
Abstract
Introduction:
After chronic impairment of the right dominant hand, some individuals are able to compensate with increased performance with the intact left nondominant hand. This process may depend on the nondominant (right) hemisphere's ability to access dominant (left) hemisphere mechanisms. To predict or modulate patients' ability to compensate with the left hand, we must understand the neural mechanisms and connections that underpin this process.
Methods:
We studied 17 right-handed healthy adults who underwent resting-state functional connectivity (FC) magnetic resonance imaging scans before 10 days of training on a left-hand precision drawing task. We sought to identify right-hemisphere areas where FC from left-hemisphere seeds (primary motor cortex, intraparietal sulcus [IPS], inferior parietal lobule) would predict left-hand skill learning or magnitude.
Results:
Left-hand skill learning was predicted by convergent FC from left primary motor cortex and left IPS onto the same small region (0.31 cm3) in the right superior parietal lobule (SPL).
Discussion:
For patients who must compensate with the left hand, the right SPL may play a key role in integrating left-hemisphere mechanisms that typically control the right hand. Our study provides the first model of how interhemispheric functional connections in the human brain may support compensation after chronic injury to the right hand.
Impact statement
This article presents the first model of how the human brain applies left-hemisphere (dominant hand [DH]) specializations to support the right hemisphere for compensatory action with the nondominant hand (NDH). Compensation with the NDH is critical for the rehabilitation after many neurological disorders that lead to irreversible impairment of the DH (e.g., stroke, amputation, nerve injury), but no effective therapies exist to promote compensation because the neural mechanisms remain unknown. Therefore, this article will open up new directions in rehabilitation neuroscience by presenting a testable model of the connections that underpin much needed (and currently nonexistent) therapies.
Introduction
Individuals who suffer from chronic unilateral impairment of their dominant hand (DH) must use their intact nondominant hand (NDH) to maintain self-sufficiency (“compensation” [Jones, 2017]), but it remains unknown how connectivity in the human brain supports this process. Chronic unilateral impairment of the DH can occur after a variety of clinical conditions, including peripheral nerve injury and amputation. The United States has 20 million peripheral nerve injury patients (Lundborg, 2003), and ∼166,000 surgical treatments occur every year to treat peripheral nerve injury of the right upper extremity (Brattain, 2013; Kouyoumdjian et al., 2017; Philip et al., 2017; Taylor et al., 2008). Thirty-three percent to 39% of upper extremity nerve injury patients never achieve satisfactory motor recovery after surgical repair, regardless of any rehabilitation they receive (Council, 1943; Dyck et al., 2005; He et al., 2014). This leaves 55–66,000 peripheral nerve injury patients per year who are not adequately served by rehabilitation strategies that focus on restoring lost function (Langhorne et al., 2009). To help these patients, we must understand the neural mechanisms and connections that enable compensation after DH injury.
Compensation is difficult, but possible. In right-handed humans, the DH is specialized for intersegmental dynamics and precision movement (Sainburg and Kalakanis, 2000; Sainburg, 2002), so a key challenge is achieving NDH performance at precision movements (e.g., writing, drawing). Amputees who lose their DH eventually achieve the ability to perform skillful precision movements with the intact NDH (Philip and Frey, 2014), but this may require decades of exclusive forced use of the NDH. Healthy adults can improve NDH precision performance with as little as 10 days of training (Philip and Frey, 2016), but other studies have shown marked interindividual differences in the ability to acquire NDH skill. NDH handwriting studies in healthy adults (Walker and Henneberg, 2007) and amputees (Yancosek and Mullineaux, 2011) show great interindividual variability, which suggests that some individuals may be better able to achieve NDH performance than others.
The neural mechanisms of NDH compensation are likely to involve interhemispheric communication in the cerebral cortex. The DH—and thus the left hemisphere—contains specializations that support precision movement with both hands (Mani et al., 2013; Sainburg and Kalakanis, 2000; Sainburg, 2002; Schaefer et al., 2009) (For clarity, we refer to right-handed individuals only, so the DH is supported by the contralateral left hemisphere.). For left-hemisphere specializations to support ipsilateral NDH movement, those specializations must involve the minority (10–25%) of uncrossed descending pyramidal fibers (Davidoff, 1990), or interhemispheric connections to right-hemisphere motor areas with direct control of the NDH. Interhemispheric connections are likely to play the primary role in precision NDH movement: the timing of interhemispheric connections indicates a functional rather than epiphenomenal role (Fiori et al., 2017), and interhemispheric coordination depends on task demand (Wischnewski et al., 2016). Interhemispheric communication for motor control is primarily transcallosal (Doron and Gazzaniga, 2008), especially in the form of interhemispheric inhibition (Ferbert et al., 1992; Hortobágyi et al., 2011; Kobayashi et al., 2003; Pelled et al., 2009) that arises from transcallosal connections between left and right primary motor cortex (M1). However, transcallosal connectivity is not dominated by motor cortices: callosotomy patients show greater interhemispheric disconnection between parietal regions than between primary sensorimotor regions (Roland et al., 2017).
Our goal was to identify the interhemispheric neural mechanisms in the human brain that support people's ability to improve NDH performance at a precision drawing task (PDT). To achieve this, we re-evaluated data from an existing study of NDH precision movement training in healthy adults (Philip and Frey, 2016), to identify pretraining functional connectivity (FC) patterns that predicted individuals' later ability to improve their NDH performance. Participants received resting-state functional magnetic resonance imaging (MRI) scans before 10 days of NDH training (∼18 min/day). We hypothesized that interhemispheric communication involving left-hemisphere primary motor cortex hand area (M1-Hand, or M1 for brevity), intraparietal sulcus (IPS), or inferior parietal lobule (IPL) would predict final NDH performance or magnitude of NDH learning. We found that interhemispheric connectivity between left M1 and IPS onto the same small region in right superior parietal lobule (SPL) predicted subsequent ability of healthy adults to improve their NDH performance.
Materials and Methods
Experimental design overview
Full methods have been previously published in Philip and Frey (2016). In brief, participants used their left NDH to practice the PDT for 10 days or until their NDH performance reached an a priori criterion of ≥80% of their baseline DH speed on two consecutive training days. DH baseline performance was recorded before and after NDH training. Participants underwent MRI scanning before the first PDT session and 1 day after the final PDT session. This study investigated relationships between pretraining MRI and two outcome variables: peak PDT performance and magnitude of PDT learning.
Data collection was performed with the approval of the University of Missouri Institutional Review Board and completed in accordance with the Declaration of Helsinki. These data are not associated with a Clinical Trials Registration Number because they were collected in 2012–2013.
Participants
We excluded individuals who were musicians, knitters, or had other hobbies involving precision bimanual skills. Twenty-two participants completed the behavioral study (age 29 ± 11, 15 female). All were right handed (Edinburgh scores 90 ± 16; Oldfield [1971]). Three participants were excluded from detailed analyses due to noncompliance (see Philip and Frey, 2016, for details). One participant did not complete the MRI study, and another participant's MRI data were discarded due to excess motion (for details, see MRI Data Analysis below), leaving 17 participants for this study (age 27 ± 8, 12 female).
Precision drawing task
Participants used a pen stylus to draw a line through each stimulus while remaining within the boundaries of mirror-symmetrical geometric forms (Fig. 1). The PDT was presented on a Cintiq 12wx tablet with integrated 1200*800 screen (Wacom Co., Otone, Japan), controlled by Presentation software, v.16.0 (Neurobehavioral Systems, Inc., Albany, CA). This system allowed participants to draw directly on the tablet screen, during which endpoint speed and position were recorded at 30 Hz.
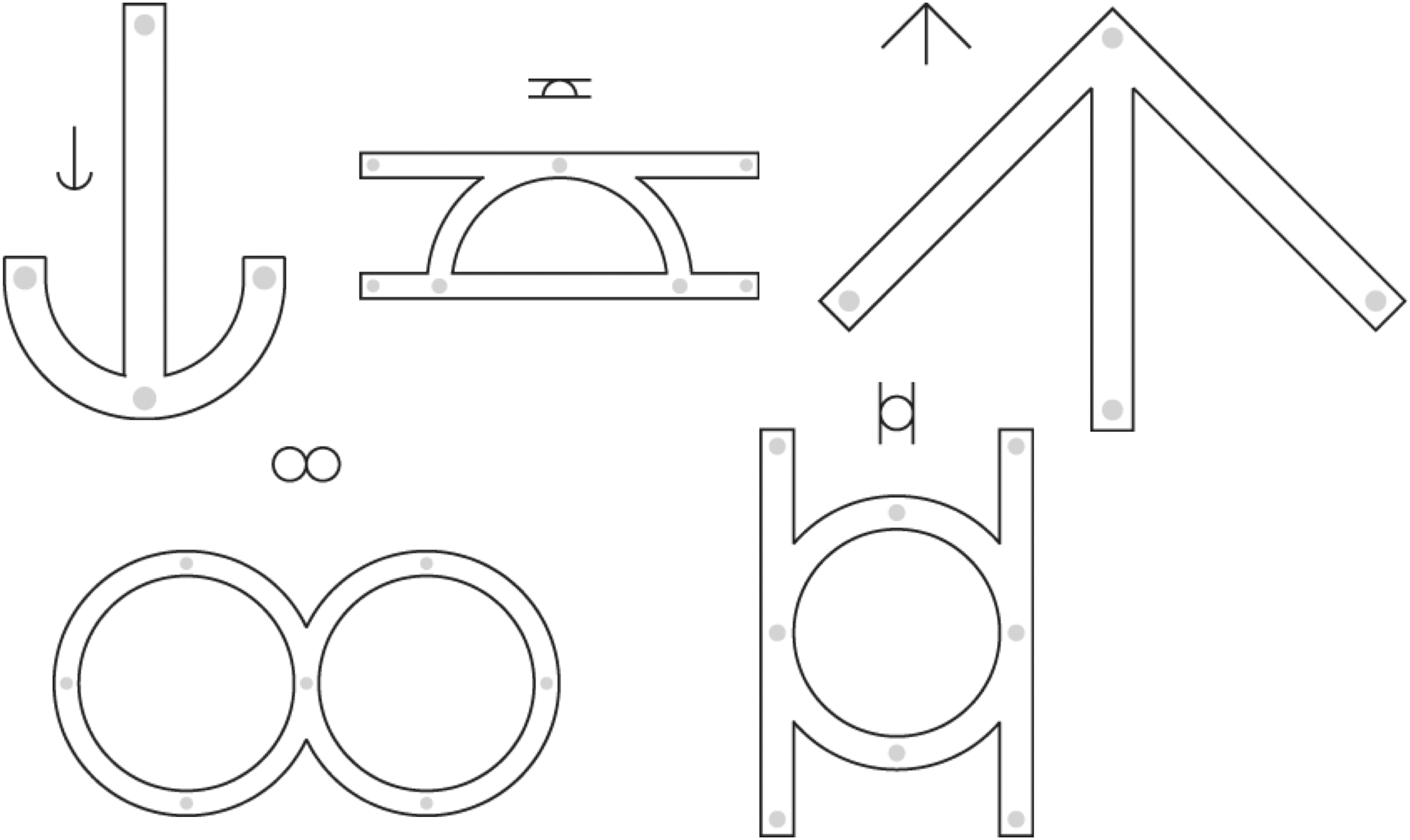
Precision drawing task. Five sample stimuli shown, out of 45 possible.
Each stimulus was built from 2, 3, or 4 geometric elements (lines or semicircles); each element had a drawing length of 45 mm, and thus total path lengths of 90, 180, or 360 mm. Precision requirements were manipulated by varying stimulus width (3, 4, or 5 mm). The full set of possible PDT stimuli entailed 15 shapes (5 of each length), each presented at all 3 widths, producing a total of 45. NDH training sessions entailed 90 trials, 2 repetitions of all 45 stimuli. DH sessions entailed 2 repetitions of 12 shapes (4 of each length) at 2 widths (3 and 5 mm), producing a total of 48 stimuli. Trials were presented in a unique pseudorandom order during each session.
Participants were instructed to move as quickly as possible without making any errors; that is, they were instructed to prioritize error minimization over speed. A trial was defined as an “error” if the drawn lines ever passed outside the stimulus margin; except for distinct accidental lines created as part of raising or lowering the pen, and for overshoots at the end of segments (following Beery and Beery, 2004). A trial was considered “incomplete” if the drawn lines did not pass fully through all open paths within the form. A trial was considered a “success” if it was neither an error nor incomplete. Both successful and error trials were used for further analyses, to keep trial counts constant across participants and training sessions.
To initially familiarize subjects with the task requirements, a DH practice session preceded the DH1 session, and a NDH practice session preceded the NH1 session. Each of these familiarization sessions consisted of 15 trials (15 shapes, 1 repetition, all at 4 mm width).
Because the number of trials per session was fixed, session duration decreased over time as participants became more skilled and increased their movement speed. NDH training session #1 took 21 ± 3 min, while the final session took 17 ± 3 min. Across all training sessions and participants, total training time averaged 182 ± 30 min.
Behavioral analysis
The tablet collected x and y positions of the pen endpoint at 30 Hz. During analysis, velocity profiles for each trial were smoothed using an acausal Gaussian filter with sigma of three samples (90 ms).
Ratings of trial status (success, error, or incomplete) were performed by hand. A senior experimenter's ratings were validated by a second experimenter. Raters were not blinded, because data were rated on the day of collection. For any session in which within-session agreement was <0.93 or inter-rater reliability (IRR) was <0.7 (mean −0.5 standard deviation [STD] from Philip and Frey [2014]), the two experimenters rerated the session collaboratively until they reached consensus on every trial. Final ratings reached 0.98 ± 0.01 agreement and 0.91 ± 0.04 IRR across participants.
This study used endpoint movement smoothness (“smoothness” for brevity) as its outcome measure, based on previous findings that smoothness provided the most hand-specific performance gains during this task (Philip and Frey, 2016), and because movement smoothness best captures the DH-specific (left-hemisphere) specializations for feedforward control and precision smooth movement (Sainburg and Kalakanis, 2000; Sainburg, 2002).
Smoothness was calculated as −1 * the number of velocity peaks per 45 mm shape element, average for each trial. Higher smoothness (closer to zero) represents fewer submovements per movement. To measure performance changes consistently across participants, smoothness was normalized to each participant's first NDH session by converting it into a “ZNDH” score. A ZNDH score was calculated for each participant and session by taking participant mean smoothness during that session, subtracting the participants' mean during their first NDH session, and dividing by the STD during their first NDH session. In other words, the ZNH score reflects a Z-score based on the participant's untrained NDH performance; this is similar to Cohen's d measure of effect size (Cohen, 2013), but based only on the STD during the first NDH session.
MRI data collection
Scans were performed on a Siemens (Erlangen, Germany) 3T Trio using a standard birdcage radiofrequency coil. During each fMRI session, participants lay still with their eyes open. Scan sessions included the following: (1) T1- and T2-weighted structural scans, (2) three 5:42 min functional runs using echoplanar imaging sensitive to the blood oxygen-level dependent contrast (BOLD-EPI), and (3) a gradient echo field map.
BOLD scans used a T2*-weighted sequence with the following parameters: temporal resolution (TR) = 2240 ms, echo time (TE) = 30 ms, 64 × 64 voxel matrix, field of view (FoV) = 256 mm, 36 contiguous axial slices acquired in interleaved order, thickness = 4.0 mm, in-plane resolution = 4.0 mm, bandwidth = 2004 Hz/pixel, 150 volumes. The initial two volumes in each scan were discarded to allow steady-state magnetization to be approached. Throughout the functional scans, participants were instructed to remain awake with their eyes open and maintain fixation on a centrally presented point. Compliance was manually monitored using an Eye-Trac 6000 (Applied Science Laboratories, Bedford, MA) eye-tracking camera.
High-resolution T1-weighted structural images were also acquired, using the 3D MP-RAGE pulse sequence: TR = 2500 ms, TE = 4.38 ms, inversion time = 1100 ms, flip angle = 8.0°, 256 × 256 voxel matrix, FoV = 256 mm, 176 contiguous axial slices, thickness = 1.0 mm, in-plane resolution: 1.0 × 1.0 mm.
DICOM image files were converted to NIFTI format using MRI-Convert software.
MRI preprocessing
Image preprocessing was performed with 4dfp Tools. Image preprocessing included the following steps: (1) compensation for slice-dependent time shifts, (2) elimination of odd/even slice intensity differences due to interleaved acquisition, and (3) realignment of all data acquired in each subject within and across runs to compensate for rigid body motion (Ojemann et al., 1997). The functional data were transformed into Talairach atlas space (Talairach and Tournoux, 1988) by computing a sequence of affine transformations (first frame of BOLD run to T2-weighted image to MP-RAGE to atlas representative target without compensation for local distortions between echo planar images and anatomy), which were combined by matrix multiplication, resampling to a 2 mm isotropic grid. For crossmodal (i.e., functional to structural) image registration, a locally developed algorithm was used (Rowland et al., 2005).
Resting-state FC models
Image analysis was performed with FIDL (Functional Independent Data Language;
Subject-specific general linear models (Friston et al., 1995) were fit to BOLD time series at each voxel as shown schematically below: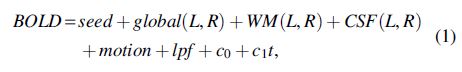
where the notation (L,R) represents 2 terms. Head movement was accounted for by motion, 24 time series formed from measured head shifts and angular displacements in three dimensions (i.e., X, Y, Z, pitch, yaw, and roll) along with their squares, derivatives, and squared derivatives (Friston et al., 1996). The low-pass filter lpf as implemented with a Fourier basis set that consisted of sine and cosine pairs modeling full cycles from the cutoff of 0.08 Hz to the Nyquist frequency (Biswal et al., 1995; Lowe et al., 1998), thus low-pass filtering was accomplished with the inclusion of high-frequency terms. Additional terms included the constant c 0 and linear trend c 1 t to model slow drifts. The partial correlation model of Eq. 1 was solved through ordinary least squares, yielding estimated weights for all regressors.
Seed selection
For the purposes of analysis, 5 mm-radius spherical seed regions were defined in left M1, left IPS, and left IPL. The M1 seed was based on normative sensorimotor hand area (Philip and Frey, 2014). The parietal seeds were based on previous analyses of the current dataset (Philip and Frey, 2016), and restricted to volume defined as ≥25% chance of being in the appropriate anatomical area according to the Juelich Histological Atlas (Eickhoff et al., 2006, 2007). Within left IPS, the voxel with peak activation was chosen from areas that previously showed interhemispheric changes related to learning—specifically, where training-related increases in FC with right M1 correlated with NDH skill learning (see fig. 4a in Philip and Frey, 2016). Within left IPL, the voxel with peak activation was chosen from areas that previously showed interhesmipheric changes related to learning and retention—specifically, where training-related changes in FC with right M1 predicted long-term retention of NDH skill (see fig. 5a in Philip and Frey, 2016). These selected voxels' Montreal Neurological Institute coordinates were as follows: left M1 X = −38, Y = −24, Z = 54; left IPS X = −51, Y = −41, Z = 43; left IPL X = −55, Y = −22, Z = 26. In all three cases, the seed was created as a 5 mm-radius sphere centered on the peak voxel, and limited to the volume with ≥25% chance of being in the appropriate anatomical area.
On a post hoc basis (after the other analyses described here), a fourth seed region was defined in left dorsal premotor cortex (PMd) to determine whether PMd–PMd interhemispheric connectivity contributed to learning of NDH drawing performance. The PMd seed's central voxel was selected as the peak voxel of drawing-specific activity in left PMd in previous studies (Potgieser et al., 2015) at X = −24, Y = −8, Z = 50. The seed was created as a 5 mm-radius sphere centered on that central voxel, and limited to the volume with a ≥ 10% chance of being in the appropriate anatomical area. This cutoff was lower than the other seeds, because the PMd peak voxel had only a 21% chance of being in Brodmann's Area 6 according to the Juelich Histological Atlas (Eickhoff et al., 2006, 2007), which made a 25% cutoff unfeasible.
Correlation of behavior with resting-state FC
Separate FC models were computed for seeds in left M1, left IPS, and left IPL (see Eq. 1). For each participant, the estimated voxelwise weights of the seed region were normalized to the constant c 0, then smoothed with a 4 mm full width at half-maximum three-dimensional Gaussian kernel to blur individual differences in brain anatomy. The Pearson correlation was then computed between each participant's behavior (see next paragraph) and their measure of FC for each seed, respectively. The correlation maps were algebraically converted to T statistics, then fit to a normal distribution yielding gaussianized T statistics, which were corrected for multiple comparisons (|z| ≥ 3.0, minimum 21 face-connected voxels, p < 0.05 corrected) with a Monte Carlo-based method (Forman et al., 1995; McAvoy et al., 2001).
For this analysis, four measures of behavior were used: “learning,” defined for each participant as the increase in movement smoothness between their initial session and the session with peak smoothness (i.e., maximum ZNH); “baseline,” defined as pretraining NDH smoothness; “forgetting,” defined as the peak NDH smoothness minus NDH smoothness at 6 months post-training; and “retention,” defined as NDH smoothness 6 months post-training. Behavioral results have been previously published in detail (Philip and Frey, 2016), but key values used for the current analysis are shown in Table 1.
Key Behavioral Results from Philip and Frey 2016 Used for Analysis Here
All values represent movement smoothness. Baseline = −1 * velocity peaks per 45 mm segment; learning, forgetting, and retention are in ZNDH (Z-score relative to baseline day). Missing data = participant lost to 6-month follow-up.
NDH, nondominant hand.
Results
Convergent connectivity onto right SPL predicts NDH learning
NDH smoothness learning was correlated with interhemispheric FC from left M1 seed onto a scattered selection of right-hemisphere areas, shown in Figure 2, including SPL, inferior frontal gyrus extending into insula, and multiple areas in the basal ganglia. Smoothness learning was also correlated with interhemispheric FC between left IPS and a selection of right-hemisphere areas, shown in Figure 3, including SPL, precuneus, cingulate sulcus, insula, lateral occipital cortex, and V7. We found no areas where NDH smoothness learning was correlated with FC from our left IPL seed. Because of the exploratory nature of this analysis, we do not delve broadly into the scattered areas showing FC with individual seed regions, though we provide the full data (including left-hemisphere connectivity, that is, intrahemispheric) in Table 2. Instead, we focus on convergent patterns of FC across analyses.
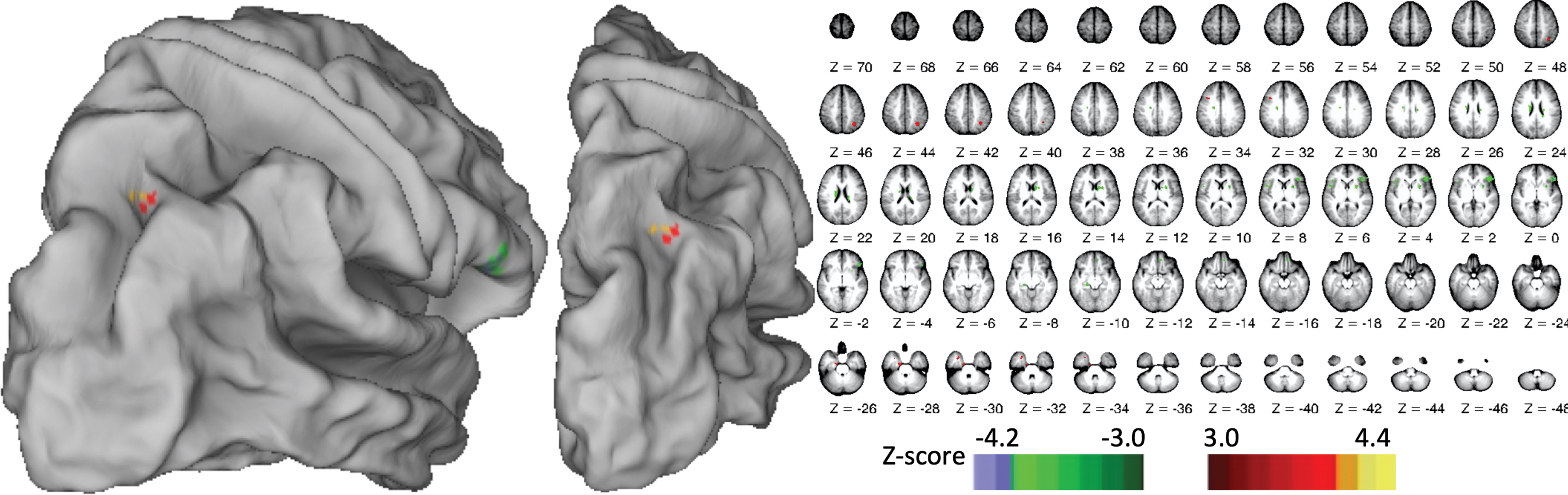
Right-hemisphere areas where FC with left M1 seed predicted NDH smoothness learning. FC, functional connectivity; NDH, nondominant hand. Color images are available online.
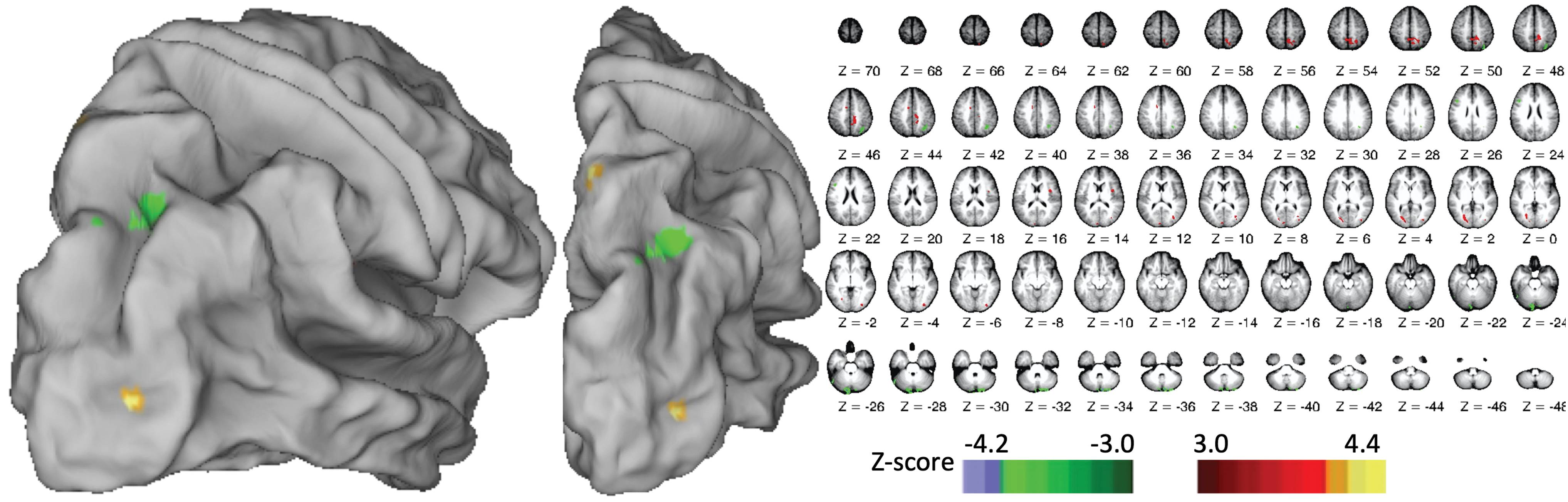
Right-hemisphere areas where FC with left IPS seed predicted NDH smoothness learning. IPS, intraparietal sulcus. Color images are available online.
Clusters with Learning-Predictive Connectivity with Left-Hemisphere Seeds
Interhemispheric connectivity (right hemisphere) data are visualized in Figures 2 + 3. Z-score = gaussianized T of correlation between seed timecourse and participant learning scores, at center of gravity (CoG).
Values reported for peak instead of CoG due to concave cluster.
IPS, intraparietal sulcus; SPL, superior parietal lobule.
We found one convergence across our analyses: the learning-predictive interhemispheric connectivity from left M1 (Fig. 2) and left IPS (Fig. 3) both included the same area in right SPL, as illustrated in Figure 4 and quantified in Table 3. The “intersection volume” shared between the two clusters was 0.31 cm3 (46% of the M1-linked cluster, 22% of the IPS-linked cluster).
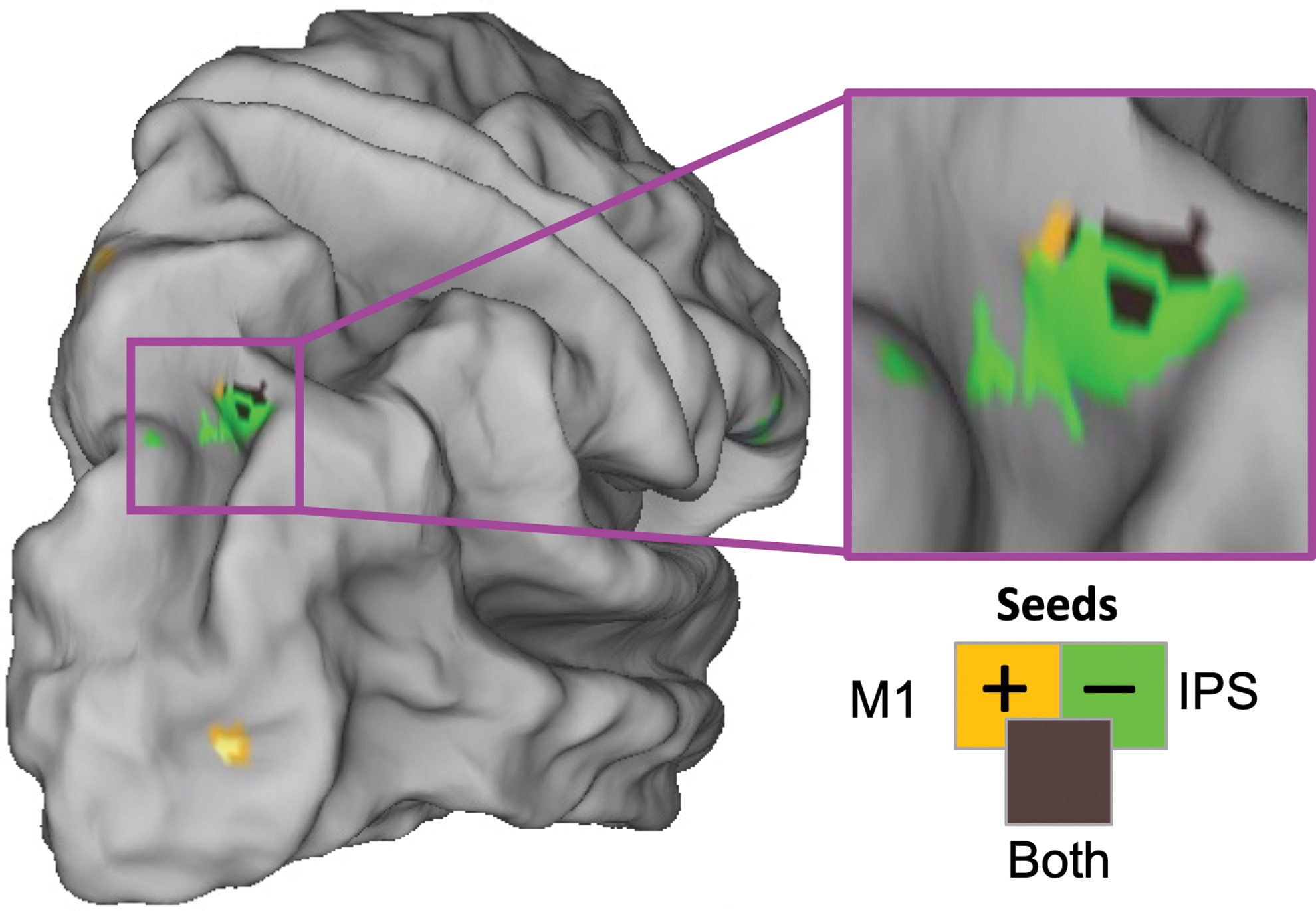
Learning-predictive interhemispheric connectivity from left M1 and left IPS converges on the same area of right SPL. SPL, superior parietal lobule. Color images are available online.
Overlapping Clusters in Right SPL Show Functional Connectivity from Left-Hemisphere Seeds
We found no areas where interhemispheric connectivity from left-hemisphere seeds predicted NDH baseline, NDH forgetting, or NDH retention.
Predictive connectivity is stable across training
To determine whether the “intersection volume” in Figure 4 represented connectivity that is stable across training, we compared it with our previously identified network of areas that show training-related changes in FC from M1 in either hemisphere (see fig. 4b in Philip and Frey, 2016), with both maps thresholded at p < 0.001. We found zero overlap between voxels in the “intersection volume” and voxels with training-related changes in M1 connectivity. Therefore, the pattern of interhemispheric FC that predicted subsequent NDH learning (Fig. 4) appears to be stable across time and the learning of relevant motor skills.
Left M1 and IPS project onto right SPL through two separate networks
Based on our results thus far, it remains unclear whether the FC from left M1 and left IPS onto right SPL (the “intersection volume”) reflects separate connections or an indirect connection (e.g., from left M1 to left IPS to right SPL). To address this, we assessed whether the two seed regions showed FC with each other. The two seed regions were not functionally connected: for each seed, the network of significantly connected voxels did not contain the other seed, as shown in Supplementary Figure S1. To further address the separability of M1-related and IPS-related FC, we identified the network of areas with differential connectivity to each seed region. Bilateral somatosensory–motor areas were preferentially connected to left M1, whereas areas involved in attention (including frontal eye fields, inferior frontal gyrus, and inferior frontal sulcus) and higher order visuomotor control (PMd) were preferentially connected to left IPS, as illustrated in Figure 5. Overall, this suggests that left IPS and left M1 functionally connect to right SPL as part of different networks.
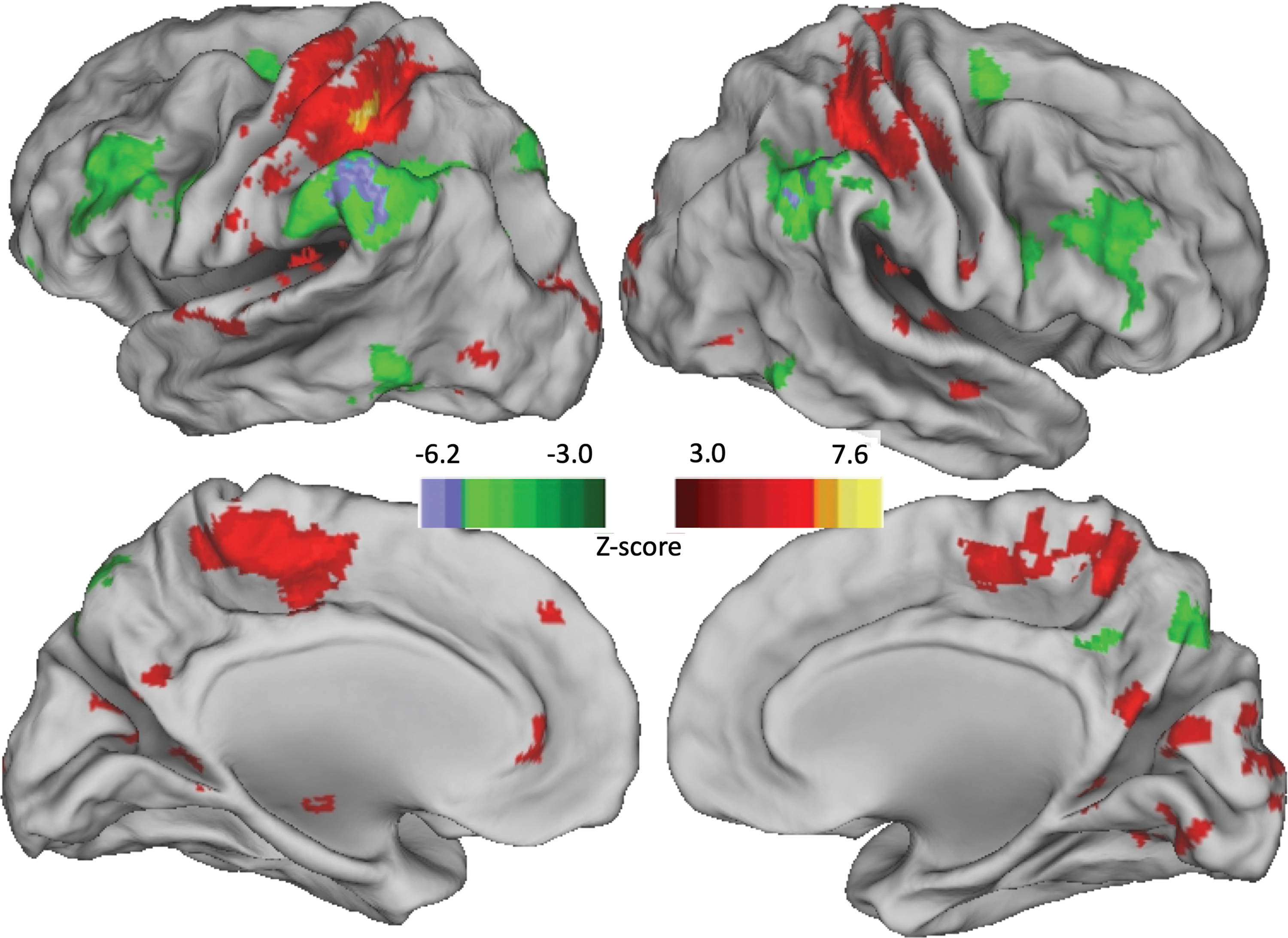
Areas where FC differed between left M1 seed and left IPS seed. Z-scores indicate correlation with “M1 minus IPS,” so positive values = M1, negative values = IPS. Color images are available online.
Post hoc analysis: PMd–PMd interhemispheric connectivity does not predict NDH learning
After seeing the above results, we performed a post hoc FC analysis to determine whether PMd–PMd interhemispheric connectivity supported NDH learning, based on previous results that showed a specific role of bilateral PMd in drawing and right PMd in NDH drawing (Potgieser et al., 2015). Our results did not support the hypothesis that PMd–PMd interhemispheric connectivity channeled left-hemisphere information to the right hemisphere: right PMd (defined as the contralateral homolog of our left PMd seed) was not among the areas with smoothness-learning-correlated FC from left PMd, as shown in Supplementary Figure S2. However, the pattern of functional connections from the left PMd included an area at the left middle frontal gyrus (z = 34), which almost coincides with the left middle frontal gyrus area connected from the left M1 seed (z = −33.2, Table 2 and Fig. 2). Left PMd was also functionally connected to the right-hemisphere homolog of this middle frontal gyrus area, which could potentially indicate an indirect contribution of left PMd to interhemispheric interaction through the bilateral middle frontal
Discussion
We explored FC patterns in the healthy adult human brain, to identify interhemispheric connectivity that predicted later PDT skill with the left NDH. We found that subsequent ability to learn the NDH drawing task was correlated with convergent FC between two of our left-hemisphere seeds (primary motor cortex hand area, M1; and IPS) and a restricted region of the right SPL. In contrast, we found no interhemispheric connectivity that predicted NDH initial or maximum performance—only NDH learning (magnitude of changes across training). This suggests that interhemispheric connectivity from left-hemisphere areas onto right SPL may specifically predict individuals' ability to learn a precision, DH-lateralized skill with the NDH.
Model of interhemispheric connectivity to support NDH skill learning
In typical right-handed adults, the dominant right hand (and left cerebral hemisphere) is specialized for coordination of muscle forces across joints, which are critical for feedforward control and precision smooth movement (Sainburg and Kalakanis, 2000; Sainburg, 2002). These left-hemisphere specializations support precision movement with both hands, as demonstrated by the pattern of bilateral impairments found in stroke patients with unilateral lesions (Schaefer et al., 2007). These left-hemisphere specializations are likely to support ipsilateral NDH movement through interhemispheric cortical connections, rather than only through uncrossed descending pyramidal fibers. Interhemispheric transcallosal connections (Doron and Gazzaniga, 2008) play a role in coordinating bilateral movements (Kennerley et al., 2002), with greater involvement for more difficult tasks (Buetefisch et al., 2014; Hummel et al., 2003; Wischnewski et al., 2016). This study provides the first model of how interhemispheric connections may recruit left-hemisphere specializations to support the ipsilateral NDH for learning of a precision feedback-driven task, as shown in Figure 6.

Model of interhemispheric connectivity supporting learning of precision skill with the left NDH.
According to our model in Figure 6, NDH learning is supported by interhemispheric connectivity from left M1 and/or IPS onto right SPL. Right M1 must also be involved for descending corticospinal control of the left NDH (Lawrence and Kuypers, 1968; Trevarthen, 1965), and SPL-M1 connections have been well established in monkeys and humans (Chen et al., 2003; Cohen and Andersen, 2002; Dejerine and Dejerine-Klumpke, 1895; Koch et al., 2007; Mountcastle et al., 1975). Each of these left-hemisphere areas may be capable of interhemispheric connections. M1 interhemispheric connections are better known (Ferbert et al., 1992; Hortobágyi et al., 2011; Kobayashi et al., 2003; Meyer et al., 1998; Pelled et al., 2009), but they do not fully explain interhemispheric connectivity: callosotomy patients show greater loss of IHC between parietal regions than between primary sensorimotor regions (Roland et al., 2017), and we found no correlations between M1-M1 FC and NDH learning.
Each of our three key areas (left M1, left IPS, right SPL) have well-established roles in control of precision movements. Left M1 has a well-established transcallosal role in motor control, both direct (Buetefisch et al., 2014; Bundy et al., 2018; Vines et al., 2008; Wischnewski et al., 2016) and as a relay (Arai et al., 2011; Fiori et al., 2017; Koch et al., 2006). Left IPS is a primary candidate for localization of the lateralized mechanisms for precision movement control (Mutha et al., 2011), and may serve as a locus for the processes underlying left/right-hand choices (Fitzpatrick et al., 2018). Right SPL has been implicated in numerous mechanisms that support movement control, including top-down (goal-directed) visuospatial attention (Shomstein et al., 2010; Wang et al., 2016), integration of multisensory information for movement (Andersen and Buneo, 2003), combining and maintaining representations of goal and movement (Averbeck et al., 2009; Wolpert et al., 1998), and motor skill learning (Della-Maggiore et al., 2004).
We found that PMd–PMd interhemispheric connectivity did not predict subsequent NDH learning, despite previous results that point toward a potential role of PMd–PMd callosal connectivity in channeling left-hemisphere information to the right parietal cortex for the control of NDH drawing (Potgieser et al., 2015), as part of PMd's role in target-directed visuomotor control of movement and associated connectivity from ipsilateral parietal cortex (Binkofski et al., 1999; Wise et al., 1997). Our data suggested that left PMd may lie “upstream” of left M1 and IPS at rest, but our hypothesized model does not encompass the (likely many) sources of FC upstream of left M1 and IPS. Regardless, it is unsurprising that our PMd findings differed from Potgieser et al.'s (2015), because the two studies differed in design and goal: at rest, callosal PMd–PMd connections do not predict NDH learning, but during task execution, callosal PMd–PMd connections could still play a role in NDH drawing performance.
One complicating feature of our current data is that our two interhemispheric connections had opposite directions. For FC from left M1 to right SPL, more connectivity predicted greater NDH learning; but from left IPS to right SPL, less connectivity predicted greater NDH learning. This seems to arise because the two areas are involved in different networks (Fig. 5), not because of inhibitory interconnections between left M1 and left IPS. Therefore, it seems likely that the left IPS network supports or is correlated with mechanisms that support the normal dominance of the DH for precision skill (and thus impairs or is anticorrelated with the ability to work against the normal pattern of dominance). We see at least two nonexclusive ways that this could happen: motor and/or attentional. Under the motor explanation, given the role of left IPS in lateralized specializations (Mutha et al., 2011), connectivity from left IPS could reflect DH-specific mechanisms' strength and/or specificity. Under the attentional explanation, interhemispheric connectivity from left IPS could reflect the attentional mechanism that supports the normal bias of attention toward the workspace and/or affordances accessible to the DH (Colman et al., 2017; de Bruin et al., 2014). Further explanations are certainly possible, and additional research will be needed to characterize the countervailing effects of connectivity from left M1 versus left IPS.
The FC underlying our model did not overlap with the network of functional connections that change across NDH skill learning (Philip and Frey, 2016). This suggests that our model may represent a network that remains stable across time and experience. If this network is stable at the within-individual level, its presence or strength could provide a diagnostic predictor for the success of therapies that focus on NDH compensation.
Interhemispheric FC predicted NDH learning, not NDH performance
We found no interhemispheric mechanisms associated with NDH performance (baseline or post-training), only with NDH learning. This finding could arise from our choice to use task-free (resting-state) FC MRI to identify interhemispheric mechanisms. Potentially, NDH performance could involve mechanisms that are only active during the execution of NDH action. Therefore, we anticipate that future studies using task MRI will reveal additional mechanisms supporting NDH performance.
Limitations
This was a post hoc exploratory study, using a pre-existing dataset (Philip and Frey, 2016). Additional behavioral outcomes were tested (maximum performance, baseline performance, and long-term retention), with negative results. Our findings for learning of NDH performance (convergent interhemispheric connectivity on right SPL), while striking, should be confirmed in an a priori hypothesis-driven study.
Conclusions
In healthy right-handed adults, we found that the ability to learn a PDT with the NDH was predicted by interhemispheric connectivity from the left primary motor cortex and left IPS onto a small cluster in the right SPL. To our knowledge, this provides the first anatomical hypothesis of the interhemispheric mechanisms that support the ability to learn a precision skill (i.e., a traditionally DH skill) with the NDH.
If future studies can confirm this mechanism, it provides anatomical targets for noninvasive brain stimulation or brain–machine interfaces to facilitate these mechanisms, and thereby facilitate the acquisition of NDH skill for individuals with chronic impairment or loss of the dominant right hand.
Footnotes
Authors' Contributions
B.A.P. contributed to conceptualization, methodology, software, investigation, writing—original draft, writing—review & editing, visualization, funding acquisition. M.M. contributed to methodology, software, formal analysis, writing—review & editing, visualization. S.H.F. contributed to conceptualization, resources, writing—reviewing & editing, funding acquisition, supervision.
Acknowledgments
The authors thank Kelli Buchanan, Drew Hensel, and Chris Saville for their assistance with data collection.
Author Disclosure Statement
B.A.P. reports a licensing agreement with PlatformSTL to commercialize the precision drawing task, outside the submitted work.
Funding Information
This work was supported by the National Institute for Neurological Disorders and Stroke at the National Institutes of Health (grant number NS083377) to S.H.F., and by the Program in Occupational Therapy at Washington University in St. Louis to B.A.P.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.