on functional connectivity. We propose an updated model suggesting a period of amyloid-induced hyperconnectivity, followed by tau-induced hypoconnectivity in older individuals.
Neuroimaging of Aging
Various functional neuroimaging methods are used to study changes in brain activity and functional connectivity with age. Changes in blood-oxygen-level-dependent signals are detected and measured with fMRI, which can be used to record brain activity while a subject is at rest or completing a cognitive or behavioral task.
Here, we focus on the frontoparietal network (FPN) and the default mode network (DMN) (Yeo et al., 2011, but see Dixon et al., 2018), because these networks show well-documented age-related changes and are most commonly studied (Avelar-Pereira et al., 2017; Campbell et al., 2012; Chan et al., 2014; Geerligs et al., 2015; Grady et al., 2016, and see reviews by Grady, 2012; Heckner et al., 2020). The FPN is a task-positive network primarily composed of the dorsolateral prefrontal cortex and posterior parietal cortex (Duncan, 2010), and the DMN is a task-negative network primarily composed of the medial prefrontal cortex, angular gyrus, posterior cingulate cortex, and precuneus (Raichle, 2015). Both age-related increases and decreases in activity and connectivity have been observed among healthy older adults, which also vary by performance and task demand.
This section will focus on age-related changes in brain activity and functional connectivity changes in cognitively normal older adults. We have summarized some of the main findings, but refer to previous reviews for more in-depth discussions (e.g., Heckner et al., 2020; Sala-Llonch et al., 2015; Stern et al., 2019).
FPN changes with age
In neuroimaging studies of healthy young adults, activations in the dorsolateral prefrontal and posterior parietal cortices are commonly observed during attention/executive function tasks requiring goal-directed cognitive control of behavior (see reviews by Corbetta and Shulman, 2002; Duncan, 2010). Neuroimaging studies of older adults have shown altered frontoparietal activation in association with their task performance level, as well as changes in both within-network and between-network functional connectivity (Avelar-Pereira et al., 2017; Gallen et al., 2016; Geerligs et al., 2014; Grady et al., 2016; Turner and Spreng, 2015).
A common observation in aging fMRI studies is greater lateral prefrontal and posterior parietal activation among older adults compared with younger adults during memory tasks (Cabeza et al., 1997; Reuter-Lorenz et al., 2000; Rosen et al., 2002). Prefrontal hyperactivation usually refers to the bilateral activation observed in older adults, whereas younger adults tend to show unilateral activation during working memory and cognitive control tasks (Cabeza et al., 2002; Turner and Spreng, 2015). The greater prefrontal activation seen in older adults is postulated to be compensatory because of the positive association between prefrontal activation and performance on a range of cognitive tasks, including working memory, episodic memory, semantic memory, perception, inhibitory control, and visuospatial processing in older adults (Cabeza, 2002; Reuter-Lorenz et al., 2000; Rypma and D'Esposito, 2000; Turner and Spreng, 2015). Similarly, heightened posterior parietal activation in older adults is associated with improved performance on tasks demanding attention (Huang et al., 2012), planning (Turner and Spreng, 2015), and memory (Sperling et al., 2003; Turner and Spreng, 2012).
A meta-analysis of task-based activation in fMRI studies of older adults showed greater FPN activation in older adults compared with younger adults, and the degree of FPN activation was associated with better memory and executive control task performance (Li et al., 2015). Further supportive evidence from transcranial magnetic stimulation studies showed less detrimental effects of stimulating unilateral prefrontal cortices in older adults than in younger adults, suggesting that additional prefrontal involvement may compensate for unilateral functional deficits in older adults (Manenti et al., 2011; Rossi et al., 2004).
However, some have found that age-related increases in activity in frontal and parietal areas are associated with worse performance on memory tasks (Campbell et al., 2012; de Chastelaine et al., 2011; Drag et al., 2016; Persson et al., 2011; Stevens et al., 2008). Using a longitudinal study design, Nyberg and colleagues (2010) found that between-group analyses suggested age-related increases in task-related frontal recruitment, consistent with most cross-sectional studies, whereas longitudinal within-group analyses revealed task-related decreases in activation with age. Such findings highlight the importance of using more sensitive longitudinal measurements when analyzing the effects of altered activation on cognitive changes over time, although further studies are necessary to replicate these observations.
Thus, while the association between greater frontal/parietal activation and cognition is frequently interpreted as evidence for a compensation mechanism for maintaining or improving cognitive performance during aging, there is evidence against the compensation notion suggesting that reduced brain activation in aging is responsible for cognitive decline.
DMN changes with age
Brain areas that are part of the DMN, such as the medial prefrontal regions, posterior cingulate/precuneus (PCP), lateral parietal, medial temporal, and lateral temporal areas (Ferreira and Busatto, 2013; Greicius et al., 2003; Raichle, 2015), tend to be less active, or become deactivated, during task performance but more active when subjects are at rest. It has been suggested that the degree of task-related DMN deactivation changes with age (Lustig et al., 2003; Raichle et al., 2001). The DMN is thought to be highly susceptible to age-related changes, as it shows reduced task-related deactivation and decreased within-network connectivity with age (Lustig et al., 2003; Raichle et al., 2001). These changes are suggested to occur at an accelerated rate in those with mild cognitive impairment (MCI) and Alzheimer's disease (AD) (see reviews by Grady, 2012; Krajcovicova et al., 2014; Sheline and Raichle, 2013).
During cognitive tasks, older adults display reduced deactivation of the DMN regions compared with younger adults, especially in the PCP (Grady et al., 2006; Lustig et al., 2003; Miller et al., 2008). Grady (2012) suggested that a failure to properly modulate DMN activity may reflect deficient resource allocation with age, especially during challenging tasks. Indeed, reduced deactivation in medial prefrontal and parietal cortices was observed with increasing difficulty in a verbal n-back task in older adults (Prakash et al., 2012). Greater DMN deactivation among older adults has been associated with better performance on memory and executive function tasks (Brown et al., 2019; Miller et al., 2008; Prakash et al., 2012). Using fMRI and diffusion tensor imaging, Brown and colleagues (2019) also found that white matter microstructure degradation in the DMN was associated with more rapid declines in executive function task performance, highlighting the effects of white matter structural changes on functional connectivity and cognition in aging. Together, these findings suggest that the failure of DMN deactivation during task performance, coupled with white matter microstructure decline, may contribute to cognitive decline in older adults.
Older adults also display aberrant DMN connectivity during resting-state scans, particularly showing decreases in within-network DMN functional connectivity (Fig. 1) (Chan et al., 2014; Geerligs et al., 2015; Grady et al., 2016; Zonneveld et al., 2019). Such findings are in general agreement with a meta-analysis showing an inverted U-shape of DMN functional connectivity over the life span, in which within-network connectivity is lowest in childhood, peaks in adulthood, and then falls again in advanced age (Mak et al., 2017). However, a recent longitudinal study from middle age to advanced age suggested a more complex picture by showing that DMN within-network connectivity exhibits an inverted U-shaped relationship with age after controlling for total DMN volume, rising through middle adulthood, peaking in the seventh decade, and then declining steeply in the eighth and ninth decades (Staffaroni et al., 2018). The unexpected increases in DMN connectivity in middle adulthood were speculated to be an outcome of amyloid-induced neuronal excitation (Staffaroni et al., 2018).
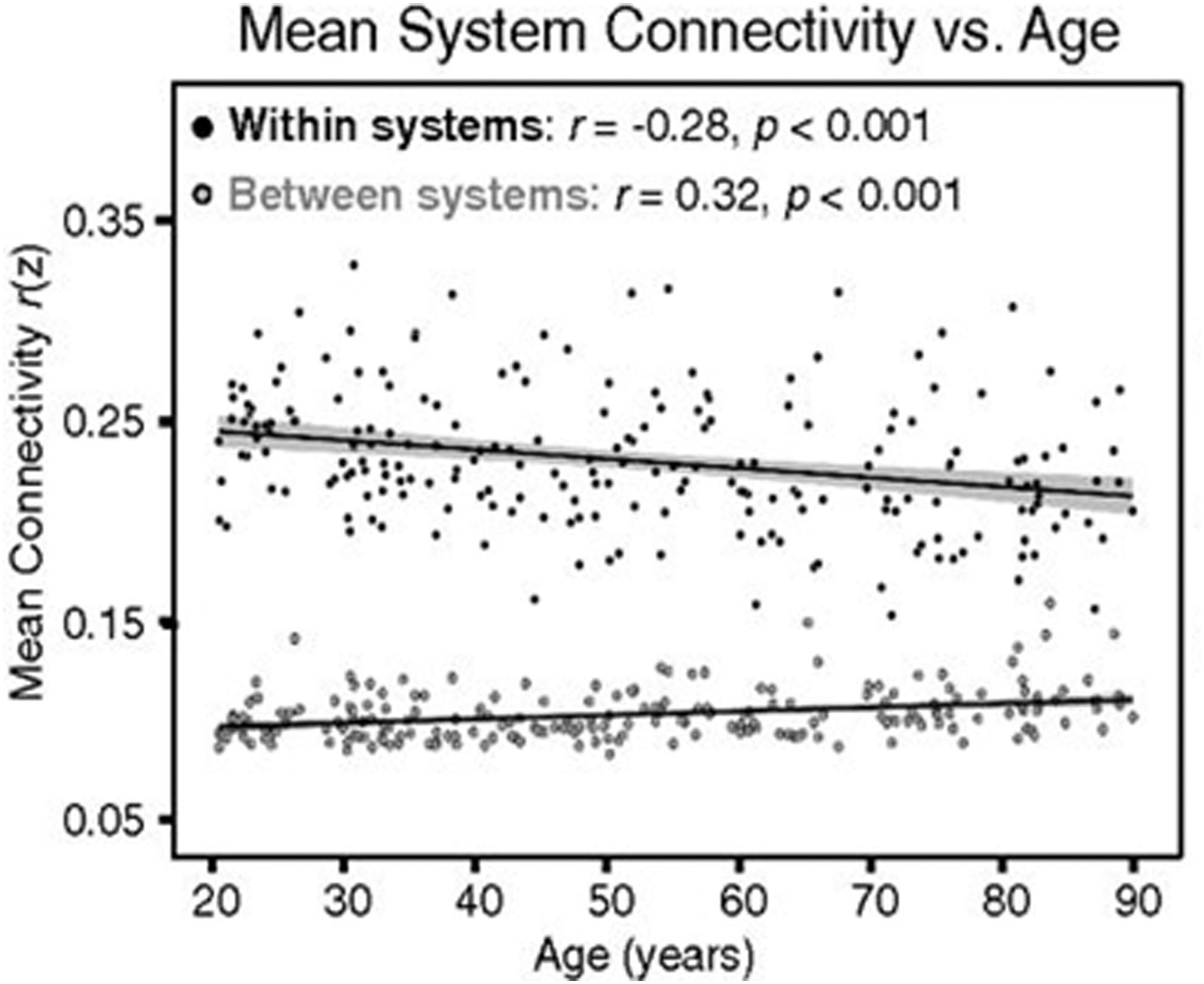
Within- and between-network connectivity changes with age using a graph theory approach. With age, whole-brain within-network connectivity decreases, whereas between-network connectivity increases, although these changes can vary. Increased connectivity between different networks is indicative of decreased network segregation with age (adopted from Chan et al., 2014).
Furthermore, reduced connectivity within the DMN at rest has been associated with decreased episodic memory performance in cross-sectional (Andrews-Hanna et al., 2007; Sala-Llonch et al., 2015) and longitudinal studies (Bernard et al., 2015; Persson et al., 2014; Staffaroni et al., 2018). However, the relationship between DMN connectivity and episodic memory may change over the course of the life span. Fjell and colleagues (2015) observed that across subjects, younger adults with worse memory performance showed greater resting-state DMN within-network connectivity, whereas older adults with better memory performance demonstrated greater DMN connectivity cross-sectionally. A longitudinal study found that within-network DMN connectivity increases with age were positively associated with episodic memory performance during the phase of increasing connectivity in mid-adulthood on an individual level (Staffaroni et al., 2018). Together, these findings suggest that increased DMN connectivity may be detrimental in younger adults but beneficial in some older adults.
Theories from fMRI studies
As described in the last section, while increased brain activity in association with improved or maintained cognitive performance has been observed among older individuals, hyperactivation has also been observed with worse performance on memory tasks among younger individuals (Davis et al., 2008; Fjell et al., 2015; Grady et al., 2005). Below, we briefly describe several proposed models that attempt to explain age-related changes in brain activation in relation to cognition.
The hemispheric asymmetry reduction in older adults model, or HAROLD (Cabeza, 2002), is based on the observation of enhanced frontal activation in older adults involving bilateral prefrontal cortices (Cabeza, 2002; Cabeza et al., 2004), whereas younger adults usually display unilateral frontal activation (Rosen et al., 2002). Evidence supporting the HAROLD model has come primarily from fMRI studies of working memory and memory encoding tasks (Dennis et al., 2007; Piefke et al., 2012) and has been linked to reductions in asymmetric prefrontal connections (Li et al., 2009). The posterior–anterior shift in aging model, or PASA (Davis et al., 2008), explains neuroimaging studies of aging that found increased frontal activation with concomitant reduced activation in posterior visual processing areas (Cabeza et al., 2004; Davis et al., 2008; Grady et al., 1994). Finally, the compensation-related utilization of neural circuits hypothesis model, or CRUNCH (Reuter-Lorenz and Cappell, 2008; Reuter-Lorenz and Lustig, 2005), suggests that age-related activity increases at low task loads and decreases at high task loads. This latter model could explain reports of negative relationships between task performance and frontal overactivation among older adults (Persson et al., 2011; Stevens et al., 2008), both of which may be caused by age-related volumetric decline (Maillet and Rajah, 2013).
However, others challenge these compensation theories based on studies that show increased activity during worse memory task performance (Campbell et al., 2012; Drag et al., 2016; Persson et al., 2011; Stevens et al., 2008). A main alternative notion is the dedifferentiation hypothesis, which suggests that aged brains show less functional specificity during cognitive tasks, displaying more similar patterns of brain activity across multiple tasks compared with younger brains (Goh, 2011; Koen and Rugg, 2019; Park et al., 2004; Rajah and D'Esposito, 2005). Support for this theory has been shown on the cellular level, as nonhuman senescent neurons in visual areas showed decreased sensitivity for specific orientations and directions, and these senescent neurons also showed greater responsiveness to a wider range of orientations and directions compared with younger neurons (Schmolesky et al., 2000).
In a key study evaluating whether prefrontal overactivation truly follows the PASA model, Morcom and Henson (2018) used a model-based multivariate approach in two independent samples to show that additional prefrontal activity did not carry additional information and concluded that prefrontal overactivation is a consequence of dedifferentiation as opposed to compensation. Even across several reviews and meta-analyses, evidence for and against age-related compensatory mechanisms remains unclear, perhaps due to the lack of longitudinal studies and wide heterogeneity across studies in terms of sample sizes and clinical diagnoses, task type and difficulty, and fMRI parameters and analysis pipelines (Grady, 2012; Spreng et al., 2010; Stern et al., 2019).
In sum, questions still remain regarding why only some older adults show prefrontal overactivation and how exactly this overactivation is related to individual differences in task performance. Longitudinal model-based studies are required to better elucidate how increased prefrontal activity relates to cognition with age. In the next section, we will discuss how amyloid plaques and tau tangles can each influence regional activity and functional connectivity, even among those that are cognitively normal (Brier et al., 2014; Hahn et al., 2019; Wisch et al., 2020). As most previous neuroimaging studies of aging do not screen for these pathologies, the mixed findings could originate from inhomogeneous samples, especially because many earlier studies have smaller sample sizes. It is essential to understand the role of these proteins on brain activity and functional connectivity to better understand age-related changes in brain function and cognition during the normal course of aging.
Effects of Pathological Proteins on Functional Connectivity and Cognition
Amyloid-beta plaques and neurofibrillary tau tangles are associated with altered brain function in those with AD (Goedert, 1993; Hardy and Allsop, 1991; Hardy and Higgins, 1992; Selkoe, 1991). Cognitive dysfunction among those with AD has been previous attributed to amyloid (Hanseeuw et al., 2019; Pereira et al., 2019), although declining cognition seems to show a stronger association with tau in recent findings (Hanseeuw et al., 2019; Ossenkoppele et al., 2019). Individuals with AD also show marked declines in functional connectivity, with the DMN being particularly implicated in the current literature, although there are other notable large-scale network alterations (see reviews by Dennis and Thompson, 2014; Hafkemeijer et al., 2012; Krajcovicova et al., 2014; Mevel et al., 2011; Sheline and Raichle, 2013). Less is known about the specific effects of amyloid and tau on functional connectivity and cognition in cognitively normal older adults who harbor these proteins, who may be in the early stages of the AD pathophysiological spectrum. In the rest of this review, we will discuss how amyloid and tau impact functional connectivity and cognitive performance in studies of cognitively normal older adults.
Amyloid-beta
Amyloid accumulation is one of the most notable neuropathologies found in AD and has been thought to be the primary cause of the disease and its symptoms, such as cell loss and dementia, as noted in the amyloid cascade hypothesis (Hardy and Higgins, 1992). Many have attempted to relate amyloid pathology to declining cognitive abilities in those with AD and MCI (see review by Koyama et al., 2012). While cross-sectional results are inconsistent, longitudinal studies and meta-analyses have shown a slight decrease in global cognitive function, particularly in memory domains, among amyloid-positive individuals compared with amyloid-negative individuals (Han et al., 2017; Hedden et al., 2013). Some have been led away from the amyloid cascade hypothesis because the presence of amyloid does not necessitate cognitive dysfunction, as amyloid pathology is found in roughly 30% of cognitively normal older adults (Jansen et al., 2015; Oh et al., 2014), and only about 50% of patients diagnosed with MCI are classified as amyloid-positive (Okello et al., 2009; Wolk et al., 2009). One possible explanation is that the neuropsychological assessments commonly used are not sensitive enough to detect the nuanced amyloid-related cognitive deficits. Another likely explanation is that there may be some other intermediary factor modulating the effect of amyloid on cognition, such as tau (see the next section).
Amyloid deposits in the human brain have been shown to alter neurovascular coupling and neurophysiology (Cohen et al., 2009; Hampel, 2013; Vlassenko et al., 2010; see review by Palop and Mucke, 2010). As such, neuroimaging studies have found amyloid-related changes in regional brain activity and functional connectivity as well as changes in large-scale networks. However, because the effects of amyloid on activity and connectivity in these many networks are not well studied, and because the literature has focused on associations between amyloid and DMN activity/connectivity, we will be primarily discussing amyloid and DMN activity and connectivity. Furthermore, many studies dichotomize amyloid status due to technical limitations and cross-sectional designs, so we present a summary of the literature in the context of differences in brain activity between amyloid-positive and amyloid-negative older adults.
Effect of amyloid on brain activation
Many studies have reported reduced deactivation in DMN regions during memory encoding tasks in amyloid-positive individuals compared with amyloid-negative individuals (Elman et al., 2014; Lim et al., 2014; Mormino et al., 2011; Oh and Jagust, 2013; Sperling et al., 2009). In contrast, amyloid-related hyperactivity has been observed in FPN regions, which was interpreted as a mechanism to overcome the negative effects of amyloid on neuronal function (Oh et al., 2015). However, reduced dorsolateral prefrontal activation during memory encoding with increasing amyloid across a wide age range (30–89 years) has also been observed (Kennedy et al., 2012). These results are not necessarily discordant, based on recent findings of a nonlinear, inverted U-shaped relationship between global amyloid and activation in DMN regions (angular gyri and medial frontal areas) during a spatial judgment task (Foster et al., 2018) and in subcortical regions during an n-back working memory task (Kennedy et al., 2018). Perhaps brain activity increases in those with low amyloid burdens but decreases in those with high amyloid burdens, although this pattern is yet to be demonstrated in task-positive areas (i.e., FPN) and remains to be replicated with longitudinal studies along with concurrent tau measurements.
Effects of amyloid on resting-state functional connectivity
In corroboration with task-based fMRI studies, resting-state fMRI studies have found differences in DMN functional connectivity among AD patients in comparison to healthy older adults (e.g., Greicius et al., 2004; Jones et al., 2011), and this was confirmed by a recent review and meta-analysis (Badhwar et al., 2017). Others further suggested that AD-related amyloid tends to deposit in brain regions that are highly interconnected to other regions, or the so-called cortical hubs, which include areas of the DMN (Buckner et al., 2005; Elman et al., 2016). This pattern of amyloid accumulation has been replicated in other AD cohorts (Buckner et al., 2009; Grothe et al., 2016), as well as in amyloid-positive cognitively normal older adults (Hahn et al., 2019; Mormino et al., 2011; Palmqvist et al., 2017). Furthermore, functional connectivity within the DMN seems to worsen with clinical severity across MCI and AD diagnoses (de Vogelaere et al., 2012; Petrella et al., 2011; Soman et al., 2020; Agosta et al., 2012). Indeed, some evidence suggests a faster aging pattern of connectivity decline in AD patients compared with age-matched controls (Jones et al., 2011).
However, functional connectivity changes among clinically normal older adults are less clear. Although studies investigating the relationship between amyloid and network connectivity have found altered DMN connectivity in those with amyloid burdens compared with those without (Grothe et al., 2016; Hedden et al., 2009; Jones et al., 2016; Mormino et al., 2011), the specific effects of amyloid on functional connectivity in cognitively normal older adults are not all consistent across these studies. Some found that amyloid-positive individuals display decreased within-network DMN functional connectivity (Hedden et al., 2009; Palmqvist et al., 2017), whereas others found the opposite (Hahn et al., 2019; Lim et al., 2014).
Considering Hahn and colleagues (2019) showed evidence that the earliest accumulation of amyloid is associated with increased functional connectivity in cognitively normal adults, it is possible that DMN connectivity changes vary with increasing amyloid levels with age. In addition, studies that examined connectivity between individual DMN nodes in greater detail found both amyloid-related hypo- and hyperconnectivity within the DMN, and they suggested that connectivity among posterior DMN regions decreases and connectivity among anterior DMN regions increases (Elman et al., 2016; Jones et al., 2016; Mormino et al., 2011), consistent with the PASA theory. It is important to note that associations between amyloid burden and aberrant functional connectivity within and across large-scale networks have also been found in the FPN, dorsal attention network, and salience network (Elman et al., 2016).
Taken together, although an overall reduction in within-network connectivity in the DMN among amyloid-positive individuals has been emphasized (Hedden et al., 2009; Palmqvist et al., 2017, and see review by Sheline and Raichle, 2013), changes in DMN connectivity seem to vary by amyloid burden (Foster et al., 2018; Hahn et al., 2019; Kennedy et al., 2018; Mormino et al., 2011) and may not be homogeneous even within the DMN (Elman et al., 2016; Jones et al., 2016; Mormino et al., 2011). However, many reported effects of amyloid on connectivity do not consider tau levels, which have been shown to affect connectivity as well (Schultz et al., 2017; Sepulcre et al., 2017).
Tau
Another major brain pathology found in AD brains is hyperphosphorylated tau, seen as neurofibrillary tangles that primarily aggregate intracellularly, causing axonal dysfunction from within the neuron (de Calignon et al., 2012; Gómez-Isla et al., 1997). Recent findings suggest that, relative to amyloid, tau levels (measured by cerebrospinal fluid [CSF] or positron emission tomography [PET]) are more closely related to cognitive decline (Brier et al., 2016; Johnson et al., 2016; Pontecorvo et al., 2017), brain volume loss (LaPoint et al., 2017; Maass et al., 2018), and hypometabolism (Adams et al., 2019; Hanseeuw et al., 2017). With the recent emergence of the PET tracer, flortaucipir (18F-AV1451) (Chien et al., 2013), which provides an in vivo visualization of tau distribution in the human brain, the role of tau on brain function has been quickly gaining attention. Although the negative effects of tau deposition on brain structure and cognition are becoming clearer (see reviews by Hanseeuw et al., 2019; Schmand et al., 2010), the associations between tau and alterations in functional connectivity are understudied.
Tau localization and spread within neural circuits
Braak and Braak (1991) first categorized the staging of AD pathology by analyzing the spatial patterns of amyloid and tau deposition in postmortem tissue. Unlike amyloid, which accumulates diffusely throughout the cortex, tau spreads through the brain in relatively distinct stages, beginning in the entorhinal cortex and other MTL structures, then spreading to other limbic and temporal areas, and eventually reaching nearly all areas of the cortex (Braak and Braak, 1991; Schöll et al., 2016). This Braak staging of tau pathology has been supported by recent tau-PET studies of older adults and AD patients (Schöll et al., 2016; Schwarz et al., 2016). A recent study further demonstrated that tau pathology first appears specifically in the transentorhinal and anterolateral entorhinal cortex, and less so in the posteromedial entorhinal cortex (Adams et al., 2019).
It has recently been shown that hyperphosphorylated tau oligomers can spread trans-synaptically via a prion-like mechanism (Kaufman et al., 2016; Mudher et al., 2017), propagating through interconnected brain areas in an activity-dependent manner (Pooler et al., 2013; Wu et al., 2016). Similarly, activity-dependent tau spread has been inferred from PET-fMRI studies (Cho et al., 2016; Franzmeier et al., 2019; Hansson et al., 2017; Hoenig et al., 2018; Sintini et al., 2020). One popular hypothesis explaining the spread of tau in the brain is the network degeneration hypothesis, which suggests that pathological proteins spread through functional networks, based on the finding of high levels of tau deposition in a group of regions that are highly interconnected with one another, such as the transentorhinal cortex and the hippocampus (Franzmeier et al., 2019, 2020; Kfoury et al., 2012). A recent PET-fMRI study further showed that tau levels are better explained by connectivity between nodes rather than merely proximity to areas of high tau deposits and that baseline connectivity and tau-PET measures can predict future tau-PET accumulation in amyloid-positive individuals (Franzmeier et al., 2020). These studies together provide convincing evidence supporting the trans-neural spread of hyperphosphorylated tau through functional networks.
Functional connectivity changes with tau
Because tau-PET tracers have not been available for long, there have been few functional neuroimaging studies examining the relationship between tau spread and functional connectivity changes. In some initial studies, several groups have examined the spatial pattern of tau deposition in relation to the functional networks identified by fMRI-based resting-state functional connectivity analysis. While they consistently demonstrated that the patterns of tau pathology in AD patients overlap with known functional networks, such as the DMN and FPN (e.g., Franzmeier et al., 2019; Hoenig et al., 2018), it is unclear whether or not tau accumulation differentially affects the DMN or FPN in AD (Hansson et al., 2017).
Several recent studies have investigated the potential impact of tau pathology on functional network connectivity. The first study to do so, to the best of our knowledge, was by Malpas and colleagues (2016), who showed that increased CSF tau is associated with reduced functional connectivity involving the right anterior entorhinal cortex in patients with AD-related dementia. Since then, several more tau-PET studies suggested detrimental effects of tau on brain connectivity in older adults and in AD patients; increasing tau levels have been associated with weakened connectivity within multiple networks (Cope et al., 2018; Wisch et al., 2020) and between highly interconnected hubs (Cope et al., 2018), as well as with reduced global and local network efficiency (Sintini et al., 2020). A PET-fMRI study of cognitively normal older adults, however, found that increased posterior DMN connectivity to brain areas outside the DMN was positively associated with global levels of amyloid and local levels of tau (Quevenco et al., 2020). Since the sample was composed of individuals with low amyloid levels, it is possible that the posterior DMN may be showing amyloid-related hyperactivity at this stage. In sum, these findings suggest that tau pathology may spread through intact functional networks in early stages, then, with increasing tau load, connectivity in these functional networks may falter.
A caveat of the previously discussed studies is that nearly all subjects who show substantial tau deposition also harbor an amyloid burden, so it is difficult to disentangle the effects of amyloid versus tau on functional connectivity. A tau-PET fMRI study in a cohort with substantial tau pathology, but not amyloid, such as in those with primary-age-related tauopathy (PART) (Crary et al., 2014), could better elucidate the specific relationship between tau and functional connectivity. Recent studies show that PART individuals typically display cognitive decline and MTL atrophy at a greater rate than age-matched controls but slower than AD patients (Bell et al., 2019; Besser et al., 2019; Josephs et al., 2017; Quintas-Neves et al., 2019). PART has typically been studied in postmortem brains, so little is known about the functional connectivity profiles of PART brains in vivo. In clinical and preclinical AD, however, amyloid and tau are present in the brain and may have variable and combined effects on functional connectivity, as discussed below.
Amyloid–tau interaction hypothesis
Since neither amyloid nor tau alone can account for the various observations of neurodegeneration patterns and cognitive decline (e.g., Schöll et al., 2016), it has been suggested that amyloid and tau may interact and produce compounded effects of cell toxicity and symptoms of dementia (Ittner and Götz, 2011). Because amyloid and tau accumulate in an activity-dependent manner (Hahn et al., 2019; Pooler et al., 2013; Wu et al., 2016), amyloid-induced hyperactivity (in early stages) may be responsible for more rapid tau spread and further amyloid accumulation across the brain (Busche et al., 2008, 2012), and high amyloid levels may contribute to even greater tau accumulation (Tosun et al., 2017). Consequently, having high levels of both amyloid and tau may lead to rapid cognitive decline, neurodegeneration, and altered functional connectivity, as we will explore next.
Interactive effects on cognition, metabolism, and atrophy
Studies analyzing the impacts of both amyloid and tau on cognition have shown that, compared with amyloid, tau is more closely linked to cognitive impairments, including global cognition, episodic memory, delayed recall, and executive function (Aschenbrenner et al., 2018; Hanseeuw et al., 2019; Pereira et al., 2019). Recent longitudinal studies, however, have suggested that older individuals with high levels of both amyloid and tau tend to show more rapid cognitive decline than those who harbor only predominantly amyloid or tau (Betthauser et al., 2020; Sperling et al., 2019).
Recent findings further suggest that multiple pathologies can produce compounding effects on the brain and worsen cognitive outcomes. It has been demonstrated that amyloid-positive individuals show an inverse relationship between MTL tau and hippocampal volume, but hippocampal volume was not related to amyloid levels alone (Wang et al., 2016). Importantly, older adults who displayed substantial amyloid, tau, and hippocampal atrophy showed the most rapid cognitive decline compared with those without all three pathologies (Aschenbrenner et al., 2018). Areas with high levels of colocalized amyloid and tau deposition have also been related to reduced regional glucose metabolism (Adams et al., 2019; Bischof et al., 2016), and this effect is commonly reported in the posterior cingulate cortex (Hanseeuw et al., 2017; Pascoal et al., 2017). These findings suggest that hippocampal atrophy, hypometabolism, and cognitive decline are more closely related to tau in the presence of amyloid and that their co-occurrence results in more rapid decline than either protein acting alone (Betthauser et al., 2020; Sperling et al., 2019, see review by Busche and Hyman, 2020). Such relationships likely change over time (Jagust, 2018), requiring further investigation using longitudinal approaches.
Interactive effects on functional connectivity
While there have been few studies measuring the relationship between tau on functional connectivity (see the section “Functional connectivity changes with tau”), even fewer have examined the effects of both amyloid and tau on functional connectivity. In a recent study, Quevenco and colleagues (2020) found that in cognitively unimpaired older adults, local amyloid and tau levels were related to greater functional coupling between a PCP seed, a central component of the DMN, and various cortical regions. Since their analysis focused on the PCP, it is not clear whether these are unique effects on DMN connectivity or if other networks are similarly affected.
Two studies examined the relationship between amyloid, tau, and connectivity in greater detail. First, Sepulcre and colleagues (2017) found that the majority of cognitively normal elderly individuals showed a negative association between tau and whole-brain connectivity but a positive association between amyloid and whole-brain connectivity. More specifically, hyperconnectivity was more commonly found in regions with high amyloid and low tau, whereas hypoconnectivity was more commonly found in regions with low amyloid and high tau levels (Sepulcre et al., 2017). Intriguingly, high levels of both amyloid and tau were found in the so-called linkage areas, where hyper- and hypoconnectivity converge, likely accounting for the observation of both forms of connectivity changes in the same brains (Sepulcre et al., 2017). Many of these linkage areas overlap with DMN areas, which might explain the heterogeneous literature regarding altered DMN connectivity in previous studies of older adults.
Around the same time, Schultz and colleagues (2017) found an inverted U-shaped relationship between within-DMN functional connectivity and an interaction term reflecting a combination of global amyloid and local tau levels, such that DMN connectivity rises and falls as the interaction term (global amyloid by local tau) increases (Fig. 2). This is consistent with Quevenco and colleagues' (2020) finding of increased pathology-associated posterior DMN connectivity in individuals with low amyloid levels, who may be showing connectivity patterns similar to the initial rising phase of the inverted U function.
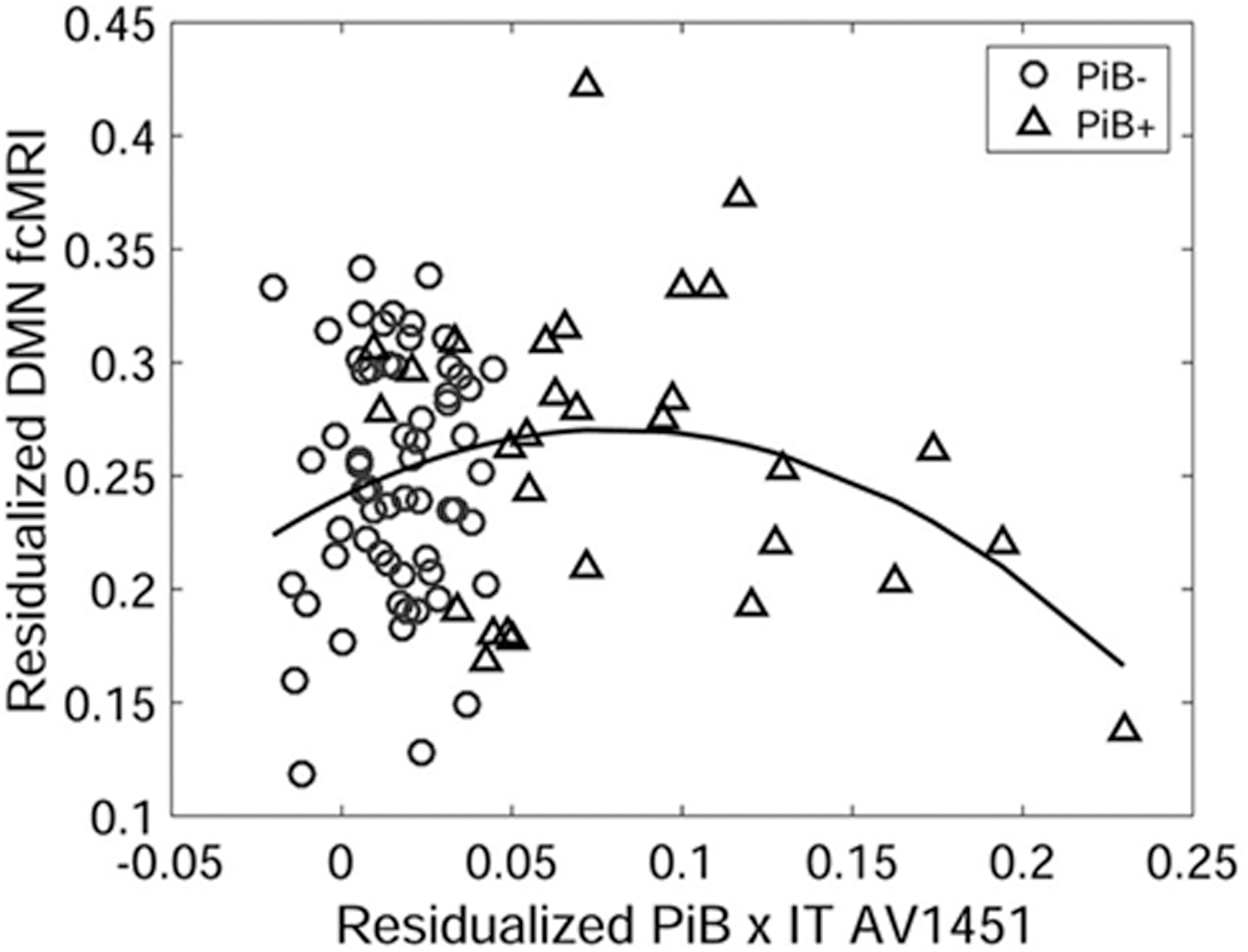
A study by Schultz and colleagues (2017) illustrated DMN connectivity as a function of the interaction of global amyloid and inferotemporal tau. Within-network DMN connectivity appears to increase and decrease with rising inferotemporal tau levels among amyloid-positive individuals (adopted from Schultz et al., 2017). DMN, default mode network.
In sum, the few studies of amyloid and tau interactions, together with other tau-related studies, suggest that tau has detrimental effects on functional connectivity at both the modular network and large-scale network level and may depend on the concurrent level of amyloid. While amyloid and tau are associated with hyper- and hypoconnectivity, respectively, they are ultimately involved in decreasing neural efficiency, and their interactivity may result in stronger effects of tau on brain functional deficits.
A model of functional connectivity changes related to amyloid and tau accumulation
Here, we propose a model highlighting the seemingly opposing effects of amyloid and tau on functional connectivity found in some previous studies and how they relate to hyperactivity and then hypoactivity with increasing amyloid and tau accumulation (Fig. 3). This model builds upon one proposed by Jagust and Mormino (2011), which shows hyper- and hypoactivity with age as a result of amyloid burden. A key ingredient missing from this previous model is tau because the tau-PET tracer was not available at that time. Although the shape of their amyloid-induced hyperactivity and then hypoactivity curve is similar to what we propose, more recent findings suggest that the cause of the hypoactivity is more likely related to tau accumulation and/or an interaction between amyloid and tau (see the section “Amyloid–tau interaction hypothesis”). However, there is also evidence that high levels of amyloid may result in reduced activation (Foster et al., 2018; Kennedy et al., 2012), which may be an effect of advancing age and tau buildup, although this cannot be ruled out without further testing. In our model, functional connectivity alterations as a function of age in presumably normal aging (simplified as decreased within-network and increased between-network connectivity) are shown as a baseline, assuming little-to-no amyloid or tau.
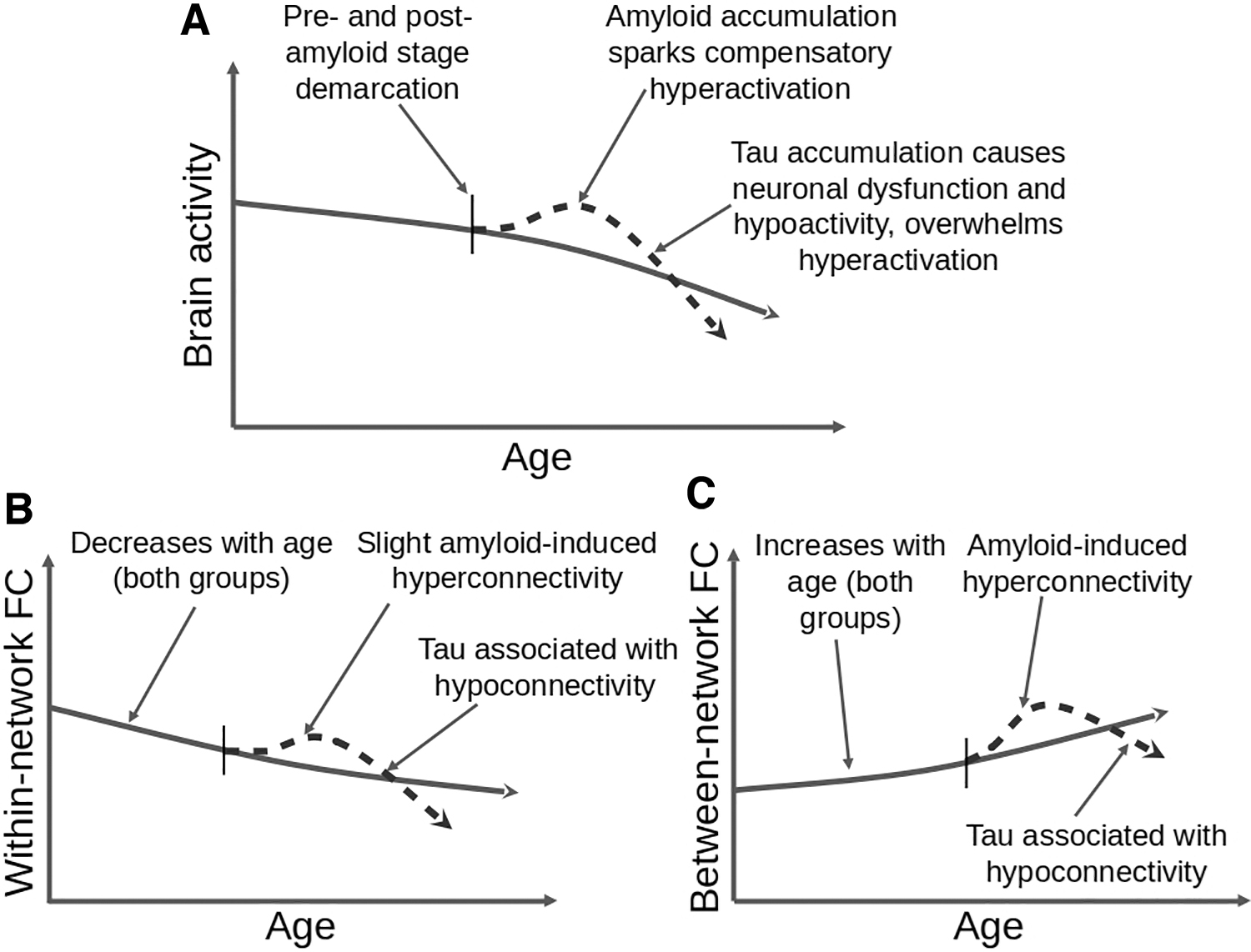
Proposed model showing the effects of amyloid and tau on fMRI-based brain activity and connectivity based on Jagust and Mormino (2011). Panel
It is important to note that the onset and timing of amyloid and tau accumulation are highly variable across individuals. Because PET studies mostly dichotomize amyloid positivity, little is known about the dose-dependent effects of amyloid in amyloid-positive individuals. Also, those with greater cognitive abilities at baseline may be more resilient to the accumulation of these pathological proteins (Snowdon et al., 1996). Based on this evidence, Jagust and Mormino (2011) suggested that the timing and onset of amyloid-related connectivity changes vary with cognitive ability. Clearly, further longitudinal testing and multimodal designs are required to determine to what extent baseline cognitive ability is associated with the onset of subsequent amyloid- and tau-related functional changes.
Conclusions
Older brains show changes in brain chemistry, vasculature, and neuronal morphology which can each influence brain function. From these age-related anatomical and physiological changes can arise cognitive changes and deficits, which may be mediated by altered regional activity and network connectivity. Overall, within-network connectivity tends to decrease, and between-network connectivity tends to increase in aging, although recent findings suggest more likely that there is an inverse U-shape trajectory of within-network connectivity (e.g., DMN) across the life span. Connectivity in the DMN and FPN has been shown to be affected by both amyloid and tau in various ways, even among cognitively unimpaired older adults, which may explain earlier inconsistent reports of both increased and decreased functional network connectivity with age. Although the spatial pattern of tau spread overlaps with brain networks showing hypoconnectivity, such as the DMN and FPN, it is still unclear how tau independently affects functional connectivity and should be studied directly in those who display primary age-related tauopathy. Phases of hyper- and hypoconnectivity seen in clinically normal older adults may be caused by amyloid-induced hyperactivation, followed by tau-induced neural dysfunction and hypoconnectivity, although these ideas require experimental confirmation.
There are several limitations we have recognized while discussing the neuroimaging studies of aging. We have not written an exhaustive review on some of the broader topics included here, so we have suggested other pertinent reviews. Regarding the compensation/dedifferentiation debate, methodological differences across studies, such as population demographics, task type and parameters, fMRI parameters, and fMRI analysis pipelines (seed-based vs. whole-brain connectivity), combined with an imprecise definition of compensation, may all have led to a heterogeneous literature that requires further replication. Despite this, a recent meta-analysis indicated that, although different studies may report different brain regions relating to AD or other conditions, each reported region can be linked to the same functional network (Darby et al., 2019); therefore, future functional imaging studies should take a more network-based approach when studying neurodegenerative diseases.
From a clinical standpoint, concerns arise regarding labeling populations as typical/healthy agers versus those with preclinical AD, as all older adults develop some degree of amyloid accumulation, and some even develop a tau burden while remaining cognitively normal. This review primarily summarizes studies concerning the effects of AD-related protein pathology in the brains of cognitively normal older adults but does not focus on AD itself. Other age-related factors besides amyloid and tau accumulation may cause functional connectivity changes, such as neuroinflammation and oxidative stress, neuronal atrophy, and neurovascular changes (Berlett and Stadtman, 1997; Fabiani et al., 2014; Raz and Rodrigue, 2006).
Finally, the present literature would benefit from large, multimodal, and longitudinal studies (see Brown et al., 2019; Jacobs et al., 2018; Zonneveld et al., 2019), which could determine how amyloid–tau-related connectivity alterations progress with age over a longer time span. Future studies should also pay attention to the various imaging and analysis parameters and consider subnetwork properties for better reproducibility (Gorgolewski and Poldrack, 2016). Elucidating the effects of pathological proteins on cognition and functional connectivity is paramount to understand both healthy and pathological aging.
Footnotes
Authors' Contributions
R.M.W. wrote the initial article, and H.-C.L. provided guidance and significant edits.
Acknowledgment
We acknowledge the work of Michelle Wong, who helped to gather and organize articles. We would also like to thank Dr. Hwamee Oh for initial discussions at the early stages of manuscript preparation.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by Stony Brook Research Foundation.