
Abstract
Background:
Mapping diffusion MRI tractography streamlines to the cortical surface facilitates the integration of white matter features onto gray matter, especially for connectivity analysis.
Method:
In this work, we present methods that combine cortical surface meshes with tractography reconstruction to improve endpoint precision and coverage. This cortical mapping also enables the study of structural measures from tractography along the cortex and subcortical structures. In addition to structural connectivity analysis, novel adaptive and dynamic surface seeding methods are proposed. These improvements are made by incorporating cortical maps such as endpoint density.
Results:
The proposed dynamic surface seeding increases the cortical coverage and reduces endpoint location biases. Our results suggest that the use of cortical and subcortical meshes together with a proper seeding strategy can reduce the variability in structural connectivity analysis.
Conclusion:
The proposed adaptive and dynamic seeding utilize cortical maps to better distribute tractography interconnections, thus increasing cortical coverage and reducing endpoint bias. This also facilitates the analysis of white matter & diffusion MRI features along the cortex, combined with cortical measures or functional activation.
Impact statement
This research presents an overview of surface mapping methods for tractography to reduce structural connectivity variability. The proposed adaptive and dynamic seeding utilize cortical maps to better distribute tractography interconnections, thus increasing cortical coverage and reducing end-point bias. This also facilitates the analysis of white matter and diffusion magnetic resonance imaging features along the cortex, combined with cortical measures or functional activation.
Highlights
Novel adaptive and dynamic cortical seeding methods for tractography.
Increased cortical coverage and reduced gyral bias from tractography end-points.
Reduction of structural connectivity variability, with Human Connectome Project test–retest data set.
Introduction
Diffusion Weighted magnetic resonance imaging (MRI), along with tractography methods, can be used to explore and estimate white matter (WM) architecture. This reconstruction of WM pathways is often employed for structural connectivity studies (Fornito et al., 2013; Hagmann et al., 2007; Wakana et al., 2007). Pathways interconnecting two gray matter (GM) regions, produced by tractography, are used to estimate structural connectivity mapping (Jbabdi et al., 2015; Yo et al., 2009).
Anatomically, WM fascicles interconnect two GM regions or emerge from the brainstem, into the cerebellum or spinal cord. However, classical tractography methods do not restrict end-points to finish in two anatomically plausible regions. This limitation causes streamlines to end prematurely inside the WM or invalid regions, such as the ventricles. In addition, current structural connectivity estimation relies on voxel-based masks and atlas labels, which are limited by image resolution, partial volume effects, and the staircase effect (voxelic discretization) (Calamante, 2019; Côté et al., 2013; Roine et al., 2014). Most of these limitations directly impact the quality of the connectivity mapping.
To mitigate these effects in connectivity analysis, GM labels are often dilated or the GM label closest to the streamline end-point is chosen (Yeh et al., 2020). However, these approaches can bias results by unfairly increasing the likelihood that streamlines will be labeled within a GM area, particularly by those closer to deep WM (Schilling et al., 2017; Van Essen et al., 2014). Recently, to mitigate these limitations, multiple tractography methods have been proposed to enforce streamlines to restrict both end-points within GM voxels or onto cortical surfaces (Girard et al., 2014; Smith et al., 2012; St-Onge et al., 2018; Teillac et al., 2017; Yeh et al., 2017).
Background
State-of-the-art tractography methods, such as anatomically constrained tractography (ACT) (Smith et al., 2012) and particle filtering tractography (PFT) (Girard et al., 2014), take advantage of probabilistic brain tissue segmentation, also called continuous map criterion, to improve streamline reconstruction near the tissues' interface. Afterward, these algorithms discriminate streamline reconstruction based on tissue maps, and streamlines going through invalid regions are discarded.
Interface seeding is often used for connectivity analysis since it requires both end-points of a streamline to reach GM regions (or the brainstem). This seeding method is preferable, because WM seeding has difficulty reaching GM at both end-points (Girard et al., 2014). Also, streamline end-points generated from WM seeding tend to be concentrated in small regions along the cortex, an effect that is accentuated when using deterministic tractography (St-Onge et al., 2018). WM seeding is suitable to reconstruct dense WM structures and bundles, but inadequate for cortical measures along end-points and connectivity analysis. Both ACT and PFT can be used to initialize streamlines from a WM–GM interface seeding mask instead of WM seeding.
Recently, these tractography algorithms were adapted to integrate cortical meshes, for example, mesh-based ACT (Yeh et al., 2017) and surface-enhanced tractography (SET) (St-Onge et al., 2018). This incorporation of surfaces was also utilized to improve a global tractography method (Teillac et al., 2017). These cortical surfaces, extracted from a T1 image with higher spatial resolution, are used to reduce voxel discretization effects and improve tractography precision close to the GM. The integration of cortical surfaces with tractography was initially proposed by Cottaar et al. (2015) and St-Onge et al. (2015), to better reconstruct the streamlines projection near the cortex, especially for the fanning structure in gyri.
Figure 1 illustrates tractography WM–GM interface seeding approaches; from using voxel-based interface mask to surface-based with and without surface flow. Figure 2 presents termination methods to map streamlines to the cortex: GM voxel-based mask, nearest position, and surface intersection.
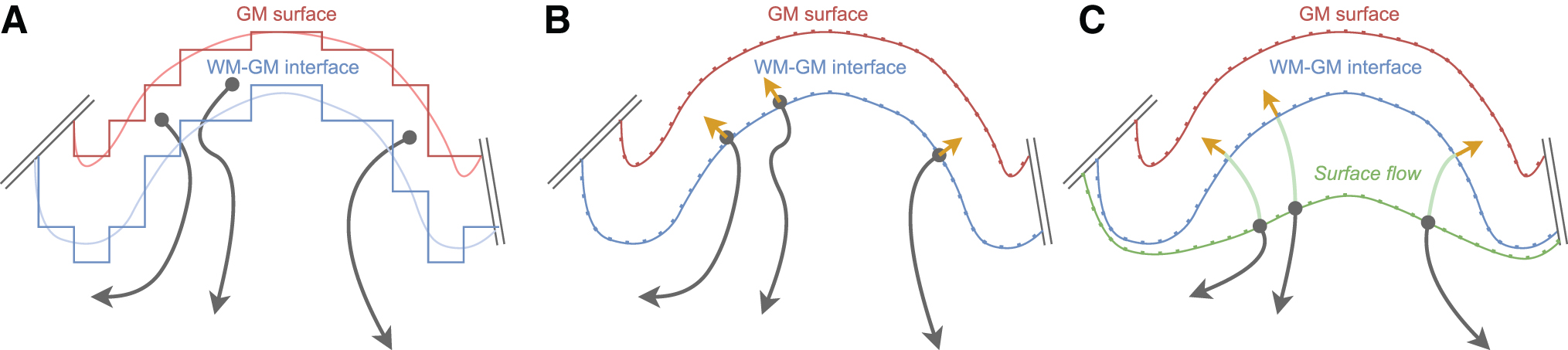
Streamlines interface seeding approaches:
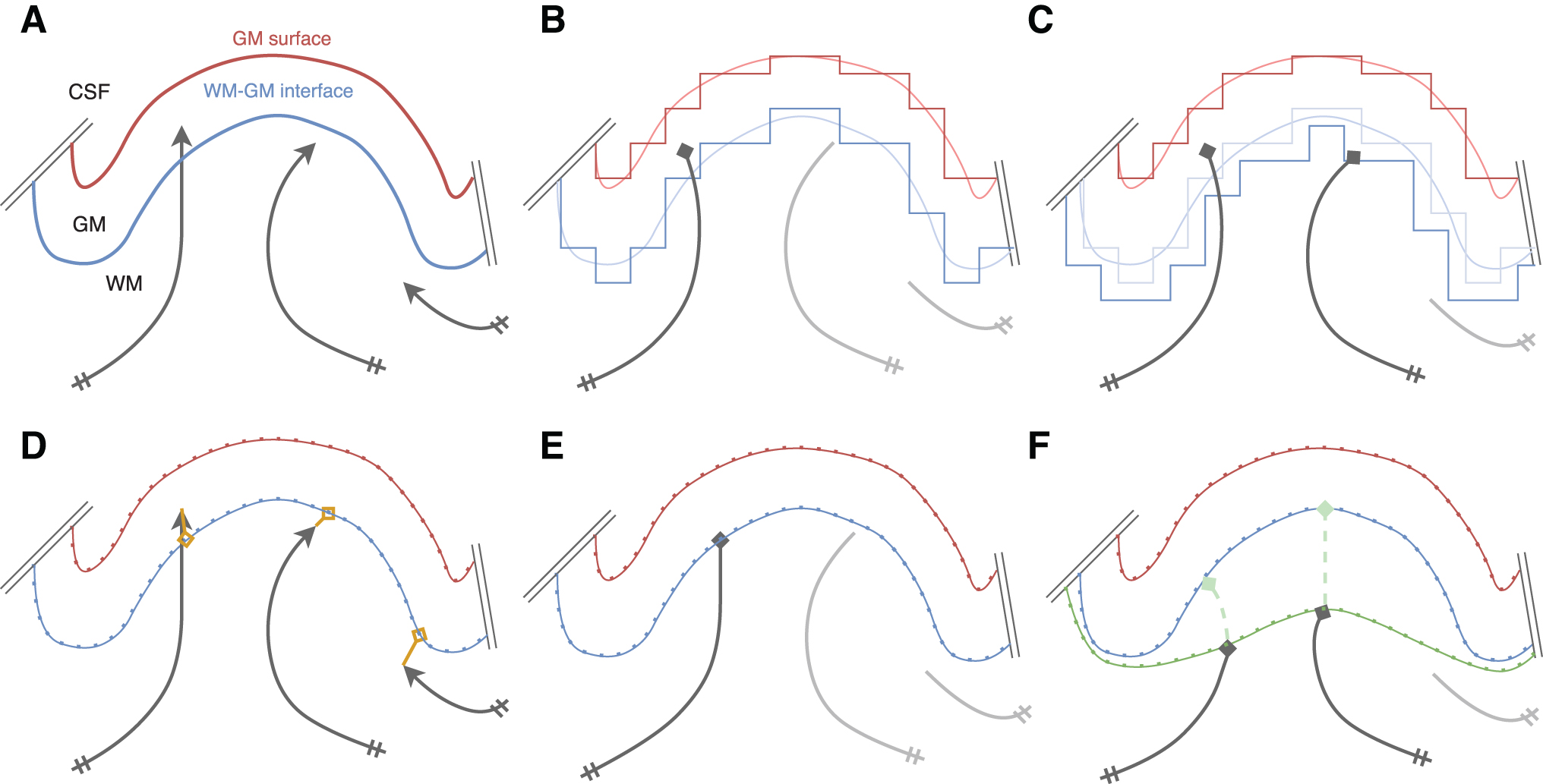
Streamline termination approaches:
Motivation
The focus of this study is to improve cortical coverage from tractography end-points with the use of cortical meshes. For this purpose, we suggest an adaptive and dynamic initialization along cortical surfaces to reduce streamline end-point bias and end-point variability. Resulting end-point distributions along the cortex were analyzed to estimate the connectivity variability. In addition, different approaches generating connectivity matrices from tractography end-points are compared, from voxel-based atlas to surface-based parcellation.
Methods
Adaptive and dynamic surface seeding
In this study, two novel tractography seeding strategies are proposed to improve cortical coverage. First, the adaptive surface seeding gives more initialization flexibility, allowing the user to specify the starting points distribution along the cortex (Fig. 3). The local cortical maps (e.g., thickness) can be given to estimate a specific end-point's connectivity, based on the GM volume distribution. Afterward, to initialize streamlines, the list of seeding positions is randomly chosen from this distribution on the mesh triangles. A mask of a region of interest (ROI) can also be used to focus tractography in a specific location along the cortex.

Adaptive seeding maps:
Second, the dynamic surface seeding is an iterative version of the adaptive method, to approach the given initial distribution. Since initial positions are given but terminations are determined by the tractography algorithm, seeding uniformly across the surface does not result in a uniform end-point distribution. Dynamic seeding is applied to iteratively adapt seeding points toward nonconnected areas of the cortex. Illustrated in Figure 4, the iterative seeding can be decomposed in four major steps:
Initialize seeding distribution along the cortex, that is, uniformly along the cortex, based on the local GM thickness/volume or specified regions of interest. Then, start tractography streamlines from these seed points.
Estimate the tractography end-point distribution/density along the cortical surface.
Generate a new tractography seeding map from the difference between the desired initial seeding distribution and the combination of all previous end-point distributions. Launch a new tractography from this new seeding distribution map.
Repeat steps II and III until reached a predetermined number of iterations is reached for the dynamic surface seeding. The resulting tractogram is the combination of all resulting tractography streamlines.
Similar iterative approaches were previously proposed (Bauer et al., 2013; Dhollander et al., 2014; Smith et al., 2015a), but only as applied to standard voxel-based seeding. Having the flexibility to choose the seeding distribution along the cortical surface is a substantial way to reduce specific connectivity biases.
Subcortical structures and ROI seeding
Subcortical structures seeding was also introduced to better explore WM structures connecting these regions. The adaptive seeding area used on subcortical meshes is determined by a WM segmentation or fractional anisotropy (FA) map, comparable with WM seeding along ROIs interface. Then, streamlines are initialized toward the WM, using the main diffusion direction. Once the tractography is completed, subcortical structure density can be estimated along interface meshes, on a per subject basis. The proposed dynamic seeding can also be used when providing a distribution per subcortical surface. Streamline terminations along these surfaces are computed employing the same technique used for the cortex mapping (nearest vertex [NV], intersection, etc.).
Experiments
Data sets and processing
Diffusion MRI
The Human Connectome Project (HCP) (Van Essen et al., 2013) test–retest data set was used to compare the variability of the different connectivity approaches, using the mean and standard deviation (SD). From the reproducibility database, detailed in Glasser et al. (2013), 88 acquisitions were used, from 44 healthy participants at two time points. Tractoflow (Theaud et al., 2020) was employed to estimate diffusion maps required for the tractography, for example, FA, probabilistic T1 tissues maps, and fiber orientation distribution function. Code implementation, parameters, and details are provided in Section 6.
Surface reconstruction
For every acquisition of the test–retest data set, cortical surfaces and parcellations were generated with CIVET (Kim et al., 2005; Lyttelton et al., 2007). The surface flow was used with a total time of 100 and an implicit time step of 1, as described in St-Onge et al. (2018). Subcortical structures were extracted from the CIVET voxelic template, and converted into surfaces using marching cubes (Lorensen and Cline, 1987).
Tractography
For all comparisons, streamlines were reconstructed inside the WM using PFT (Girard et al., 2014) available from DIPY (Garyfallidis et al., 2014). As mentioned before, PFT can be used with multiple seeding methods: WM seeding (voxel-based), interface seeding (voxel-based, which is default), and surface seeding with or without surface flow. For voxel-based methods, streamlines were seeded at the WM–GM interface and stopped in GM, refer to Girard et al. (2014) for details. For surface-based approaches, tractography streamlines were initialized and terminated in cortical surfaces (WM–GM interface mesh). With the surface-enhanced approach (St-Onge et al., 2018), the surface flow trajectory was used to initialize and back-project tractography streamlines over cortical surfaces. For all methods, features extracted from resulting tractograms were projected along the cortical mesh, based on streamline end-points.
Evaluation
To estimate potential biases, such as the gyral bias (Reveley et al., 2015; Schilling et al., 2017; Van Essen et al., 2014), we studied tractogram end-point distribution along the WM–GM interface. Surface measurements, alongside tractogram connections (streamlines that end in GM), were used to compare tractography algorithms. These measurements include the tractogram cortical coverage, the number of connections at each vertex (end-point density) and the mean curvature on the surface at those vertices. The mean curvature was used to separate gyri from sulci (Dale et al., 1999; Fischl et al., 2004), to afterward estimate the gyral bias for each approach. Any streamline that did not reach these surfaces, stopping prematurely inside WM without reaching GM, were considered invalid and removed from the tractogram. An initial intrasubject average and SD was computed over each tractogram of the same subject. Afterward, a global average and intersubject SD was computed from the subject-wise average. Since the SD (σ) is strongly correlated with the average value (µ), the variability is presented using the coefficient of variation (CoV), to measure dispersion relative to the mean (CoV =σ/µ).
Different streamlines to surface mapping approaches were compared, employing interface seeding (PFT) or surface seeding (SET), illustrated in Figures 1 and 2: PFT using surface intersection (SI) PFT using surface intersection with surface flow (SFI) PFT using surface NV with a maximum of 2 mm distance (NV) SET using a uniform seeding (SET) SET using the proposed dynamic seeding (SET dyn).
Test–retest variability was compared using analysis of variance (sum of squares) from reconstructed DW-MRI connectivity matrices. This was done to assess whether a change in variability appeared between connectivity matrix reconstruction methods. Approaches that take the nearest label or dilate labels are common in connectivity matrix reconstruction to increase the chance that streamlines reach labels (Smith et al., 2015b; Yeh et al., 2020; Zhang et al., 2018). For connectivity matrix reconstruction, three parcellation/binning methods were evaluated:
Voxelic parcellation included by HCP
Voxelic parcellation included by HCP, dilated by 2 mm
Surface parcellation, computed with CIVET. For PFT, the mapping of streamlines end-points to the NV of the surface, within 2 mm, was employed to map streamlines to surfaces.
Results
Cortical coverage
Figure 5 presents the progression of cortical surface coverage measured in percentages. It is important to note that this graph is a function of the number of valid streamlines (with both end-points reaching GM) generated with each approach. It can be observed that both methods employing surface intersections for streamline end-point mapping, with surface flow (SFI) and without (SI), do not reach 90%. Compared with the 2 mm NV projection reaching 92%, surface seeding approaches are able to increase coverage significantly (p < 0.001) to 94%(SET) and 96%(SET dyn) after a million valid streamlines (Appendix Table A1). The percentage of seeded streamlines resulting in a valid streamline is detailed in Appendix Table A2, and the coverage as a function of the number of seeds is presented in Appendix Figure A1.
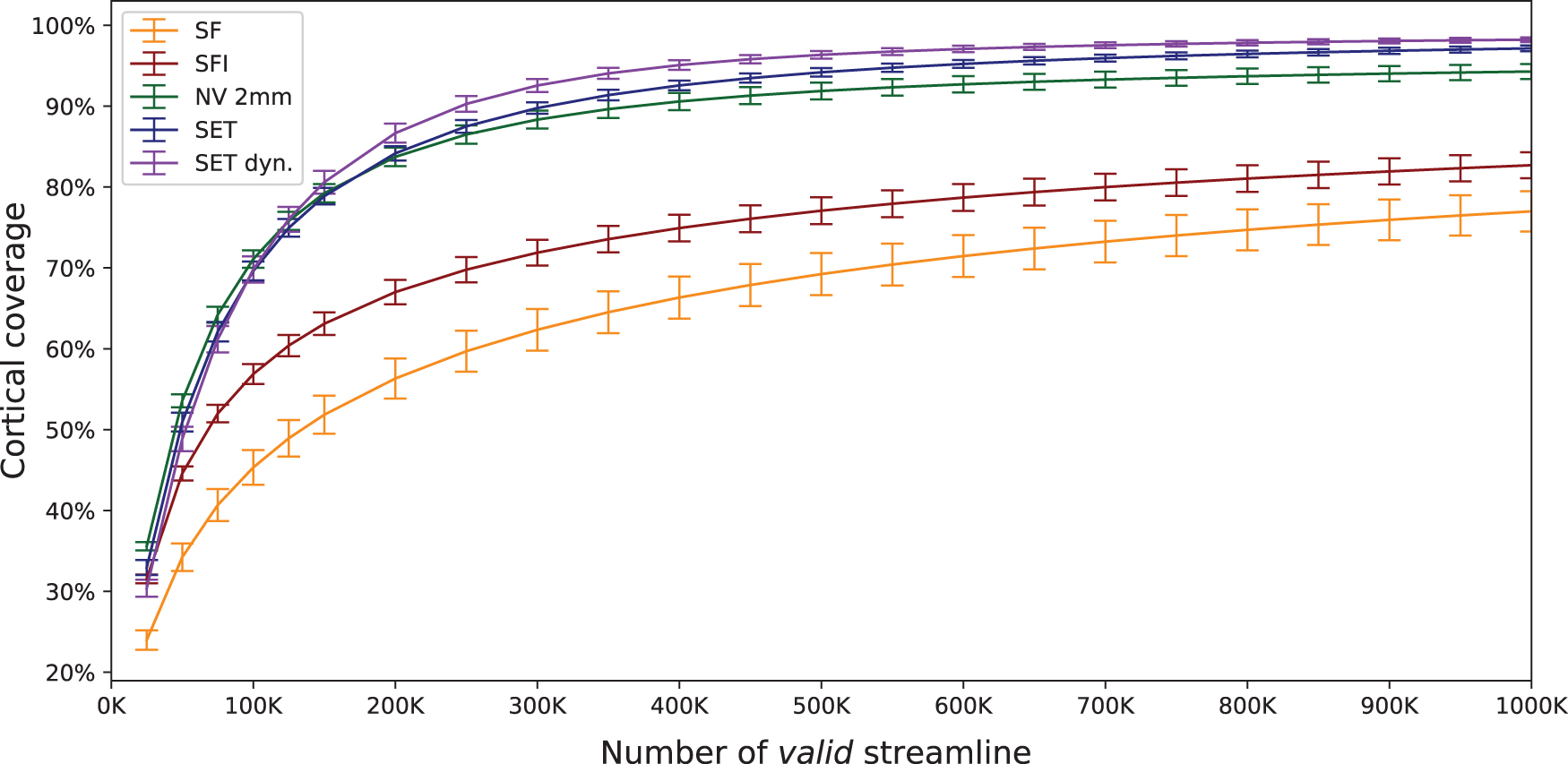
Percentage of cortical surface coverage with respect to the number of resulting streamlines reaching two GM regions. GM, gray matter. Color images are available online.
End-point distribution
The streamline end-point distributions, along cortical surfaces, can be observed in Figure 6, averaged over all subjects for each mapping method. The surface flow increases cortical coverage, but end-points along the cortex remain uneven. NV, SET, and SET dynamic further improve the uniformity of the averaged end-point distribution. SET dynamic results in an overall lower intrasubject CoV, averaged along the cortex. An estimation of this nonuniformity was measured with the SD of the resulting density at each position (vertex) Appendix Figure A2.
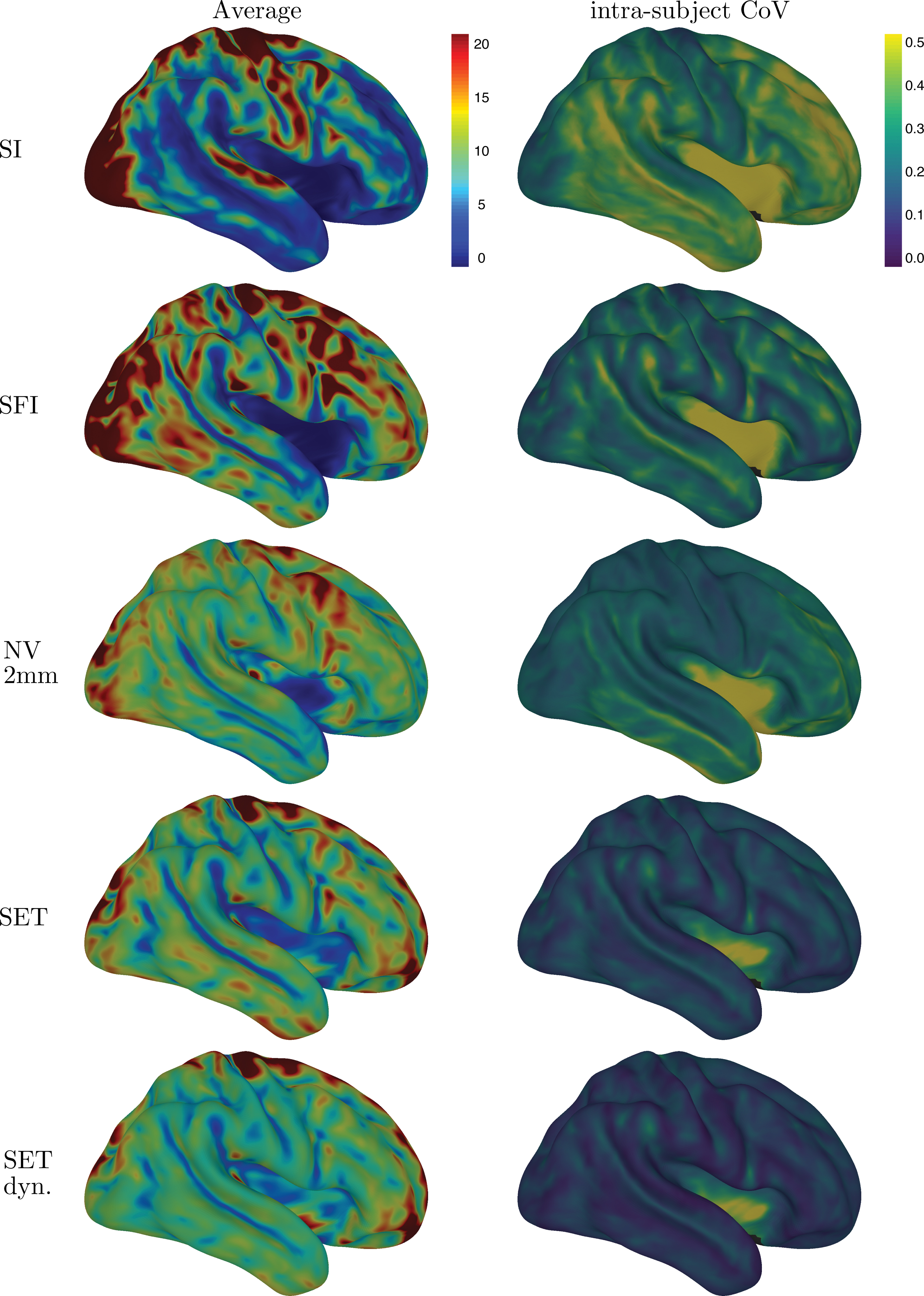
Surface end-point density per million streamlines, subjects average and intrasubject CoV. Interface seeding tractography with SI, SFI, and NV. CoV, coefficient of variation; SET, uniform surface seeding; SET dyn, dynamic surface seeding. Color images are available online.
Surface measurements nearby end-points
Figure 7 shows the percentage of end-points in positive mean curvature (gyri) and the distribution of mean curvature for each method. Even where surface flow (SFI) is able to reduce the percentage (SI) by 10%, both methods are strongly biased to end in gyri crowns. SET with the dynamic seeding approach is able to reduce this percentage to 58.7%. The NV mapping 2 mm is on average at 48.4% end-points in gyri, thus slightly over-representing sulci (equivalent to 51.6% end-points in sulci).
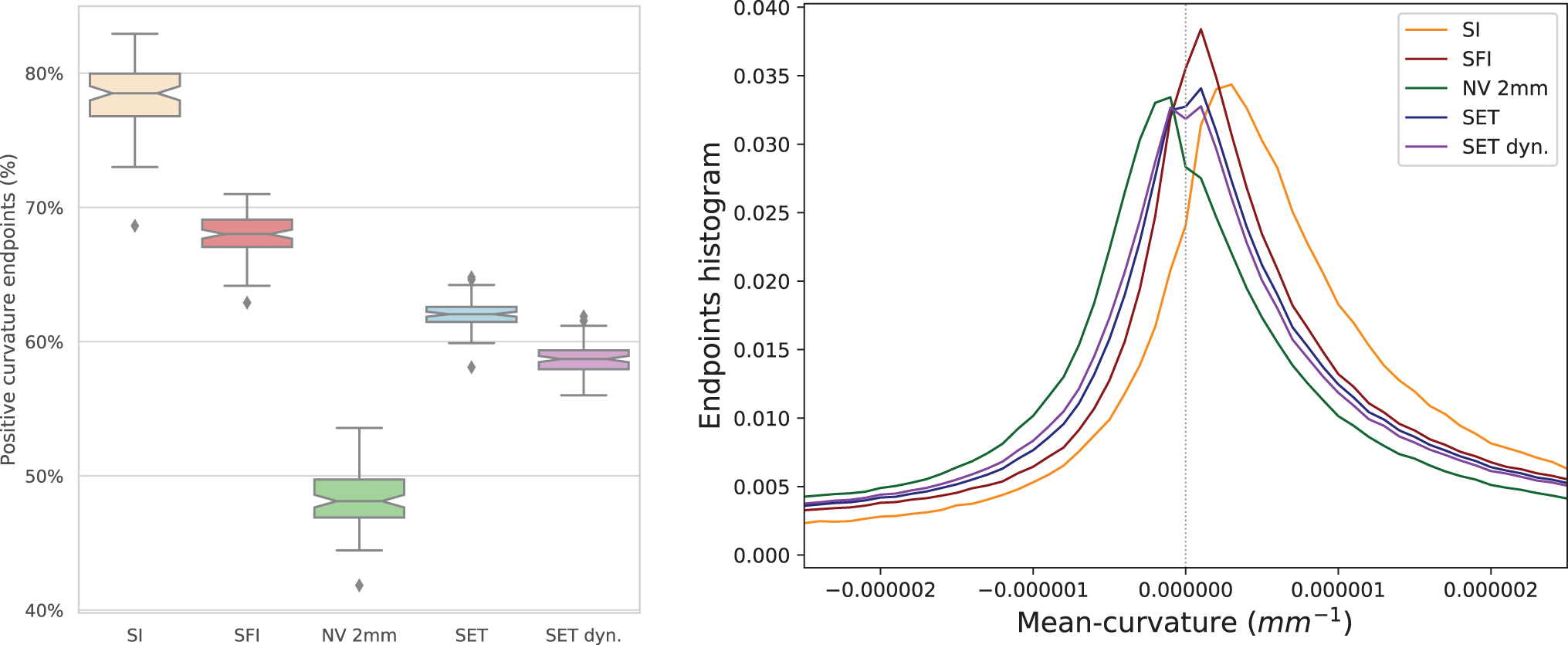
Positive mean-curvature percentage representing the gyri/sulci bias, with the mean-curvature histogram, for each method. Color images are available online.
Variability
Figure 8 illustrates the connectivity average (first row) and intrasubject CoV (second row), for five matrix reconstruction methods. The sum squared ratio (intra/total) for the same approaches is presented in Figure 8B. A lower intrasubject variability, relative to the intersubject variability, results in a lower ratio. Surface-based connectivity significantly (p < 0.001) (NV, SET) reduces this ratio compared with the nondilated voxelic atlas method. However, there is no significant improvement over the dilated voxelic version of the template (Appendix Figure A3). The averaged intrasubjects variability is shown in Figure 6 for end-points along the cortex, and in Appendix Figure A5 for connectivity matrices.
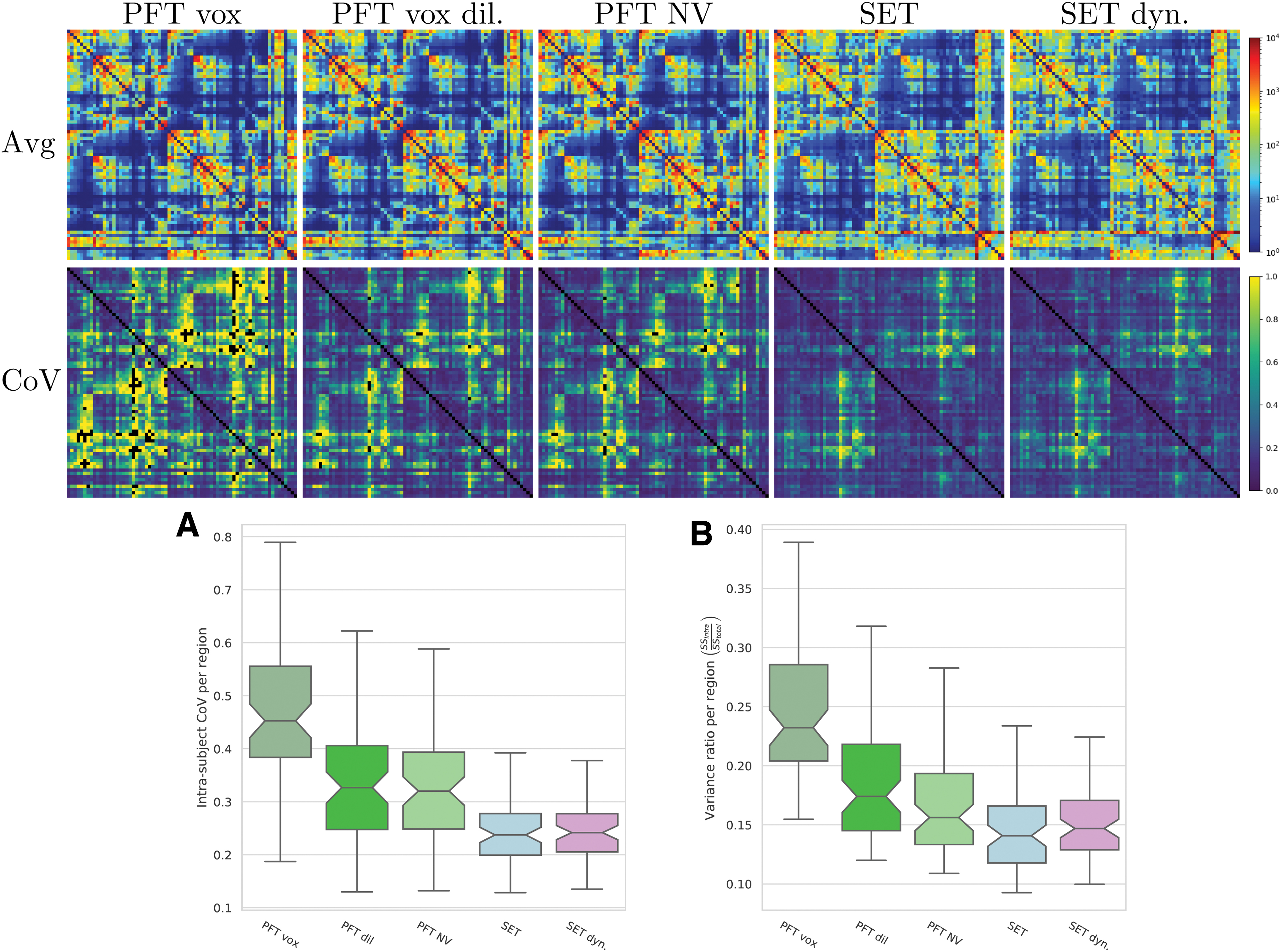
The first two lines are connectivity matrices per million connections: subjects average and intrasubject CoV. Quantitatives variability of connectivity matrices:
Discussion
Utilizing mesh surfaces for streamline termination not only improves end-point precision and coverage, but also facilitates the analysis of WM features projected onto the cortex, such as end-point density, coverage, and gyral bias. Recent methods that start and finish along the cortex improve the surface distribution and coverage, but might also suffer from the sensitivity/specificity trade-off (Girard et al., 2020). Further validation from histology will be crucial to assess the precision of reconstructed structural density and connectivity.
Cortical coverage and end-point distribution
Streamline intersection with surfaces (SI), is very restrictive for streamlines resulting in low-coverage and high-density regions. Surface flow intersection (SFI) moderately improves the coverage and density variation, but is far from matching surface seeding approaches. NV mapping gives overall good results and a high coverage. The NV approach only requires standard maps for tractography (PFT), so integrating surfaces can be done post-tracking only to map end-points along cortical surfaces, which can be advantageous. Surface seeding methods (SET, SET dynamic) require cortical surfaces before tracking to uniformly initialize streamlines along the cortex. SET increases the end-point coverage and uniformity, especially compared with other intersection approaches (SI, SFI). The dynamic seeding further improves SET coverage and uniformity results.
Gyral bias
The proposed adaptive and dynamic seeding improve the coverage and reduce the gyral bias. However, there is still room for improvement and, even when seeding from the whole surface uniformly (SET), streamlines finish more often in gyri (62%) than in sulci (38%) with 3% of the cortex left without connections. SET with dynamic seeding further reduces this bias (59%) by forcing a certain uniformity in seeding points, but resulting streamline terminations are still biased toward gyri. Alternatively, the NV mapping slightly favors sulci (48% gyri), since there are more deep WM areas that pass nearby sulci banks, within the 2 mm, which get a termination assignment there. This phenomenon can be observed in Figure 2C and D. Nevertheless, mapping with the surface flow (SFI or SET) recovers a full trajectory from the superficial WM to the cortex, instead of simply connecting to the nearest GM position. This trajectory is generated using a geometric flow at the position of the streamline intersection with the surface (Fig. 2F). It is hard to determine the anatomical validity of streamline terminations in sulci banks, especially when produced by NV mapping of 2 mm. Validation with other techniques such as tract-tracing could be important in the future.
The exact distribution of axon termination along the cortex in the human brain is not precisely known and varies across subjects. The fiber density is often assumed to be relatively proportional to the unit volume of the cortex, implying that the gyral crown percentage would be higher than in sulci by a few percent (Donahue et al., 2016; Van Essen et al., 2014). Recent research on primates Schilling et al. (2017) suggests that the gyri crown to wall ratio is around 1.13 (53%) and that histologically the fiber density profile is not the same across all gyri.
Variability
Mapping with NV is a good alternative to standard intersection approaches (SI, SFI) to reduce the variability without needing to include surfaces before tractography. Nonetheless, surface reconstructions (e.g., Freesurfer) are widely available, easy to incorporate and often used to recover atlas segmentation or segmentation maps used along with tractography. Integrating those cortical surfaces, before the tractography, with uniform surface seeding reduces the intrasubject variance for both end-point density and connectivity matrices. Finally, the proposed dynamic seeding further reduces the intrasubject variability, which may be crucial for connectivity analysis.
In Figure 6, low and high end-point variability can be observed from intrasubject coefficients of variation. For all methods, the insular area and sulci depth tend to have higher variability. Moreover, connectivity variability is very high in some specific regions (Fig. 8), such as the anterior cingulate (medial and rostral), the entorhinal cortex, the medial orbitofrontal cortex, the insula, and the globus pallidus. These regions have similar CoV and sum squared ratios (SSintra/SStotal) for both hemispheres.
Future studies
By giving an appropriate distribution, based on prior knowledge of anatomy or given regions of interest, the proposed adaptive and dynamic surface seeding methods could be used to reduce specific connectivity biases. Cortical surface local features such as area, thickness, and volume, among others, could be included with the adaptive seeding strategy to iteratively fill GM regions with no end-points. It is important to reduce the effect of streamline end-point bias and validate results, with test–retest variability, before performing connectivity analysis. The dynamic surface seeding could be, furthermore, improved by integrating other types of priors, such as SIFT (Smith et al., 2013) or a version of COMMIT (Daducci et al., 2015) based on cortical coverage and density. Microstructure-informed tractography could further reduce biases in streamline distributions (Daducci et al., 2016; Girard et al., 2017).
Conclusion
Utilizing mesh surfaces for streamline termination improves end-point precision and coverage, and facilitates the analysis of WM features projected onto the cortex, such as end-point density, coverage, and gyral bias. The proposed adaptive seeding with a uniform initialization significantly improves tractography coverage along the cortex. The dynamic approach further improves the cortical coverage with reduced gyral bias from tractography end-points. The integration of a surface-based atlas, with a proper mapping method (NV, SET, and SET dyn), can increase the test–retest reproducibility of connectivity matrices. This can boost future structural brain connectivity and the application of connectomics in the research.
Code Availability
The proposed surface-tractography integration and the SET processing pipeline is available online with a documentation page. This method requires a surface reconstruction from CIVET or alternatively FreeSurfer. The pipeline already includes a diffusion MRI processing pipeline Tractoflow-PVE to incorporate segmentation maps from CIVET, based on Tractoflow.
Footnotes
Acknowledgments
We thank Alessandro Daducci, Kevin Whittingstall, and François Rheault for their help and insights.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Thanks to the Fonds de recherche du Québec—Nature et technologies (FRQNT) and Natural Sciences and Engineering Research Council of Canada (NSERC) for research funding. A special thanks to the Neuroinformatics Chair of the Sherbrooke University, which helped push forward neurosciences research.