
Abstract
Background:
Tractography uses diffusion magnetic resonance imaging to noninvasively infer the macroscopic pathways of white matter fibers and it is the only available technique to probe in vivo the structural connectivity of the brain. However, despite this unique and compelling ability and its wide range of possible neurological applications, tractography is still limited, lacks anatomical precision, and suffers from a serious sensitivity/specificity trade-off. For this reason, in the past few years, tractography postprocessing techniques have emerged and proved effective for improving the quality of the reconstructions. Among them, the Convex Optimization Modeling for Microstructure Informed Tractography formulation allows incorporating the anatomical prior that fibers are naturally organized in fascicles, and has obtained exceptional results in increasing the accuracy of the estimated tractograms.
Methods:
We propose an extension to this idea and introduce a multilevel grouping of the streamlines to capture the white matter arrangement in fascicles and subfascicles. We tested our proposed formulation in synthetic and in vivo data.
Results:
Our experiments show that using multiple levels allows considering information about the white matter organization more adequately and helps to improve further the accuracy of the resulting tractograms.
Conclusion:
This new formulation represents a further important step toward a more accurate structural connectivity estimation.
Impact statement
Tractography is an invaluable tool for studying noninvasively the neuronal architecture of the brain, but recent studies have shown that the presence of a large number of false positives can significantly bias any connectivity analysis. Recently, a filtering technique called Convex Optimization Modeling for Microstructure Informed Tractography (COMMIT)-2 has proven particularly effective in dramatically reducing their incidence by considering the prior knowledge that white matter fibers are organized in fascicles. In this work, we propose an extension to this method, which allows us to increase further the anatomical accuracy of tractography reconstructions. Our new formulation represents an additional step forward toward a more veridical characterization of brain connectivity.
Introduction
Diffusion-
Despite these unique opportunities, several studies have recently exposed the poor anatomical accuracy of tractography, showing, in particular, that the estimated tractograms are not truly quantitative and present problems such as partial, duplicate, and false-positive streamlines, that is, pathways that are recovered but are not anatomically valid (Drakesmith et al., 2015; Girard et al., 2020; Maier-Hein et al., 2017; Rheault et al., 2020b; Schilling et al., 2019; Thomas et al., 2014; Zalesky et al., 2016). In particular, Zalesky et al. (2016) investigated the topological properties of the networks estimated with tractography and concluded that specificity (true-negative rate) is more important than sensitivity (true-positive rate) for the study of brain connectivity. Maier-Hein et al. (2017) conducted a large comparison study and showed that false-positive streamlines are a major problem that affects all tractography algorithms. Therefore, it is clear that the presence of false positives in the estimated tractograms represents a crucial limitation of tractography that can heavily bias any analysis based on this technique.
A number of postprocessing solutions have been recently proposed to tackle this problem and improve the quality of the tractograms. This class of methods is called Microstructure Informed Tractography (Daducci et al., 2016) and their rationale is that tractography needs more information to overcome the problem of anatomical inaccuracy. In general, these approaches combine the input tractogram with signal forward models and evaluate the contribution of each streamline to the acquired data, filtering out the most implausible or those that are not supported by the data. The Spherical-deconvolution Informed Filtering of Tractograms (SIFT; Smith et al., 2013) considers the integral of the fiber orientation distribution to be proportional to the volume of the white matter tissue inside the voxel and uses this value to optimize the streamline density and to remove streamlines that do not fit the data. SIFT2 (Smith et al., 2015) does not remove streamlines, but it rather attempts to determine the cross-sectional area of each streamline and uses this information as a multiplier in the streamline connectivity quantification problem. The Linear Fascicle Evaluation (Pestilli et al., 2014) and the Convex Optimization Modeling for Microstructure Informed Tractography (Convex Optimization Modeling for Microstructure Informed Tractography [COMMIT]; Daducci et al., 2013, 2015) use similar approaches, as both estimate the contributions of the streamlines from the full DW-MR signal by solving a linear system. The filtering with all these methods showed promising results for improving the quality of the tractograms (Daducci et al., 2013, 2015; Pestilli et al., 2014; Smith et al., 2013, 2015), but none of them proved effective in reducing the false positives (Schiavi et al., 2020).
To tackle this limitation, COMMIT was recently extended to take into account the fundamental knowledge that fibers in the brain are naturally organized in fascicles. In fact, developmental neurobiology studies have demonstrated that “axons, as they grow, remain neighbors with axons that come from neighboring neurons in the presynaptic structure, and therefore arrive at the postsynaptic structure already topographically arranged” (Udin and Fawcett, 1988). The novel formulation, called COMMIT2 (Schiavi et al., 2020), proved effective in increasing the specificity of the tractograms without decreasing their sensitivity. However, if both true- and false-positive streamlines are reconstructed between two cortical regions, as illustrated in Figure 1 (top), then COMMIT2 would promote or penalize them as a whole and would not be able to disentangle them.

Possible improvement over the COMMIT2 formulation. Top: This simplistic example shows some representative streamlines between two cortical regions obtained with a generic tractography algorithm. This bundle consists of three TP sub-bundles (blue, red, and green), but there are also some streamlines that seem to describe implausible trajectories (purple), which are probably FPs. However, using the parcellation-based approach introduced in COMMIT2, all these streamlines are grouped together as one unique bundle and considered a whole, and so, they cannot be differentiated in true and false trajectories. Bottom: Example of an actual bundle of streamlines from in vivo data as reconstructed in the input tractogram and after filtering with COMMIT2, using either large or small groups, and with COMMIT2tree using both large and small groups in a hierarchical structure. This bundle consists of plausible streamlines that directly connect the brainstem to the postcentral gyrus (cyan arc), as well as of implausible streamlines that incorrectly pass through the corpus callosum (orange arrow). COMMIT, Convex Optimization Modeling for Microstructure Informed Tractography; FP, false positive; TP, true positive. Color images are available online.
In this work, we propose a strategy to further improve the accuracy of the tractograms by extending the assumption made in COMMIT2 and considering that fibers in the brain are anatomically organized in fascicles and subfascicles, for example, the pyramidal tract is formed by the corticospinal tract and the corticobulbar tract (Chenot et al., 2019; Nieuwenhuys et al., 2008). This biological organization of the fibers naturally induces a hierarchy in the reconstructed streamlines, which in turn could be exploited to provide more information to the filtering procedure. To test the effectiveness of this additional anatomical prior, we created a nested arrangement of the streamlines into bundles and subbundles by refining the parcellation-based approach of COMMIT2 using clustering techniques. The proposed method was tested both on synthetic and in vivo data.
Materials and Methods
Microstructure informed tractography
Given a tractogram ℐ and the corresponding DW-MRI  , where
, where 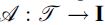 is an operator describing the signal contribution of each streamline to
is an operator describing the signal contribution of each streamline to 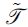 that best describes the acquired image
that best describes the acquired image
where the vector 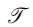 and possibly other tissues inside each voxel according to a given multicompartment model, and
and possibly other tissues inside each voxel according to a given multicompartment model, and

To address the problem of false positives, COMMIT was recently extended by Schiavi et al. (2020), who added a regularization term to take into account that fibers in the brain are naturally organized in fascicles and the streamlines can be grouped in bundles:
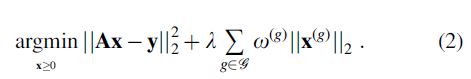
In this formulation, called COMMIT2, 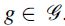 . This additional penalty term promotes solutions that explain the measured DW-MR signal using a minimal number of groups and the parameter
. This additional penalty term promotes solutions that explain the measured DW-MR signal using a minimal number of groups and the parameter
where
Adding priors on the hierarchical organization of the fibers
In the previous formulation, the partition
To take into account this organization of the streamlines, we implemented the hierarchical sparse encoding approach proposed by Jenatton et al. (2011), which uses sparsity-inducing norms as regularization terms to promote tree-structured sparse solutions. The set of groups  are created by splitting each group
are created by splitting each group  , creating a nested structure. The reconstruction problem can be written as follows:
, creating a nested structure. The reconstruction problem can be written as follows:
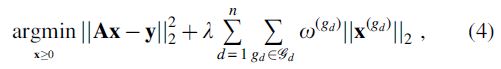
where 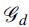 represents a partition into groups of level d,
represents a partition into groups of level d, 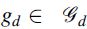 , and
, and
where
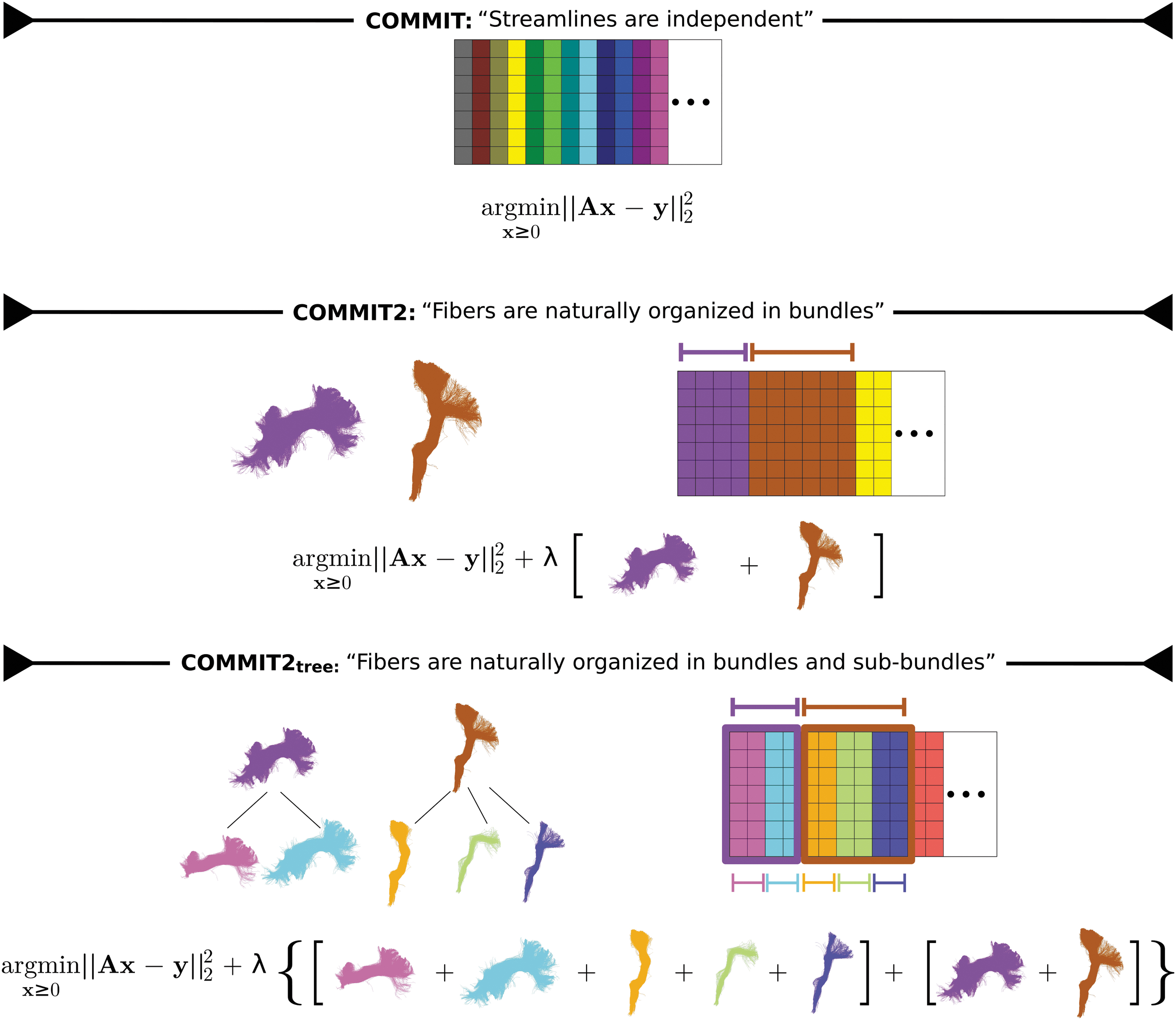
Visual comparison between the COMMIT, COMMIT2, and COMMIT2tree formulations. COMMIT considered the streamlines as independent entities, and COMMIT2 added a regularization term to take into account that streamlines are instead naturally organized in bundles. With COMMIT2tree we propose to improve this regularization by allowing a multilevel hierarchical organization of the streamlines, where every bundle may be composed of several sub-bundles and each can be selectively promoted or penalized. Color images are available online.
Illustrative toy example
Figure 3 shows a simple synthetic example to illustrate how the introduction of the hierarchical regularization can improve the accuracy of the tractograms. The ground truth consists of four ROIs (A, B, C, and D) connected by three streamlines (S1, S2, and S3). From this data set, a generic tractography algorithm (top row, center) reconstructed the three true-positive streamlines as well as two false positives (S4 and S5). This tractogram was then filtered with COMMIT, COMMIT2, and COMMIT2tree. Without any prior information, COMMIT keeps all five streamlines (top row, right), as they are all compatible with the underlying data. COMMIT2 groups streamlines based on a partition with a single level and attempts to find solutions that explain the data with the minimum number of groups. If large groups are used (bottom row, left), for example, streamlines that connect the same pair of ROIs, the group formed by S3 and S4 is necessary to explain the data in the top-right voxel, which is covered only by S3. Therefore, both S3 and S4 are kept even though S4 is a false positive. This choice, in turn, requires to keep also S5 to explain the data in the remaining voxels using the minimum number of groups. This causes the removal of the true positives S1 and S2 because they are no longer necessary to fit the data. In general, if small groups are used (bottom row, center), for example, dividing B into two subregions B1 and B2, and C into C1 and C2, we obtain more groups consisting of fewer streamlines each; in this example every group has a single streamline. This situation contains many ambiguous configurations as seen before for COMMIT (top row, right) and, as groups are treated as independent entities, these ambiguities cannot be solved. Please note that this situation was already analyzed in Schiavi et al. (2020; Fig. 3). On the contrary, the grouping implemented in COMMIT2tree is more flexible and helps to disentangle ambiguous configurations such as the one presented here. In fact, according to the hierarchical structure, the groups S3–S4 can be further subdivided into two groups (bottom row, right), that is, one containing the true positive S3 and the other one the false positive S4. This allows COMMIT2tree to remove all the false positives and keep the true positives.

Illustrative toy example. First row: ground-truth streamline configuration as well as representative streamlines reconstructed with a generic tractography algorithm, that is, raw tractogram, and after filtering with COMMIT. Second row: results after filtering with COMMIT2 (single level, once using large groups and another with small groups) and COMMIT2tree (multiple levels, combining parcellation and clustering). S1, S2, and S3, in green shades, represent TP streamlines, while S4 and S5, in red shades, are FPs.
Data and experiments
For the scope of this work and to keep the presentation simple, we demonstrate the improvements of COMMIT2tree over the previous formulation using a hierarchical structure with only two levels; however, our proposed formulation can easily accommodate any number of levels. In the first level, we grouped the streamlines connecting the same pair of cortical regions as in COMMIT2. Then, to capture the presence of possible sub-bundles, we constructed a second level where we subdivided these parcellation-based groups by clustering the streamlines in each with QuickBundles (Garyfallidis et al., 2012). In brief, QuickBundles is an unsupervised clustering algorithm for tractography data sets, which does not require knowing the number of clusters in advance, but it rather uses a threshold to group streamlines based on distance metrics. In this work, we used the Average of Pointwise Euclidean Metric. The algorithm is very fast and well suited for large tractograms, as its complexity is O(kN), where k is the number of clusters and N the number of streamlines [for technical details see Garyfallidis et al. (2012)]. For a more direct comparison with COMMIT2, we adopted the same forward model as in Schiavi et al. (2020), which associates a contribution to each streamline proportional to its cross-sectional area using a map of the intra-axonal signal fraction (IASF).
Quantitative evaluation on synthetic data
To quantitatively evaluate the performance of our proposed formulation with respect to COMMIT2, we adopted the same synthetic phantom used in Schiavi et al. (2020). We compared the sensitivity and specificity of the tractograms after filtering with both techniques by assessing the number of valid bundles (VB), that is, true-positive connections in the corresponding connectomes, and invalid bundles (IB), that is, false-positive ones. The phantom is shown in Figure 4A and consists of 27 ground-truth bundles between 53 ROIs that mimic challenging configurations such as branching, kissing, and crossing with different curvatures and sizes (Caruyer et al., 2014). The IASF map of this phantom was computed from the ground-truth geometry. The corresponding DW-MR signal was generated using the Composite Hindered And Restricted Model of Diffusion (Assaf and Basser, 2005) along 64 directions with b = 3000 s/mm2 and Rician noise was added with a signal-to-noise ratio of 30. All tractograms and the corresponding connectomes were estimated using the MRtrix3 software (Tournier et al., 2019). First, we computed the fiber orientation distributions in each voxel using Constrained Spherical Deconvolution (Tournier et al., 2007). Then, we reconstructed 1 million streamlines with both deterministic (Tournier et al., 2012) and probabilistic (Tournier et al., 2010) algorithms, using a white matter mask as seeding region and default parameters. Finally, we assigned each endpoint of a streamline to a node if that point fell within 2 mm from one of the 53 gray matter ROIs. A streamline was considered connecting two nodes if both endpoints were assigned, otherwise it was discarded from the analysis.
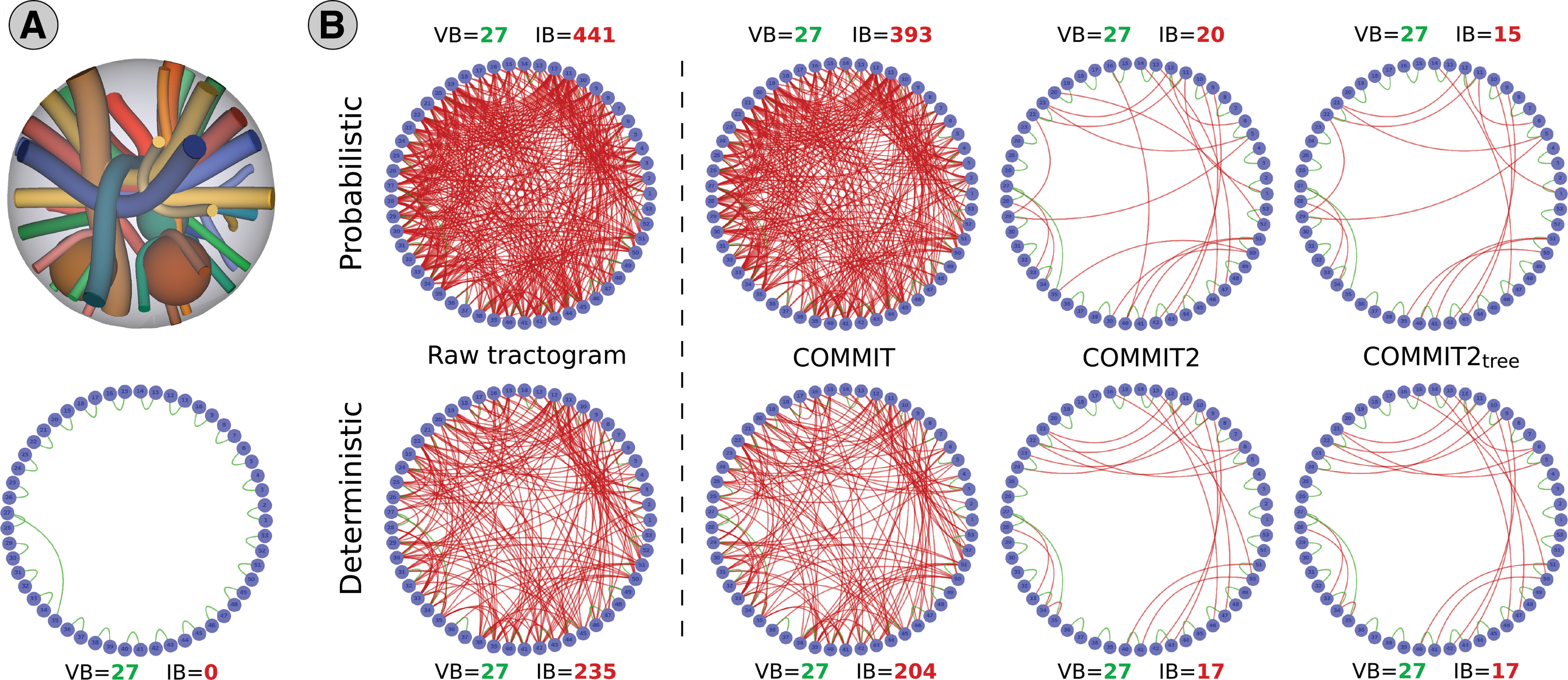
Quantitative comparison on synthetic data between COMMIT2 and COMMIT2tree. We compared the VB (reported in green) and IB (reported in red) in the connectomes generated on a synthetic phantom
Qualitative evaluation on in vivo data
To appreciate the improved quality of the tractograms, we visually inspected known true- and false-positive bundles in 10 subjects (5 males and 5 females of ages 26–30 years) of the open access data of the Human Connectome Project (HCP; Van Essen et al., 2013). These bundles were extracted with RecobundlesX (Garyfallidis et al., 2018; Rheault, 2020), a multiatlas, multiparameter segmentation tool that uses bundles manually segmented by an expert neuroanatomist as reference (description in Appendix A1). We performed RecobundlesX using nine different parameter configurations and a threshold for the voting system of 75%. If a streamline had been selected as true positive, it was not taken into account in the segmentation of the false-positive bundles. The intersections of streamlines in different bundles were eliminated, that is, each streamline could only belong to one bundle. We also reported the voxel coverage of each of them, that is, the number of voxels traversed by the streamlines associated with each bundle. We segmented the T1-weighted image using the FMRIB's Automated Segmentation Tool (Zhang et al., 2001) to derive the multitissue image and perform the multishell, multitissue constrained spherical deconvolution (Jeurissen et al., 2014) on the preprocessed DW-MRI data (Glasser et al., 2013). We generated 5 million streamlines of length between 20 and 200 mm with the probabilistic algorithm (Tournier et al., 2010), using Anatomically Constrained Tractography (Smith et al., 2012) and default parameters. As in the synthetic experiments, the white matter mask was used as seeding region. The IASF map needed by the forward model used in COMMIT2 and COMMIT2tree was computed using the Spherical Mean Technique (Kaden et al., 2016). Besides inspecting individual bundles, we also compared the resulting connectomes, which were created using the 85 gray matter ROIs defined in the Desikan–Killiany atlas (Desikan et al., 2006) derived from the T1-weighted images using FreeSurfer, and replacing the brainstem with its last part, that is, medulla oblongata (Iglesias et al., 2015). Finally, as edge weights we used the streamline count for the raw tractograms and the sum of the estimated streamline contributions for COMMIT2 and COMMIT2tree. For each connectome, we report its density, that is, the ratio between the actual and the possible connections.
Results
Figure 4 compares the sensitivity and specificity of the tractograms estimated from the synthetic data with both deterministic and probabilistic tracking, before and after filtering with COMMIT2 and COMMIT2tree. As a reference, results with COMMIT are also reported. All reconstructions included the 27 true-positive bundles and, as expected, the raw tractograms were contaminated by a large number of false-positive bundles, respectively, IB = 441 (probabilistic) and IB = 235 (deterministic). Only few IB were removed with COMMIT, whereas COMMIT2 was able to drastically reduce them from 441 to 20 (probabilistic) and from 235 to 17 (deterministic). Using the proposed multilevel regularization, COMMIT2tree was able to further improve these results, although not substantially. Using different distance metrics in the clustering did not produce particular effects on the results (Supplementary Fig. S1), probably due to the simple geometry of this phantom.
Figure 5 evaluates the effectiveness of the multilevel formulation in real brain data, where the fiber configurations are expected to be more complex. We qualitatively compared known true- and false-positive bundles obtained in one HCP subject after filtering the tractograms with COMMIT2 and COMMIT2tree. The volume coverage of the bundles is reported in terms of number of voxels and, in the last two cases, we also report the estimated contributions (weight). The filtering with COMMIT2 does not negatively affect the true-positive bundles, that is, their volume is comparable with the raw tractogram, and, as expected, the false-positive bundles are sensibly thinned, that is, reduced volume coverage. The introduction of the additional level in the regularization term of COMMIT2tree further improved these performances. On the one hand, the voxel coverage of the true-positive bundles is comparable but the estimated contributions are larger, which translates into a higher likelihood to be true positives. On the other hand, the contributions of the false positives are further reduced and all of them are almost completely removed. These results were consistent across all the 10 subjects (Table 1).
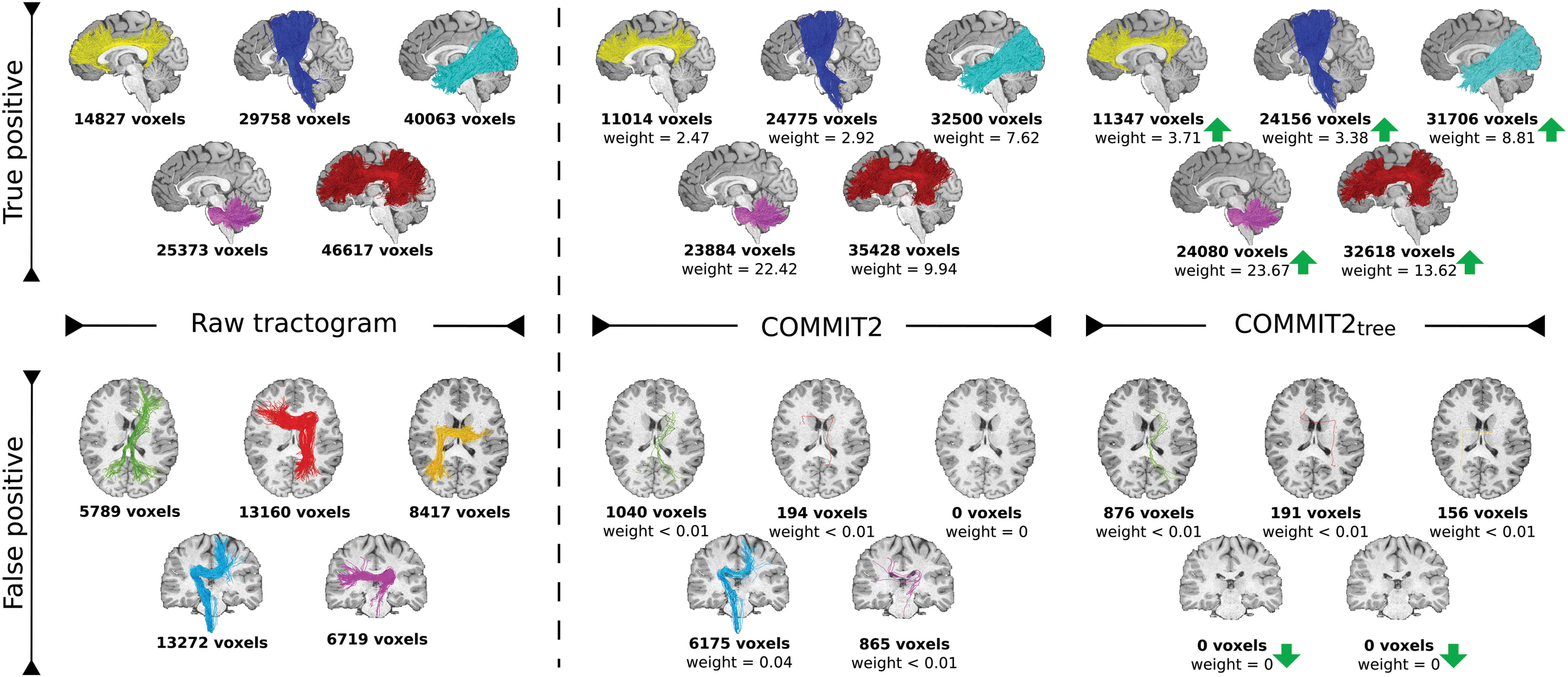
TP and FP bundles estimated in one HCP subject. We report the raw tractogram and after filtering it with COMMIT2 and COMMIT2tree. The voxel coverage is reported for all the bundles, and for the filtered ones, it is also reported the sum of the streamline contributions estimated (weight). The performance of both methods is similar, the TPs keep comparable coverage with the raw tractogram, and the FPs are drastically reduced. Green arrows highlight the cases where COMMIT2tree assigned larger contributions to the TPs and further reduced the FP bundles than COMMIT2. HCP, Human Connectome Project. Color images are available online.
Stability Across the 10 Human Connectome Project Subjects
Voxel coverage (voxels) and estimated sum of streamline contributions (weight) of the five TP bundles and the five FP bundles shown in Figure 5. TPs: #1 (yellow), #2 (dark blue), #3 (cyan), #4 (fuchsia), #5 (dark red). FPs: #1 (green), #2 (light red), #3 (orange), #4 (light blue), #5 (magenta).
COMMIT, Convex Optimization Modeling for Microstructure Informed Tractography; FP, false positive; TP, true positive.
To better appreciate the improved anatomical accuracy of the tractograms, in Figure 6 we visually inspect two representative bundles as reconstructed in each of the 10 subjects. They correspond to the streamlines connecting the medulla oblongata to, respectively, the right precentral gyrus (Bundle 1) and the left precentral gyrus (Bundle 2). As can be seen, these bundles have streamlines that are coherent and well organized in two sub-bundles, and one of these describes an implausible pathway passing through the corpus callosum. The streamlines in the raw tractogram are colored based on their local orientation, whereas for COMMIT2 and COMMIT2tree they are colored according to the estimated sum of the streamline contributions in each sub-bundle. To make this visual comparison more effective, we use two different colormaps, cool for the true-positive streamlines and hot for the false positives. As expected, since COMMIT2 treats all the streamlines in each bundle as a whole, it is not able to completely remove any false-positive sub-bundles that are included in such bundles. However, if subgroups of streamlines are considered in the filtering as done in COMMIT2tree, it is possible to assign low contributions to these false positives or even remove them completely. For all the subjects, there is an evident improvement using COMMIT2tree: lower values are assigned to all the false positives compared with COMMIT2, for example, yellow and green arrows, and in some subjects it was able to completely remove them. Moreover, COMMIT2tree assigned higher contributions than COMMIT2 to all the true positives. We can also appreciate that the patterns of the streamlines of these two bundles are consistent across subjects. Table 2 reports the mean and standard deviation among the 10 subjects of the sum of the streamline contributions and the voxel coverage for each sub-bundle.
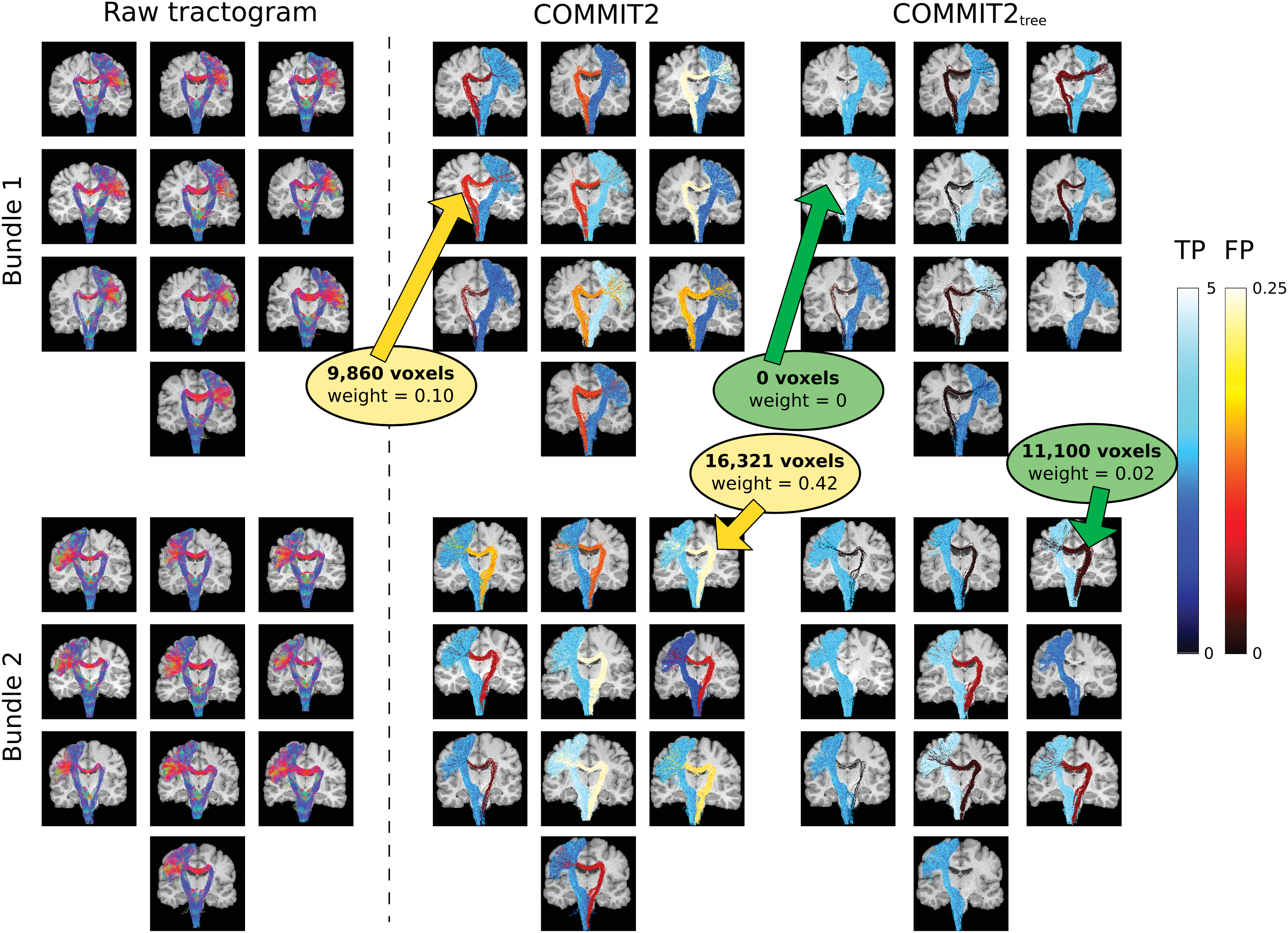
Visual inspection of two representative bundles across the 10 HCP subjects. Reconstructed streamlines belonging to two representative bundles that connect, respectively: medulla oblongata with the precentral gyrus left (Bundle 1), and medulla oblongata with the precentral gyrus right (Bundle 2), as obtained across the 10 subjects before (raw tractogram) and after filtering the tractograms with COMMIT2 and COMMIT2tree. For display and visual inspection purposes, the filtered bundles of streamlines are divided into two sub-bundles, TPs and FPs; these definitions were not used in the filtering procedure. The colormaps display the sum of the streamline contribution per sub-bundle, cool for TP and hot for FP. For all the bundles, the contributions estimated with COMMIT2tree are better than the ones estimated with COMMIT2, the FPs have smaller contributions and the TPs are more significant. The yellow and green arrows point to the region in the raw tractogram with FP sub-bundles after filtering with COMMIT2 and COMMIT2tree, respectively, highlighting their voxel coverage and the sum of the streamline contributions. Color images are available online.
Quantitative Comparison of Sub-Bundle Across the 10 Human Connectome Project Subjects
Mean and standard deviation of the sum of the streamline contributions (weight) and voxel coverage (voxels) for the TP and FP sub-bundles in Figure 6. Bundle 1: Medulla oblongata and precentral gyrus left. Bundle 2: Medulla oblongata and precentral gyrus right.
Figure 7 compares the estimated connectomes in one HCP subject before and after filtering the tractogram reconstructed with probabilistic tractography with COMMIT2 and COMMIT2tree. To assess how well each filtered tractogram explains the input data, we report the input IASF map used for the filtering and the one predicted from the filtered tractograms, using Eq. (1), as well as the root mean square error (RMSE) between the input and the fitted maps. We observe that the connectome of the raw tractogram is very dense (density = 0.896), which is an expected result for probabilistic tracking, whereas after the filtering with COMMIT2 and COMMIT2tree, the connectomes are sparser and have comparable density between them (0.245 and 0.307, respectively). Despite sparser, the corresponding tractograms seem to explain well the input IASF map; the RMSE map is comparable and shows very small errors in the white matter, with higher values located in voxels at the interface with cortical and subcortical gray matter.
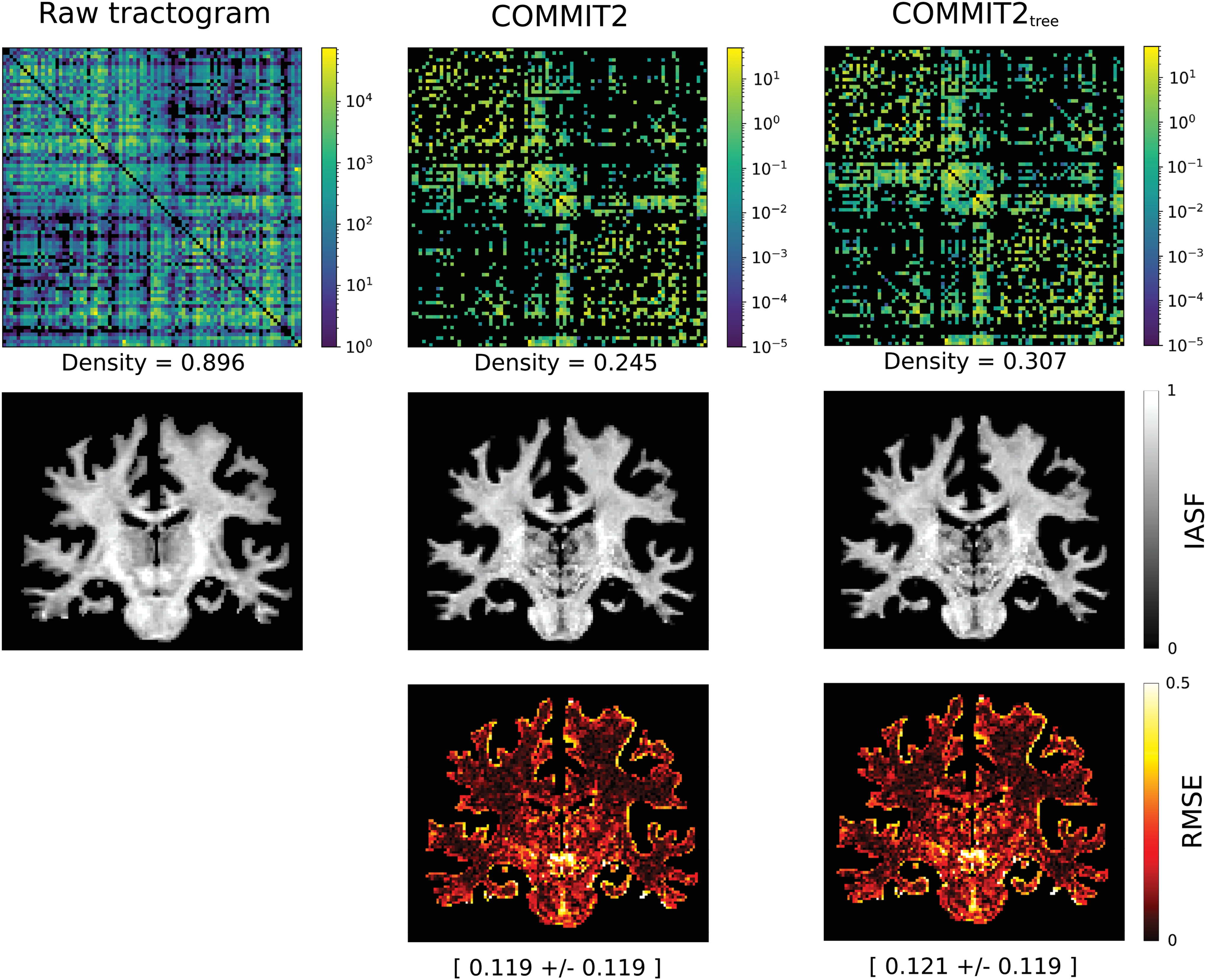
Connectomes estimated in one HCP subject. In the first row, we show the estimated connectomes; in the case of the raw tractograms, we used the number of streamlines, whereas for COMMIT2 and COMMIT2tree, the sum of the streamline contributions was used. In the second row, we present the IASF maps used as input and estimated to perform a visual inspection. The third row presents RMSE maps of the estimations, along with their mean and standard deviation for a quantitative comparison. IASF, intra-axonal signal fraction; RMSE, root mean square error. Color images are available online.
Discussion
Tractography is a unique technique that is able to describe the major neural pathways in the white matter. In the last decades, it has been used to study the structural network organization of the brain, but its anatomical inaccuracy and limitations have been exposed in different studies (Drakesmith et al., 2015; Girard et al., 2020; Maier-Hein et al., 2017; Rheault et al., 2020b; Schilling et al., 2019; Thomas et al., 2014; Zalesky et al., 2016). One of the major limitations still unresolved is the intrinsic trade-off between specificity and sensitivity. In a recent international challenge (Maier-Hein et al., 2017), it was suggested that a possible way to overcome this issue would be to inject microstructural and anatomical prior during tractography reconstruction. A step forward in this direction was achieved by the recent work of Schiavi et al. (2020), where microstructure informed tractography was combined with the anatomical prior that white matter fibers (axons) are organized in fascicles giving origin to COMMIT2. Using the prior knowledge that bundles are organized in sub-bundles (Chenot et al., 2019; Mandonnet et al., 2018), in this work, we extended COMMIT2 proposing the use of a multilevel organization of the streamlines to identify streamlines following implausible trajectories inside VB. This new formulation, called COMMIT2tree, considers partitions of the streamlines with different granularity and organizes them in a hierarchical structure, which provides more degrees of freedom than COMMIT2 but in a constrained clever way. The subgroups used in COMMIT2tree share anatomical constraints such as the starting and ending region in the gray matter, but they have different geometrical properties, that is, the trajectory. Figure 6 clearly shows that by considering this way of grouping in the optimization, more false-positive streamlines inside true-positive bundles are removed, boosting further the accuracy of the final tractogram.
In synthetic data, the connectivity graphs obtained after filtering the tractograms with COMMIT2 and COMMIT2tree showed comparable results. However, the simple geometry of this phantom does not allow us to fully exploit the advantages of our proposed multilevel regularization, as the bundles consist of regular and smooth tubes. Nonetheless, our findings indicate that this enhanced formulation results in a stable and robust regularization of the original problem. In particular, Figure 4 shows that adding more levels to the structure does not affect the stability of the method, but can slightly improve the quality of the estimated tractograms.
To evaluate the quality of the in vivo reconstructions, we analyzed 10 HCP subjects and we looked both at the anatomical accuracy of individual bundles and at the overall structural connectivity estimated after filtering the tractograms with COMMIT2 and COMMIT2tree. The density of the filtered connectomes was about one-third of the unfiltered one, showing again similar performances of both methods. These results confirm and extend the results presented in Schiavi et al. (2020), which were in agreement with the theory of the economy of brain networks (Bullmore and Sporns, 2012). However, when looking more closely at the individual bundles, before and after the filtering, it is evident that a significant improvement was achieved by COMMIT2tree. In fact, in Figure 5 and Table 1 we can appreciate how COMMIT2tree was able to filter more the false-positive bundles than COMMIT2 without negatively affecting the true positives, in particular because of its capability to remove invalid streamlines inside VB (Fig. 6; Table 2). From a quantitative point of view, we observe that both methodologies assigned lower contributions to the false positives than the true positives; notably, it is important to note that COMMIT2tree assigned higher contributions to the true positives and smaller values to the false positives compared with its predecessor. This is an important property when one is interested in network properties relying on connection strengths.
Although COMMIT2tree was able to further improve the anatomical accuracy of the tractograms, it did not eliminate completely all known false positives in every analyzed subject. This may be partly due to the regularization parameter
It is worth noting that using a finer partitioning of the streamlines may create groups that might capture better the plausible and implausible sub-bundles; for instance, in the extreme case where all groups have a cardinality equal to 1, that is, they consist of a single streamline, true- and false-positive streamlines are clearly separated in distinct groups. However, as the granularity of this partitioning becomes smaller, the number of generated groups increases accordingly. However, since the groups are treated as independent entities in the penalty term in Eq. (4), this process has the side effect of gradually reintroducing the ambiguous configurations that, instead, COMMIT2 was able to disentangle using a coarser partition. Supplementary Figure S2 shows that, indeed, the performances gradually deteriorate as the groups become smaller. As it is not possible to predict a priori the behavior of the algorithm using partitions with different granularity, our proposed approach represents an effective way to consider and take advantage of more than one granularity in the filtering procedure. Clearly, future research will be required to identify the optimal configuration of groups in this multilevel structure.
Finally, it is important to note that the proposed approach is still a filtering procedure, and so, even if it can be applied on any type of input tractograms (coming from either deterministic or probabilistic tractography approaches), it is still limited on the quality of the input reconstruction. Clearly, if a bundle is not present in the original tractograms, it will not be recovered by COMMIT2tree. So, particular attention should be given to this aspect. Another important factor that affects the results is the selection of the regularization parameter
Conclusion
The anatomical accuracy of tractography has been heavily challenged in the past few years and seriously questioned the use of this technique for mapping reliably the structural connectivity of the brain. Our group recently developed a postprocessing filtering procedure called COMMIT2 that allowed a dramatic increase in the accuracy of the reconstructions. However, this formulation did not consider the fact that fibers in the brain are anatomically organized in fascicles and subfascicles. In this work, we showed that taking into account this multilevel organization of the fibers allows considering information about the white matter structure in a more specific way. This improved formulation, that is, COMMIT2tree, not only reduces the number of false-positive connections in the estimated tractograms as COMMIT2 but it is also able to filter out implausible streamlines within true-positive bundles, boosting even further the quality of the reconstructions. Our new formulation represents an additional step forward to improve the anatomical accuracy of the tractograms and our understanding of how different brain regions are interconnected.
Footnotes
Code and Data Availability
The numerical phantom used as validation is publicly available and can be downloaded from
Acknowledgments
Data were provided by the Human Connectome Project, WU-Minn Consortium (Principal Investigators: David Van Essen and Kamil Ugurbil; 1U54MH091657) funded by the 16 NIH Institutes and Centers that support the NIH Blueprint for Neuroscience Research; and by the McDonnell Center for Systems Neuroscience at Washington University.
Authors' Contributions
All the authors, M.O.-P., S.S., F.R., G.G., L.P., M.D., and A.D., contributed significantly to the work. M.O.-P., S.S., and A.D. conceptualized the problem. M.O.-P., S.S., G.G., and A.D. developed, implemented, and tested the technical framework. L.P. manually segmented the in vivo VB and IB toward which we compared our method. M.O.-P., F.R., and M.D. conceptualized and developed the in vivo validation framework. M.O.-P., S.S., F.R., G.G., L.P., M.D., and A.D. wrote the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Rita Levi Montalcini Programme for young researchers of the Italian Ministry of Education, University and Research.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Appendix
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.