
Abstract
Introduction:
The cingulum bundle and uncinate fasciculus are major limbic white matter tracts involved in emotion, memory, and cognition. The main goal of the present study was to investigate the relationship between age and structural properties of the uncinate fasciculus and the cingulum bundle using diffusion tensor imaging (DTI) tractography in a large cohort of healthy individuals. The second goal was to determine the effects of the catechol-O-methyl transferase (COMT) gene polymorphism on the DTI measurements of these white matter tracts.
Methods:
We recruited 140 healthy participants (18–85 years old). DTI data sets were acquired on a 1.5T magnetic resonance imaging system. The rostral, dorsal, and parahippocampal cingulum, as well as uncinate fasciculus, were delineated using deterministic tractography. Fractional anisotropy (FA), mean (MD), radial (RD), and axial (AD) diffusivities, tract volume, linear (Cl ), planar (Cp ), and spherical (Cs ) tensor shapes were calculated. The COMT polymorphism (methionine homozygous vs. valine carriers) was determined using single nucleotide polymorphism.
Results:
We found that age was negatively associated with FA, but positively associated with MD and RD for the rostral cingulum, dorsal cingulum, and the uncinate fasciculus but not for the parahippocampal cingulum. Furthermore, individuals with the COMT methionine homozygous had higher FA and lower MD, RD, AD, and Cs values in the right rostral cingulum compared with the valine carriers across the entire adult life span.
Discussion:
This study indicates that limbic tracts might be nonuniformly affected by healthy aging, and the methionine homozygous genotype might be associated with micro/macro white matter properties of the right rostral cingulum.
Impact statement
To better understand pathological age-related changes of brain's tracts, understanding normal age-related changes and its contributing factors including genetic polymorphisms are fundamental. To do so, we studied the associations between age and catechol-O-methyl transferase (COMT) polymorphism with structural properties of uncinate fasciculus and cingulum segments in a large cohort of healthy adults using deterministic tractography and fractional polynomial regression. Our results suggested that aging nonuniformly affects limbic tracts. Furthermore, in accordance with the major role of COMT in prefrontal cortex function, our findings indicated that COMT polymorphism was associated with white matter properties of the right rostral cingulum.
Introduction
The universal percentage of people at 65 years and older will reach 16.7% by 2050 (He et al., 2016). Therefore, it is important to broaden our understanding of how healthy aging affects the human brain to have a better understanding of pathological aging (Yap et al., 2013). Previous diffusion tensor imaging (DTI) studies of brain white matter in healthy aging demonstrated a decrease in fractional anisotropy (FA) with an increase in mean (MD), radial (RD), and axial (AD) diffusivities from mid-adulthood to old age (Yap et al., 2013).
The limbic system is a group of cortical and subcortical structures interconnected by white matter tracts involved in memory, spatial orientation, emotion, and motivation (Catani et al., 2013; Morgane et al., 2005; Rolls, 2015). The cingulum bundle and uncinate fasciculus are two major limbic white matter tracts (Catani et al., 2013).
The cingulum is located just above the corpus callosum and consists of short and long association fibers as well as striatal, commissural, and subcortical fibers linking the cingulate gyrus with different brain areas (Catani and Thiebaut de Schotten, 2008; Schmahmann and Pandya, 2006). Based on the connectivity profile, structural and functional properties, the cingulate cortex can be divided into four subregions including the anterior cingulate, midcingulate, posterior cingulate, and retrosplenial cortices (Vogt and Palomero-Gallagher, 2012). The cingulum is therefore considered a heterogeneous tract since it carries afferent and efferent projections from structurally and functionally distinct cingulate subregions; however, most studies of healthy aging have analyzed the cingulum either as a single structure that corresponds to (1) the dorsal cingulum (Lebel et al., 2012), (2) rostral+dorsal cingulum (Sala et al., 2012; Stadlbauer et al., 2008), (3) dorsal+parahippocampal cingulum (Voineskos et al., 2012) or as (4) two separate structures that correspond to the rostral+dorsal cingulum and parahippocampal cingulum (Bennett et al., 2015; Cox et al., 2016; Westlye et al., 2010).
Only two studies (Jang et al., 2016; Michielse et al., 2010) have reported age-related differences in DTI measurements for more than two cingulum subdivisions. Age-related decline in FA has been reported in the anterior cingulum, anterosuperior cingulum (Jang et al., 2016), dorsal cingulum (Lebel et al., 2012), rostral+dorsal cingulum (Bennett et al., 2015; Cox et al., 2016; Sala et al., 2012; Westlye et al., 2010), but not in the parahippocampal cingulum (Bennett et al., 2015; Cox et al., 2016; Jang et al., 2016; Michielse et al., 2010). In contrast, other studies did not report any associations between age and cingulum's FA (Michielse et al., 2010; Stadlbauer et al., 2008).
The uncinate fasciculus arises from the temporal pole, uncus, parahippocampal gyrus, and amygdala (Catani et al., 2013). It connects the anterior portion of the temporal lobe to the inferior frontal gyrus and the orbitofrontal cortex (Nieuwenhuys et al., 2008). It has been suggested that the uncinate fasciculus might be involved in emotion, memory, and language functions (Catani and Thiebaut de Schotten, 2008). Previous DTI studies of healthy aging reported either curvilinear (Michielse et al., 2010) or negative linear (Bennett et al., 2015; Cox et al., 2016; Voineskos et al., 2012) trajectories for the FA of the uncinate fasciculus with age; this inconsistency might be due to differences in sample characteristics, DTI acquisition protocols, DTI data analyses, and statistical approaches.
Several previous studies suggested that different genetic polymorphisms could influence both gray and white matter changes in limbic structures (Kanchibhotla et al., 2013; Petrella et al., 2008). One of the genes related to the prefrontal cortex's structure and functions is the catechol-O-methyl transferase (COMT) gene involved in dopamine neurotransmission and degradation (Deary et al., 2004; Mier et al., 2009; Petrella et al., 2008). Dopamine has been found to exert protective effects on oligodendrocytes (Rosin et al., 2005) and it may be involved in oligodendrocytes differentiation (Bongarzone et al., 1998). The Val158Met polymorphism (rs4680) is a common COMT polymorphism in which the amino acid valine (Val) is substituted with methionine (Met) (Goldman et al., 2005). The Val allele has higher dopamine degrading activity than the Met allele, resulting in lower synaptic dopamine levels. Since the presynaptic dopamine transporter in the prefrontal cortex is low, COMT is a major regulator of dopamine levels in the prefrontal cortex (Sambataro et al., 2012). Previous studies of the Val158Met polymorphism mainly focused on gray matter structures and found differences between alleles on the volumes of the hippocampus, temporal and frontal lobes, anterior cingulate and dorsolateral prefrontal cortices, as well as on cortical thickness of the right inferior prefrontal and the right superior temporal sulci (for review, see Sambataro et al., 2012; Witte and Flöel, 2012). A single DTI study that employed tract-based spatial statistics (TBSS) approach investigated the effects of COMT polymorphism on brain white matter (Papenberg et al., 2015), and the authors found higher FA and lower MD in the cingulum of the Met homozygotes compared with Val carriers only in the oldest adult group (81–87 years old). However, associations between COMT polymorphism and DTI measures from limbic white matter tracts have not been investigated using deterministic DTI tractography.
Therefore, the main goal of the present study was to investigate the effects of age on white mater structure of three cingulum subdivisions and the uncinate fasciculus in a large sample of healthy individuals across the entire adult life span. Considering findings from previous studies on the rostral (Jang et al., 2016; Michielse et al., 2010), dorsal (Bennett et al., 2015; Cox et al., 2016; Lebel et al., 2012; Michielse et al., 2010; Sala et al., 2012; Westlye et al., 2010), and parahippocampal (Cox et al., 2016; Jang et al., 2016; Michielse et al., 2010) cingulum and the uncinate fasciculus (Bennett et al., 2015; Cox et al., 2016; Hasan et al., 2009; Lebel et al., 2012; Michielse et al., 2010; Sala et al., 2012; Voineskos et al., 2012; Westlye et al., 2010) in healthy aging, which showed age-related decrease in FA and increase in MD in the uncinate fasciculus, rostral cingulum, and dorsal cingulum, but not in the parahippocampal cingulum, we hypothesized that the parahippocampal cingulum would show relative preservation with age, compared with the uncinate fasciculus, rostral cingulum, and dorsal cingulum. Our second goal was to determine whether COMT polymorphism is associated with the DTI characteristics of these white matter tracts. Due to the limited knowledge regarding the effects of COMT polymorphism on white matter tracts, we made no prior hypothesis regarding this investigation.
Materials and Methods
Participants
One hundred forty healthy volunteers (62 men, 78 women), aged between 18 and 85 years (mean: 48.27, standard deviation: 18.47), were recruited through online, poster, and local advertisements. The recruitment procedure, exclusion criteria, and inclusion criteria have been previously reported in detail elsewhere (Aghamohammadi-Sereshki et al., 2019). For more details, see the Supplementary Material. The study was approved by the University of Alberta Health Research Ethics Board (Pro00004229). Written informed consent was obtained from each participant.
Magnetic resonance imaging acquisition and data analysis
Participants were scanned on a 1.5T Siemens Sonata MRI system. A cradle and bilateral head support were used to minimize subject motion in the magnetic resonance imaging (MRI) scanner. A whole-brain T1-weighted 3D MPRAGE sequence (TR: 2080 ms; TE: 4.38 ms; inversion time: 1100 ms; flip angle: 15°; bandwidth: 130 Hz/Px; FOV: 256 × 192 × 144 mm3; voxel size: 1 × 1 × 1 mm3; scan time: 5 min 49 sec) was acquired for intracranial volume (ICV) estimation (Eritaia et al., 2000). DISPLAY (Montreal Neurological Institute, QC, Canada) software was used to trace the ICV. DTI data sets were acquired using a twice-refocused spin echo, echo planar imaging sequence (Reese et al., 2003) with the following parameters: TR = 10,000 ms, TE = 94 ms, 5 nondiffusion weighted (b = 0 sec/mm2) and 30 diffusion directions (b = 1000 sec/mm2), FOV: 212 × 212 × 143 mm3, voxel size: 2.2 × 2.2 × 2.2 mm3, with full brain coverage without gap and a scan time of 4 min 39 sec. The diffusion tensor data sets were preprocessed by the rigid body co-registering to b0 image using Automatic Image Registration (AIR) to correct for subjects' motion and eddy current distortion. All imaging data sets were reconstructed and visually inspected for motion artifacts while the subject was in the scanner, allowing a second or a third data set to be collected if required.
Detailed reliable protocols for the deterministic DTI tractography of the uncinate fasciculus and the rostral, dorsal, and parahippocampal cingulum were previously reported (Malykhin et al., 2008; Michielse et al., 2010) (Fig. 1). The program DTI-studio V2.40 (Johns Hopkins University, Baltimore, MD) was utilized to quantify FA, MD, AD, RD, and tract volume. In addition, linear [Cl = (λ1 − λ2)/λ1], planar [Cp = (λ2 − λ3)/λ1], and spherical (Cs = λ3/λ1) measures were calculated as representations for geometrical shapes of diffusion tensor (Westin et al., 2002).

Three-dimensional reconstructions of the cingulum segments and uncinate fasciculus from a healthy participant. Uncinate fasciculus is shown in pink, rostral cingulum is shown in cyan, dorsal cingulum is shown in purple, and parahippocampal cingulum is shown in lime green. Color images are available online.
All measurements were performed by a single rater (A.A.S.) who was blind to all demographic and genotype information. Raw tract volumetric measurements were adjusted to the ICV by using the following formula: ICV-adjusted volume = [Raw tract volume (mm3)/ICV of the same individual (cm3)] × sample average ICV (cm3) (Aghamohammadi-Sereshki et al., 2019). Fiber tracking parameters were >0.2 for FA threshold to initiate and continue tracking, and a termination threshold of FA <0.2 or a >60° angular deviation from the propagating line.
The inter/intra-rater reliabilities were assessed using intraclass-correlation coefficient [as we previously described in the studies of Michielse et al. (2010) and Malykhin et al. (2008); see the Supplementary Material and Supplementary Table S1].
Genetic analysis
Genotype was obtained using single nucleotide polymorphism (SNP) derived from cheek swabs from our participants. Primer3PLUS was used to generate primers using sequence data from dbSNP. Primers were positioned to generate a clean single PCR product with the variant in a position near the center of the read to avoid common technical artifacts that can occur during the sequencing step. All primers were purchased from Integrated DNA Technologies (Coralville, IA). COMT polymorphism was obtained using rs4680 (G; A), where the G allele encodes Val and the A allele encodes Met. Due to the possible advantage of the Met allele over the Val allele in cognitive-related functions (Dixon et al., 2014; Mier et al., 2009; Petrella et al., 2008), individuals were grouped according to COMT polymorphism as homozygous “Met/Met” vs. “Val/x” (i.e., Val/Val and Met/Val).
Statistics
Demographic information
All inferential statistics other than regression models for age and COMT effects were carried out using IBM SPSS Statistics 26. Independent sample t-test was used to compare age, education, and ICV between men and women. Moreover, demographic characteristics between COMT (Met/Met vs. Val/x) polymorphism groups were also analyzed using a t-test. In addition, the chi-squared test was used to evaluate deviation from the Hardy–Weinberg equilibrium.
Hemisphere-related effects analysis
To investigate potential Age × Hemisphere interactions in DTI measures, we tested for significant Pearson's correlations between age and asymmetry indices for each DTI measurement using the following formula: [({(Value of the right hemisphere)/(Value of the left hemisphere)} × 100) − 100]. This test demonstrated whether the differences between hemispheres were consistent across the adult life span. Also, to study hemispheric differences in DTI characteristics, we used paired sampled t-test or repeated-measures analysis of covariance (ANCOVA; using age as a covariate), as appropriate.
Modeling the relationships between age, white matter structural properties, COMT polymorphism, and sex
Lower order regression models (i.e., linear, quadratic, and cubic) tend to impose severe shape restrictions to trend lines in regression analyses. Fjell and colleagues (2010) illustrated that studies utilizing quadratic models to estimate parabolic appearing age trajectories of brain structures are significantly influenced by the age range of the subjects. Therefore, Fjell and colleagues (2010) recommended the use of nonparametric smoothing splines. However, since a smoothing spline is a local nonparametric regression model, it is not well suited for inferential statistics. Also, using nonparametric smoothing splines on smaller sample sizes may produce higher order functions, leading to overfitting and poor generalizability (Fjell et al., 2010; Royston and Sauerbrei, 2008a). Considering the aforementioned limitations, we used multivariable fractional polynomial (MFP) regression modeling to assess the relationship between DTI measurements with age and COMT (Royston and Sauerbrei, 2008b).
To analyze the associations between age, DTI measurements, and COMT polymorphism, DTI measurements were selected as dependent variables and “Age” (centered), dummy-coded variables of the “Sex” and COMT (Met/Met and Val/x) as well as “Age × Sex” and “COMT × Age” interaction terms were selected as independent variables in MFP. In addition, to account for the partial volume effects on DTI measurements (Alexander et al., 2001; Vos et al., 2011), the raw tract volume was included as an independent variable not-of-interest (Vos et al., 2011) in MFP regression model or ANCOVA-related analyses. MFP regression models were analyzed by STATA 16.1. For more details about MFP, see the Supplementary Material.
If the “Sex” and “COMT” terms were selected as influential independent variables in the regression models, we conducted ANCOVA, using raw tract volume and/or age as covariate(s) to indicate if the aforementioned impacts were statistically different between either sexes or genotypes.
Finally, the Holm–Bonferroni correction for type I error inflation due to multiple comparisons was used. Since several previous studies, including our own (Hasan et al., 2009; Lebel et al., 2012; Michielse et al., 2010; Sala et al., 2012; Yap et al., 2013), demonstrated the effects of age on the FA, MD, RD, AD, and tracts' volumes of the cingulum bundle and uncinate fasciculus, the Holm–Bonferroni correction was only applied to the analyses of tensor shape-related measurements. Furthermore, the Holm–Bonferroni correction was also used in hemispheric, sex, and genetic analyses. p Values that remained significant after the Holm–Bonferroni correction are shown with an asterisk in the tables and the Results section.
Results
Demographics
Participants' characteristics for the entire sample are shown in Table 1. Males had larger ICVs (p < 0.0001*) than females while males and females did not differ in age and education (both ps > 0.09, Table 1). Demographic characteristics did not differ between COMT polymorphism groups (“Met/Met” vs. “Val/x,” all ps > 0.19). Moreover, the chi-squared test showed that our sample for the COMT (χ2: 0.38, p: 0.54) was in the Hardy–Weinberg equilibrium. Alleles' frequency for Met and Val was 0.464 and 0.536, respectively.
Demographic Information (Mean ± Standard Deviation)
Remained significant after the Holm–Bonferroni correction.
ICV, intracranial volume.
Hemispheric analyses
Between all white matter tracts, only asymmetry indices for the RD (p < 0.0004*) and MD (p < 0.008*) of the rostral cingulum showed significant relationships with age. Therefore, we analyzed DTI measurements of the left and right rostral cingulum separately, whereas the averaged DTI measurements of other white matter tracts were used in the regression analyses.
There were significant hemispheric differences in most of the DTI measurements in all studied limbic white matter tracts. Significantly, higher FA and lower MD, RD, and AD values in the uncinate fasciculus, rostral cingulum, and dorsal cingulum were found in the left hemisphere compared with the right hemisphere (Table 2). In contrast, the right parahippocampal cingulum had significantly higher FA and lower MD, RD, and AD compared with the left counterpart (Table 2).
Hemispheric Effects on Diffusion Tensor Imaging Measurements
ANCOVA represents repeated-measures ANCOVA, t-test represents paired sample t-test. Significant p values that survived the Holm–Bonferroni correction are shown in asterisk.
AD, axial diffusivity; ANCOVA, analysis of covariance; Cl , linear geometry; Cp , planar geometry; Cs , spherical geometry; DC, dorsal cingulum; FA, fractional anisotropy; MD, mean diffusivity; PHC, parahippocampal cingulum; RC, rostral cingulum; RD, radial diffusivity; SD, standard deviation; UF, uncinate fasciculus.
Relationships between age, sex, and COMT polymorphism with DTI measurements
Associations between age and DTI measurements
We found a significant linear negative relationship between age and FA values for the rostral cingulum in both hemispheres, dorsal cingulum, and the uncinate fasciculus (all ps < 0.0004), but not for the parahippocampal cingulum (Fig. 2).
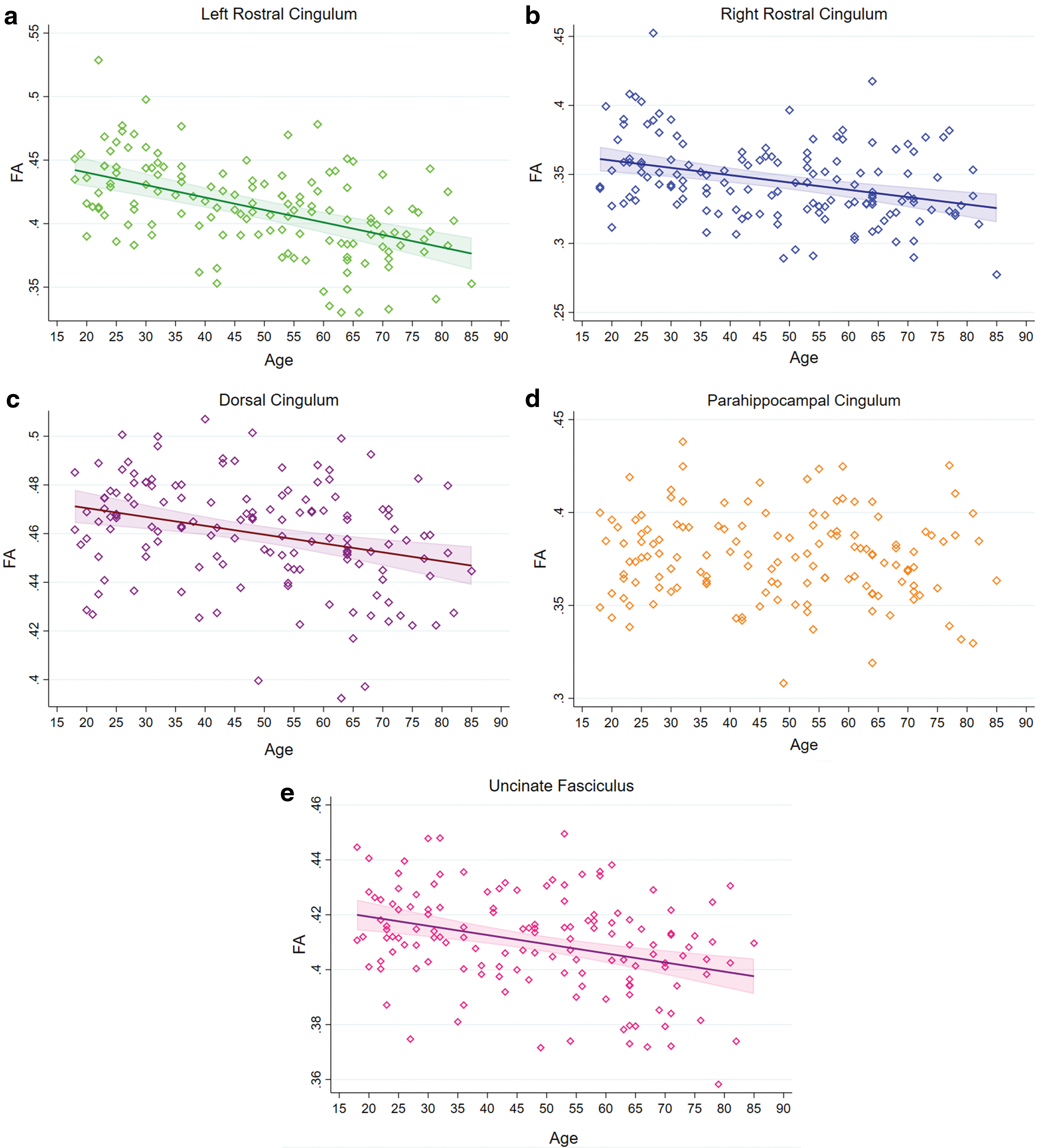
Regression plots showing the relationship between age and FA in the cingulum segments
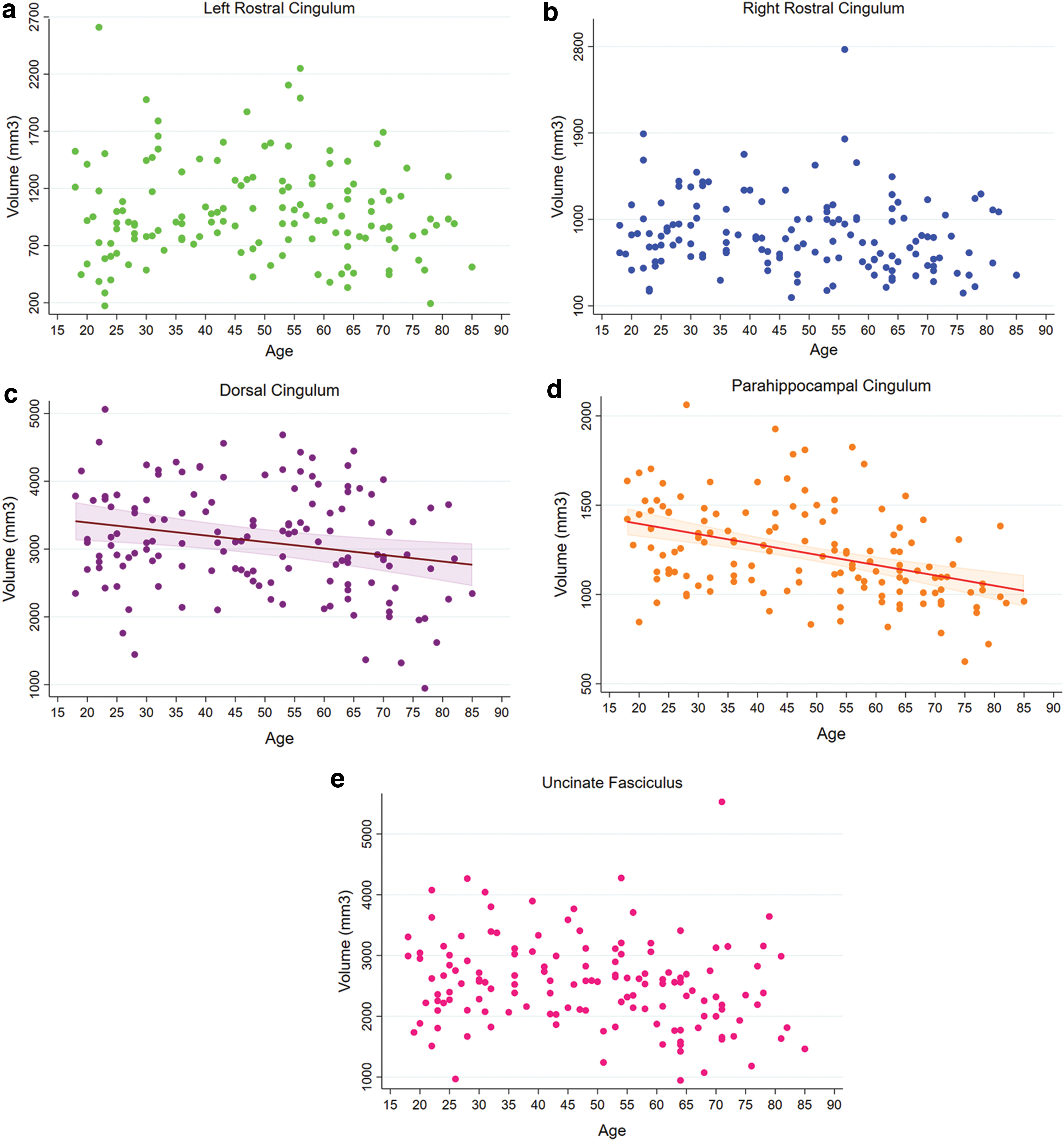
Significant associations between MD (all ps < 0.0036) and RD (all ps < 0.0001) with age were observed for the left rostral cingulum, dorsal cingulum, and the uncinate fasciculus, but not for the right rostral cingulum and parahippocampal cingulum. Furthermore, while the associations between age and MD or RD of the dorsal cingulum and uncinate fasciculus were nonlinear, for the left rostral cingulum, these were positive and linear (Fig. 3). Significant associations between age and AD were found for the dorsal cingulum and the uncinate fasciculus (both ps < 0.0021), but not for the rostral and parahippocampal cingulum. These significant associations were nonlinear toward higher AD in elderly individuals (Fig. 3).

Regression plots showing the relationship between age and MD, RD, and AD for the cingulum segments and the uncinate fasciculus. Values for MD


We found significant negative linear associations between age and normalized tract volume for the dorsal cingulum and the parahippocampal cingulum (both ps < 0.0069), but not for the uncinate fasciculus and rostral cingulum (Fig. 4).
Regression plots showing the relationship between age with normalized tract, linear tensor shape, and the spherical tensor shape for the cingulum segments and the uncinate fasciculus. Normalized tract volumes in mm3
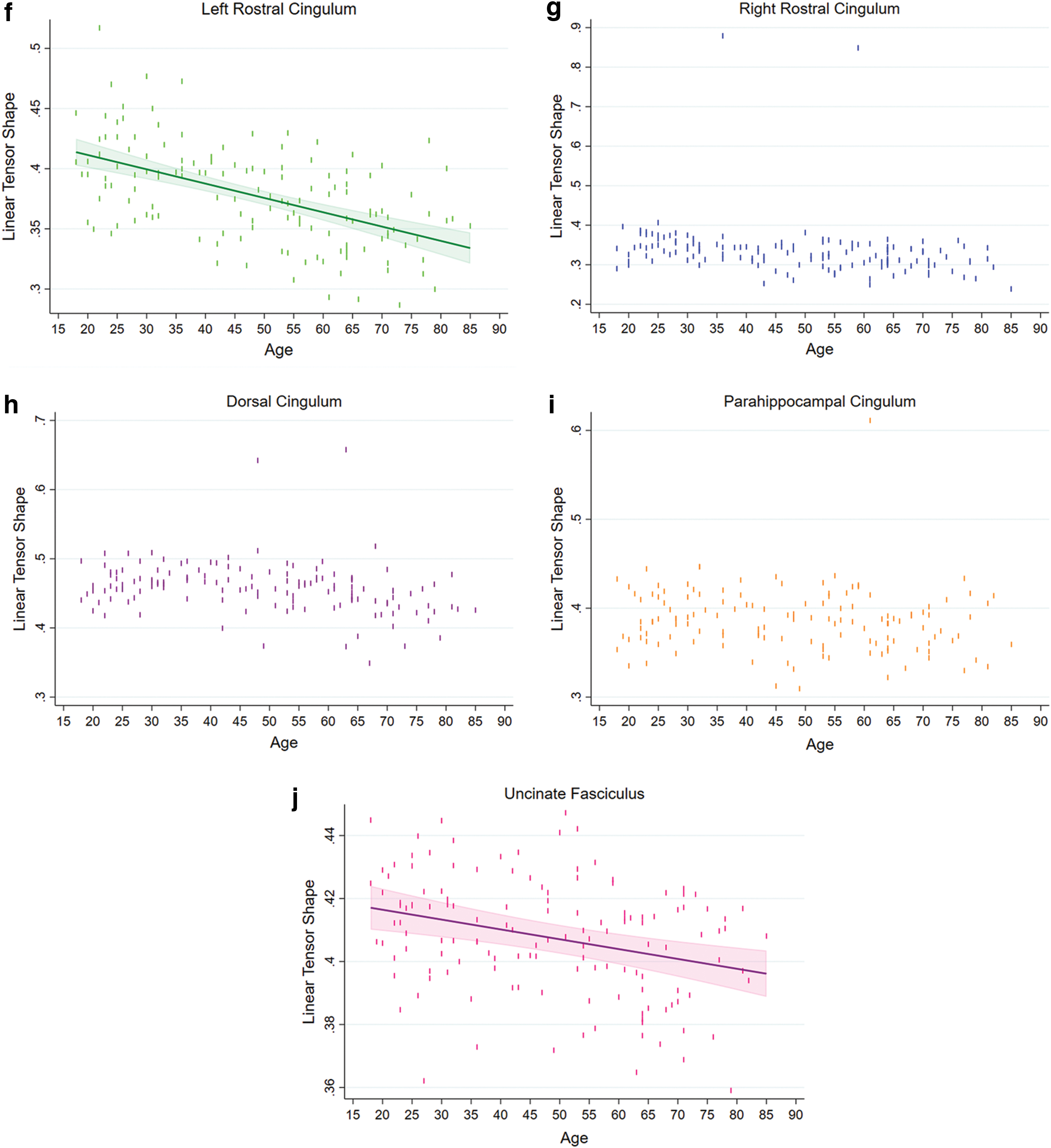
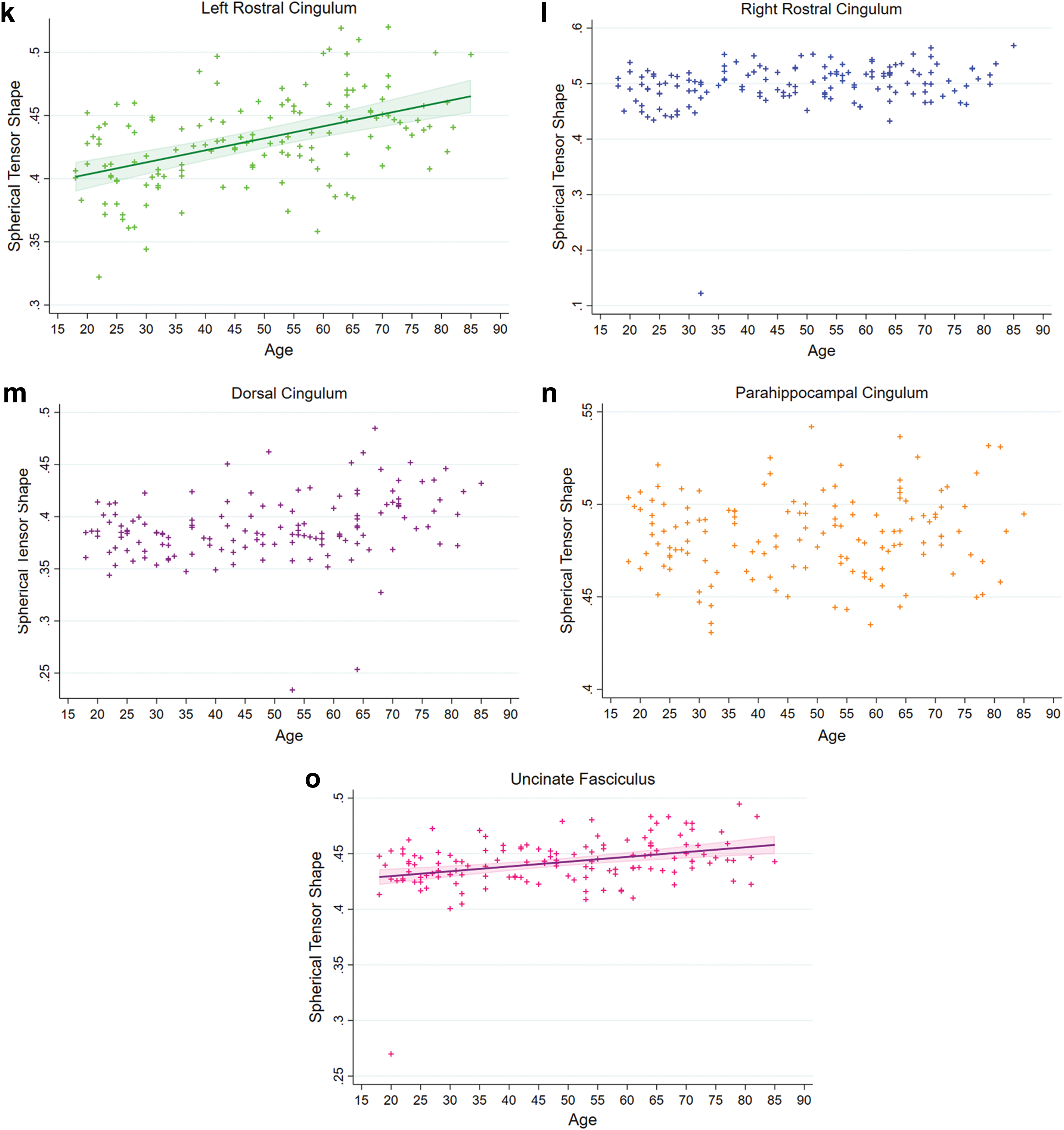
The Cl values of the left rostral cingulum and the uncinate fasciculus had significant negative linear associations with age (both ps < 0.0004), whereas there was no significant association between age and Cl value for the other tracts (Fig. 4). We found significant positive linear associations between Cs values and age for the left rostral cingulum and the uncinate fasciculus (both ps < 0.0001), but not for the right rostral, dorsal, and parahippocampal cingulum (Fig. 4). We did not find any significant associations between Cp values and age in the studied tracts.
Associations between sex and DTI measurements
The regression analysis showed that sex term was significantly associated with the FA of the left rostral cingulum, volume of the rostral cingulum in both hemispheres, volume of the dorsal cingulum, Cl of the dorsal cingulum and uncinate fasciculus, and Cp and Cs of the left rostral cingulum (Table 3). No significant “Age × Sex” interactions for any of the DTI measurements of the cingulum sections and uncinate fasciculus were found (Table 3). ANCOVA and independent sample t-test showed that except for the volume of the left rostral cingulum, which was statistically larger in males compared with females (p < 0.0017*), none of the above-mentioned DTI measurements statistically differed between sexes (all corrected ps > 0.10, Supplementary Table S2).
Regression Models of the Age-, Sex-, and Catechol-O-Methyl Transferase-Related Effects on the Diffusion Tensor Imaging Measurements of the Cingulum Bundle and Uncinate Fasciculus with the Corresponding Regression Results [R 2, Adj-R 2, and Standardized β (p Value)]
Remained significant after the Holm–Bonferroni correction.
Negative standardized βs represent higher values in men or Val carriers, whereas positive standardized βs represent higher values in women or Met/Met individuals.
Due to the previous studies, FA, MD, RD, AD, and volume were not corrected for multiple comparisons, but the Holm–Bonferroni correction was applied to Cl , Cp , and Cs .
The statistical significance of these associations was affected as a result of inclusion of the raw tract volume in the regression models. However, the findings of the statistical significance for the rest of the associations were not altered by incorporation of the raw tract volumes in regression models.
COMT, catechol-O-methyl transferase; DTI, diffusion tensor imaging; FP1, fractional polynomial regression degree 1; FP2, fractional polynomial regression degree 2; Met, methionine; NA, not applicable; NS, not significant; Val, valine.
Associations between COMT polymorphism and DTI measurements
Regression analysis showed that COMT genotype was significantly associated with the FA, MD, RD, AD, and Cs of the right rostral cingulum (Table 3). Met/Met carriers had higher FA, and lower MD, RD, AD, and Cs values compared with Val carriers (Table 3). We did not find significant effects of the COMT polymorphism on any other DTI measurements in other white matter tracts. Finally, we did not find any significant “Age × COMT” interactions for any of the DTI measurements of the cingulum sections and uncinate fasciculus (Table 3).
ANCOVA and independent sample t-test demonstrated that while the FA (p < 0.022*) was significantly higher in Met/Met individuals, the MD (p < 0.00008*), RD (p < 0.0002*), and AD (p < 0.009*) were significantly higher in Val carriers, compared with Met/Met individuals (Table 4). In addition, there was a trend toward higher Cs values in Val carriers compared with Met/Met individuals (p < 0.064). All regression-related results of DTI measurements are presented in Table 3.
Comparing the Diffusion Tensor Imaging Measurements of the Right Rostral Cingulum Between Catechol-O-Methyl Transferase Genotypes
Remained significant after the Holm–Bonferroni correction.
Val/x represents Val/Val and Met/Val carriers.
p Value is adjusted for the violation of the homogeneity of variance (Levene's test).
There was no significant difference in raw volume of the right rostral cingulum between Met/Met and Val/x controlled for age as covariate (p > 0.65). Therefore, raw volume of the right rostral cingulum was not included as a covariate in this analysis.
Discussion
In this study, we used DTI tractography to examine the associations between age, sex, and COMT polymorphism with the DTI measurements of the cingulum subdivisions and uncinate fasciculus. Our results showed that limbic tracts were differentially affected by healthy aging. First, we found significant relationships between age and most of DTI measurements in the uncinate fasciculus, dorsal cingulum, and left rostral cingulum, whereas the parahippocampal cingulum and right rostral cingulum showed relative preservation with age. Second, although we found larger volume of the left rostral cingulum in males compared with females, our results did not reveal any other significant sex or Age × Sex interaction effects on any studied limbic tracts. Third, despite the fact that we found significant hemispheric differences in DTI measurements in all studied white matter tracts, significant age by hemisphere interactions were found only in the rostral cingulum, suggesting potential differences in aging trajectories between hemispheres for this structure. Finally, we found that COMT polymorphism was associated with DTI measurements of the right rostral cingulum independent of age.
Patterns of aging for the limbic white matter tracts
Previous studies demonstrated that the older age was associated with lower FA in the rostral (Jang et al., 2016), dorsal (Lebel et al., 2012), or rostral+dorsal cingulum subdivisions (Bennett et al., 2015; Cox et al., 2016; Sala et al., 2012; Westlye et al., 2010), but not in the parahippocampal cingulum (Bennett et al., 2015; Cox et al., 2016; Jang et al., 2016; Michielse et al., 2010). Although Westlye and colleagues (2010) found an inverse U-shaped relationship between age and the FA in the parahippocampal cingulum, they acknowledged that the parahippocampal cingulum had a high degree of interindividual variability, which caused to have a “less clear-cut trajectory.” In contrast, other studies did not find any associations between age and FA in the rostral+dorsal cingulum (Stadlbauer et al., 2008) or in all the three cingulum segments (Michielse et al., 2010). Our findings on negative association between age and FA values in the rostral and dorsal cingulum, but not in the parahippocampal cingulum, are in accordance with most of the mentioned studies.
Our findings on age-related reduction in FA values in the uncinate fasciculus also agree with previous studies that consistently reported decline in FA with age (Bennett et al., 2015; Cox et al., 2016; Hasan et al., 2009; Lebel et al., 2012; Michielse et al., 2010; Sala et al., 2012; Westlye et al., 2010).
Similarly, the majority of the previous studies observed increases in MD and/or RD values with age in the dorsal, rostral+dorsal cingulum segments, as well as in the uncinate fasciculus (Bennett et al., 2015; Cox et al., 2016; Hasan et al., 2009; Lebel et al., Michielse et al., 2010; Sala et al., 2012; Westlye et al., 2010), which agree with our findings. However, Stadlbauer and colleagues (2008) did not find any associations between age and the MD of rostral+dorsal cingulum. Our findings on the absence of the significant correlations between age and MD and RD in the parahippocampal cingulum agree with some previous studies (Bennett et al., 2015; Michielse et al., 2010), but contradict the studies of Cox and colleagues (2016) and Westlye and colleagues (2010) who reported quadratic associations with higher values in older age.
Compared with the MD and RD results, findings on the effects of healthy aging on AD are less clear. While some previous studies did not find any associations between age and AD in the rostral+dorsal cingulum (Bennett et al., 2015; Cox et al., 2016; Stadlbauer et al., 2008) and uncinate fasciculus (Bennett et al., 2015; Hasan et al., 2009), others reported both decreasing (Sala et al., 2012) and increasing AD (Cox et al., 2016; Michielse et al., 2010) in the uncinate fasciculus and cingulum segments with increasing age. Our results suggest that age was associated with nonlinear changes in AD in both the dorsal cingulum and uncinate fasciculus. Finally, although Cox and colleagues (2016) found a U-shape correlation between age and AD in the parahippocampal cingulum in healthy adults aged between 44 and 77 years, we did not find any significant relationship between age and AD in the parahippocampal cingulum.
There are several factors that might have contributed to the observed differences between studies, including DTI acquisition protocols, FA threshold for tractography, methods of DTI analyses (e.g., ROI-based vs. TBSS; deterministic vs. probabilistic), anatomical definition of tracts, and age range of participants. More importantly, most above-mentioned studies tested only linear and quadratic models to explain the relationships between age and DTI measurements, which might not adequately explain the nature of the relationships. As Fjell and colleagues (2010) noted, in age-related studies, the quadratic trajectory shape is significantly affected by the age range of the sample. Therefore, while the quadratic regression might properly fit data in studies that included children, adolescents, younger, and older adults, it could significantly affect the maximum or minimum values as well as the aging trajectories in studies that only recruit older adults. The quadratic model is only one type of nonlinear model. Therefore, other nonlinear regression models should be statistically tested to determine whether more complicated models are statistically better for describing these relationships. In the current study, we used the MFP method to test if a nonlinear model was statistically better than a linear model. Furthermore, more complicated nonlinear models (FP2) were tested against the simpler nonlinear models (FP1) to determine if more complicated nonlinear models were statistically better than the simple nonlinear models. Our analyses showed that among all 40 tests (5 tracts × 8 DTI measurements), the age term was significant for 18 tests. Of these, 12 models were linear and 6 were nonlinear models. None of the age-related regression models was quadratic.
Considering the noted differences between studies, it is difficult to directly compare the aging trajectories for DTI measures across studies. However, the studies that included only adults reported linear negative association between age and FA (Bennett et al., 2015; Cox et al., 2016; Jang et al., 2016; Voineskos et al., 2012), whereas MD, RD, and AD showed nonlinear relationships with age (Cox et al., 2016; Michielse et al., 2010).
Finally, it was demonstrated that the direction of DTI major eigenvector (ν1) may not have the same alignment as the underpinning anatomical structures, which make the biological-related interpretations of RD and AD difficult (Wheeler-Kingshott and Cercignani, 2009). Therefore, it was recommended providing information about geometrical features besides RD and AD (Wheeler-Kingshott and Cercignani, 2009). The geometric nature of diffusion tensors (i.e., Cl , Cp , and Cs ) provides information about the macrostructural diffusion, which can be exploited to examine fiber tract organization (Westin et al., 2002). In our study, the Cp values were not associated with age in any of the studied tracts and Cl showed significant negative linear relationships with age in the left rostral cingulum and the uncinate fasciculus. However, for the left rostral cingulum and the uncinate fasciculus, Cs had positive linear associations with age. This might represent less restricted diffusion with aging. Finally, in addition to microstructural diffusion parameters of the parahippocampal cingulum that were not associated with age, tensor shapes of the parahippocampal cingulum did not correlate with age either, suggesting that its structural geometry and fiber tract organization are not affected by healthy aging.
Effects of age on interhemispheric and sex-related differences in DTI measures
The present study demonstrated significantly higher FA, lower MD, RD, and AD values in the left hemisphere compared with the right hemisphere throughout the adulthood in the rostral and dorsal cingulum segments and in the uncinate fasciculus, which agree with our previous study (Malykhin et al., 2008). Furthermore, our results indicated significantly larger volume of the uncinate fasciculus in the right hemisphere compared with the left hemisphere. This is in agreement with both our previous results (Malykhin et al., 2008) and a postmortem study that showed the uncinate fasciculus was 27% larger on the right hemisphere compared with the left hemisphere (Highley et al., 2002). For the parahippocampal cingulum, we found an opposite pattern, significantly higher FA and lower MD, AD, and RD in the right hemisphere compared with the left hemisphere throughout adulthood. However, no hemispheric differences in FA for the uncinate fasciculus, and dorsal+parahippocampal cingulum (Voineskos et al., 2012) or in FA, MD, and AD for the rostral+dorsal cingulum (Stadlbauer et al., 2008) were reported.
In support of previous studies (Hasan et al., 2009; Lebel et al., 2012), we did not find any significant effects of sex on age-related trajectories of limbic white matter tracts. Nevertheless, Cox and colleagues (2016), who studied 27 tracts across the entire brain with a large sample size (N = 3513, age range: 44.64–77.12), reported that although there were some significant Age × Sex interactions, they were small and inconsistent. Furthermore, the authors concluded that power calculation analyses indicated with sample size of 1000 individuals, the modest Age × Sex interactions would not be detected (Cox et al., 2016).
Finally, our results indicated larger left rostral cingulum in males compared with females and an absence of sex differences in the DTI measurements in other limbic white matter tracts.
Effects of the COMT polymorphism on limbic white matter tracts
Our findings suggest that the COMT polymorphism is associated with DTI measurements of the right rostral cingulum. Homozygous Met/Met carriers had significantly higher FA and significantly lower MD, RD, and AD compared with Val/+ carriers. These differences were preserved across entire adult life span as indicated by the absence of the COMT × Age interactions.
Most of the healthy aging studies of COMT have focused on cognition (Sambataro et al., 2012; Witte and Flöel, 2012) and the effects of COMT on brain structure have been less studied. Papenberg and colleagues (2015) used the TBSS method and found higher FA and lower MD in the rostral+dorsal cingulum in Met homozygotes compared with Val carriers only in the oldest adult group (81–87 years old). Although our findings on higher FA and lower MD in Met homozygotes compared with the Val carriers are in accordance with those of Papenberg and colleagues (2015), the authors found that significant associations between the COMT polymorphism and microstructural diffusion properties of the cingulum were present only in individuals between the ages of 81 and 87 years and were absent in participants between the ages of 60 and 78 years. In contrast, in the current study, the COMT-related associations with diffusivity measurements were present across the entire adult life span. Future research is necessary to clarify whether the COMT (rs4680) polymorphism affects the frontal white matter due to its effects on dopamine signaling in myelination or other mechanisms.
Our study has some limitations. This study was a cross-sectional, and therefore, all age-related findings should be interpreted as correlations/associations with age. DTI data sets were acquired on a 1.5T scanner, while more recent DTI studies employed 3T scanner that can improve signal-to-noise ratio. Although individuals' motion-related effects were minimized using head support and correction with AIR, more sophisticated motion correction methods are available, but at a greater computational cost. Generally, DTI measures in tractography studies could be affected by proximity to cerebrospinal fluid (CSF) (Concha et al., 2005), tract anatomy, and volume (Vos et al., 2011). Consequently, using different approaches such as CSF suppression (Concha et al., 2005), correction for the tract volume (Vos et al., 2011), and correcting on the voxel-wise basis using free water elimination (Metzler-Baddeley et al., 2012; Pasternak et al., 2009) have been proposed. Based on our results, we found that using tract volume as a control independent variable affected associations between age with Cs and Cl , but not age with FA, MD, RD, and AD or COMT-related associations (Table 3). Therefore, including tract volume as an independent variable of not-of-interest would improve the specificity of DTI analysis (Vos et al., 2011). Finally, although our sample covers the adult life span, a larger sample size for the genetic component of the current study would be more satisfactory.
Conclusion
In summary, we found that limbic tracts might be differentially affected by healthy aging. While diffusivity parameters of the uncinate fasciculus, rostral cingulum, and dorsal cingulum showed strong associations with advanced age, the parahippocampal cingulum was relatively preserved. In addition, our findings indicate that the COMT Met/Met genotype was associated with higher FA and lower MD, RD, AD, and Cs in the right rostral cingulum.
Footnotes
Acknowledgments
We would like to thank members of the Applied Genomics Core (TAGC, Faculty of Medicine & Dentistry, University of Alberta), Dr. Andrew L. Mason (Director), Dr. Georgina Macintyre, and Ms Susan Kenney for their help with genetic data collection. Also, we would like to express our gratitude to Dr. Richard Camicioli, and Wojciech Pietrasik for reviewing the article. Finally, the authors are thankful to all individuals who participated in this study.
Authors' Contributions
N.V.M. designed the study. F.O., P.S., and N.V.M. acquired the data. A.A.S. and N.V.M. analyzed the data. A.A.S. wrote the original draft of the article, which all authors reviewed.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Financial support for this study was provided by the Canadian Institutes of Health Research (CIHR) operating grant MOP115011 to N.V.M.
Supplementary Material
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.