
Abstract
Introduction:
Three prominent resting-state networks (rsNW) (default mode network [DMN], salience network [SN], and central executive network [CEN]) are recognized for their important role in several neuropsychiatric conditions. However, our understanding of their relevance in terms of cognition remains insufficient.
Materials and Methods:
In response, this study aims at investigating the patterns of different network properties (resting-state activity [RSA] and short- and long-range functional connectivity [FC]) in these three core rsNWs, as well as the dynamics of age-associated changes and their relation to cognitive performance in a sample of healthy controls (N = 74) covering a large age span (20–79 years). Using a whole-network based approach, three measures were calculated from the functional magnetic resonance imaging (fMRI) data: amplitude of low-frequency fluctuations (ALFF), regional homogeneity (ReHo), and degree of network centrality (DC). The cognitive test battery covered the following domains: memory, executive functioning, processing speed, attention, and visual perception.
Results:
For all three fMRI measures (ALFF, ReHo, and DC), the highest values of spontaneous brain activity (ALFF), short- and long-range connectivity (ReHo, DC) were observed in the DMN and the lowest in the SN. Significant age-associated decrease was observed in the DMN for ALFF and DC, and in the SN for ALFF and ReHo. Significant negative partial correlations were observed for working memory and ALFF in all three networks, as well as for additional cognitive parameters and ALFF in CEN.
Discussion:
Our results show that higher RSA in the three core rsNWs may have an unfavorable effect on cognition. Conversely, the pattern of network properties in healthy subjects included low RSA and FC in the SN. This complements previous research related to the three core rsNW and shows that the chosen approach can provide additional insight into their function.
Impact statement
Using a whole network-based approach, our study characterizes the normal patterns (including resting-state activity [RSA], short- and long-range functional connectivity [FC]) of three prominent resting-state networks (rsNW) within the context of age-dependent changes and explores their relevance for different cognitive domains. Our results revealed a pattern with low RSA and FC in the salience network in healthy volunteers, whereas higher RSA, particularly in the central executive network, seemed to have a negative effect on cognition. These results increase the knowledge about the three core rsNWs and the understanding about their relevance for cognition.
Introduction
The global trend toward increasing life expectancy has prompted many neuroscientists to focus on the investigation of age-related changes in cognition and the corresponding underlying neurobiological correlates. To this end, most research approaches have relied on a modular paradigms, where brain areas are postulated to act as independent processors for specific complex cognitive functions (Bressler and Menon, 2010). However, in response to several shortcomings in these concepts (Fuster, 2000), there has been a redirection in methodology away from the simplistic mapping of circuits involved in cognition into individual brain areas toward the conjoint function of brain areas working together as large-scale networks (Bressler and Menon, 2010).
Our understanding of these networks has been significantly broadened in the past decade through the continued technical improvement of resting-state functional magnetic resonance imaging (rsfMRI), a method used to measure intrinsic brain activity when participants are not engaged in any particular action (task-free state). Brain regions, strongly functionally linked during rest, are commonly summarized as resting-state networks (rsNW) (Biswal et al., 1995, 1997). Within the several stable rsNWs identified in recent years (Raichle, 2015), three networks stand out for their synchronized interplay and their importance: the default mode network (DMN), the central executive network (CEN), and the salience network (SN). These three networks are often jointly referred to as the triple network model (Menon, 2011). They are also known as the core elements of neurocognitive networks due to their engagement in a wide range of cognitive tasks (Menon and Uddin, 2010). Further, several rsfMRI studies have shown that disruption in the coordinated activity of the three core rsNWs is a crucial factor in various psychiatric diseases (Dong et al., 2017; Menon, 2011), which often show partially overlapping dysfunctions particularly in those three networks. The observations mentioned substantiate the hypothesis that alterations in these three core rsNWs might mirror conclusively the influence of multiple etiological factors on neuropsychiatric disorders (Menon, 2019).
Despite insights into the predictive value of the three core rsNWs for psychopathological/disease-relevant conditions, our understanding of their relevance in terms of cognition is limited (Mill et al., 2020). Knowledge in this context is mainly derived from clinical research linking resting-state functional connectivity (rsFC) alterations to pathology (Buckholtz and Meyer-Lindenberg, 2012) and from the investigation of rsFC changes at a whole-brain level in association with various forms of healthy and unhealthy aging (Andrews-Hanna et al., 2010; Geerligs et al., 2015). However, the question of whether other concrete characteristics of the rsNW, beyond classical connectivity, have a relevant influence on certain cognitive domains and, particularly, whether this influence can be attributed entirely to age-related network alterations, remains unanswered. In response to this question, we chose to investigate patterns of different network properties within the three core rsNWs as well as the dynamics of age-associated changes and their relation to cognitive performance in a sample of healthy controls during the adult lifespan.
The resting-state examinations may be used for calculation of several different measures of rsFC, each representing a separate property of brain networks. For this approach, we chose three such measures: the amplitude of low-frequency fluctuations (ALFF) (Zang et al., 2007), the regional homogeneity (ReHo) (Zang et al., 2004), and the degree of network centrality (DC) (Zuo et al., 2012).
The ALFF is considered to be an indicator of the local intensity of spontaneous fluctuations in the blood oxygen level-dependent signal (BOLD), which represents the spontaneous neural activity of specific regions and physiological states of the brain (Zang et al., 2007). It is defined as the total power within the low frequency range between 0.01 and 0.1 Hz (Biswal et al., 1995). Several ALFF studies suggest that intrinsic resting-state activity (RSA) promotes or allows specific brain circuits to participate in cognitive tasks, consecutively predicting task-induced brain responses and behavioral performance (Wang et al., 2019).
ReHo is used commonly as a voxel-based parameter to characterize functional homogeneity of rsfMRI signals within a small region (Zang et al., 2004). It reflects the degree of synchronization between a given voxel and its nearest 26 neighboring voxels time series. Kendall's coefficient of concordance (KCC) (Kendall, 1990) is used as an index to measure the degree of this synchronization. Thereby, it is assumed that a given voxel is temporally similar to that of its neighbors and that ReHo could be modulated in pertinent cognitive tasks (Zang et al., 2004).
DC (Joyce et al., 2010), often referred to as global functional connectivity (FC) density (Tomasi and Volkow, 2012), characterizes the correlation of the functional patterns of the BOLD signal between spatially distant brain areas at a network level. This is a measure of FC, which mirrors the number of direct functional connections between a given voxel and all the other voxels in the brain. The estimation of the DC is carried out by the Pearson's correlation coefficient. DC is useful for revealing the cortical functional hubs (Buckner et al., 2009; Zuo et al., 2012).
DC and ReHo complement each other to detect both remote and local activity synchronization in the brain (Cui et al., 2016). Together with the ALFF, they may be used for a comprehensive rsNW characterization and a display of patterns comprising spontaneous brain activity, regional temporal integration, and connectivity.
For the characterization of cognitive performance, a test battery was compiled to represent key central cognitive domains: processing speed, memory, attention, executive functioning, and visual perception (Harvey et al., 2019). We expected the most pronounced age effects on short-range connectivity. Further, we hypothesized that these connectivity changes would be related to cognitive decline in an age-dependent manner.
Materials and Methods
Participants
Healthy volunteers (mean years of education 11.78 [standard deviation; SD: 1.24], range between 9 and 13 years) were recruited for the study through flyers on notice boards. Participants were without a history of psychiatric or neurological diseases, were right-handers and had a normal or corrected-to-normal vision. The screening for mental health was performed before inclusion by using the Structured Clinical Interview for DSM-IV (First et al., 2002). People with current and previous substance abuse were not eligible for inclusion. Physical and neurological health was assessed by clinical examination. Five individuals were excluded due to left-handedness, as indicated by the Edinburgh inventory (Oldfield, 1971).
The final sample consisted of 74 (35 female [47.3%]) German-speaking participants aged 20 to 79 years (mean age 47.26, SD 19.06), where both behavioral and imaging data were collected. Verbal and written informed consent was obtained from all participants. The study was conducted according to the Declaration of Helsinki and was approved by the Medical Ethics Committee of the RWTH Aachen University.
Following the functional magnetic resonance imaging (fMRI)-measurement (45 min), participants performed a range of well-established neuropsychological tasks. The instructions for the tasks were given by experienced neuropsychologists who strictly adhered to the official protocols (∼3 h, with one break in the middle of testing). At the end of the session, the participants received monetary compensation.
Assessment of neurocognitive performance
The neurocognitive test-battery was composed to examine the following cognitive domains (Harvey et al., 2019): memory, executive functioning, processing speed, attention, and visual perception. Therefore, the following tests were used:
The German version of the Wechsler Memory Scale—Revised (WMS-R) (Härting et al., 2000) enables the capture of different subcategories of memory performance by using several subtests. The first subtest, which assesses cognitive capability to perform all other tests, was omitted, as only healthy adults were included in the study. Nine subtests were administered in total, providing scores for verbal memory (subtests: “Logical memory” and “Verbal paired associate learning”), visual memory (“Figural memory,” “Visual paired associate learning,” and “Visual reproduction”), and delayed memory (subtests “Logical memory—delayed,” “Verbal paired associate learning—delayed,” “Visual paired associate learning—delayed,” and “Visual reproduction—delayed”).
The digit span test from the Wechsler Adult Intelligence Scale—III (WAIS-III) (Von Aster et al., 2006) was included to assess working memory. In the test, a sequence of numbers is read out to the participants and they are asked to repeat the same sequence in the same (forward span) or in the reverse order (backward span). Forward span captures attention efficiency and capacity. Backward span is an executive task that is particularly dependent on working memory. The scores were reported separately for forward and backward performance.
The 64-cards version of the Wisconsin Card Sorting Test (WCST) (Kongs et al., 2000) is considered to be the gold standard for the clinical assessment of executive function (Rabin et al., 2005). The test performance involves a range of different functions, such as set switching, error detection, attention, working memory, abstraction, and decision making (Nyhus and Barcelo, 2009). Studies using factor analysis to elucidate the validity of the test resulted in two main factors (the first relating to mental flexibility and the second relating to memory, attention, and motivation) being mainly represented by the number of perseverative errors and the number of nonperseverative errors (Greve et al., 1998).
The Trail Making Test (TMT) (Reitan, 1958) is an extensively used neuropsychological instrument considered to measure processing speed, sequencing, mental flexibility, and visual–motor skills (Bowie and Harvey, 2006). Performance time in TMT-A is believed to provide a baseline measure of psychomotor speed, visuospatial search/visual perception, and target-directed motor tracking (Varjacic et al., 2018). Part B is more complex and additionally assesses set-switching and working memory. The difference score (TMT-B–TMT-A) minimizes visuo-perceptual and working memory demands and is considered to be a relatively pure indicator of executive control abilities (Sanchez-Cubillo et al., 2009).
Further, for processing speed, the prototypical number symbol substitution test from the WAIS-III (Von Aster et al., 2006) was included. Participants are asked to pair symbols to numbers by using a predefined encoding kay sequence outlined at the top of the examination sheet. The coding should have been carried out as quickly as possible, and the result is the number of correct answers in the given time (120 sec). The test primarily quantifies not only the speed of processing but also cognitive flexibility, concentration, short-term visual memory, attention, learning ability, and motivation.
Attention and visual perception were investigated by using the subtests Alertness, Divided Attention, Flexibility, and Visual Scanning of computerized Test of Attentional Performance (TAP) (Testbatterie zur Aufmerksamkeitsprüfung (Zimmermann and Fimm, 1993).
Intelligence was measured with the German version of the WAIS-III (Von Aster et al., 2006) to confirm the inclusion criterion of average intelligence.
Functional magnetic resonance imaging
Data acquisition
The MRI data were acquired by using a 3-Tesla Magnetom Trio (Siemens Medical Solutions, Erlangen, Germany). All participants received a whole-brain scan that included structural and rsfMRI measurements. Structural measurements were performed by using a magnetization-prepared rapid acquisition gradient-echo sequence, resulting in T1-weighted anatomical images, with echo time (TE) = 3.03 msec, repetition time (TR) = 2250 msec, field-of-view (FOV) 256 × 256 × 176 mm3, flip angle (FA) = 9°, 176 sagittal slices, voxel size = 1 × 1 × 1 mm3, and a GeneRalized Autocalibrating Partial Parallel Acquisition acceleration factor of 2 with 70 auto-calibration signal lines.
The rsfMRI measurement was performed by using T2*-weighted echo-planar imaging, with TE = 30 msec. TR = 2200 msec. FOV = 200 × 200 × 108 mm3, FA = 80°, number of slices = 36, voxel size = 3.125 × 3.125 × 3.0 mm3, and number of volumes = 180. During the rsfMRI measurements, participants were asked to lie motionless in the scanner for 6 min with their eyes closed, without doing or thinking of anything specific (let their minds wander).
fMRI data processing
Data were processed by using Matlab based software packages SPM12 (
Subsequent to the pre-processing steps, FC measures were calculated at voxel level for each subject. Temporal filtering of the BOLD fMRI signal between 0.01 and 0.08 Hz was only performed before ReHo and DC calculation. The ALFF measures were calculated between 0.01 and 0.1 Hz of the BOLD fMRI signal (Biswal et al., 1995, 2010; Zuo et al., 2010). The ReHo connectivity measure was calculated over a cluster of 27 neighboring voxels (Li et al., 2012) by using KCC as the homogeneity metric. The DC measure was computed based on a Pearson correlation coefficient cut-off of 0.25 (p = 0.001).
To allow statistical comparisons, the obtained measures were linearly standardized into Z-values for each subject (Aiello et al., 2015) by subtracting the mean whole-brain voxel value from each voxel and then dividing the difference by the SD of the whole brain. These standardized connectivity measures were co-registered to the MNI152 (2 × 2 × 2 mm3) standard space and smoothed with a Gaussian kernel size of 6 mm along all three directions.
The masks of the triple networks (DMN, SN, and ECN) were created by using an atlas of 90 functional regions of interest (Shirer et al., 2012). Only those voxels that showed more than 50% probability of being gray matter (GM) were considered (GM correction) and co-registered to a tissue-segmented MNI152 (2 × 2 × 2 mm3) template. The voxel values within the triple network masks were extracted for each subject, and a one-sample Kolmogorov–Smirnov test verified their normality. The mean values of ALFF, ReHo, and DC were calculated from the extracted voxel values for each subject.
Statistical analysis
The statistical analysis comprised several independent steps. Thereby, an alpha level of less than 0.05 was considered statistically significantly. All tests were two-tailed.
First, the age dependence of the neurocognitive parameters was examined by means of performing bivariate correlations of each parameter with age calculated as Pearson's (for variables with a normal distribution) or Spearman's correlation coefficient (for data without normality). The influence of age on the fMRI RS parameters of the three networks was examined by linear regression analysis. For a visualization of the age dependence of the fMRI parameters, scatter plots were created. In some cases, the associations seemed to follow a quadratic pattern, so an additional curvilinear regression analysis was carried out. Those parts of the statistical analysis were performed by using SPSS Statistics for Windows, Version 25.0 (IBM Corp.). As the results of the individual tests, as well as the fMRI parameters, are not entirely independent of each other, we used the Hochberg adjustment for multiple comparisons (HOCHBERG, 1988), which is considered to be more suitable than a Bonferroni correction in such constellations (Chen et al., 2017).
In addition, a partial correlation analysis was performed between the fMRI parameters and the neurocognitive variables by using a MATLAB software package [Version 9.5 (R2018b), Mathworks]. Due to the explorative approach of this examination, no correction for multiple testing was applied in these analyses, which are primarily intended to guide the way to further studies.
The comparison of the individual parameters between the three networks was carried out with the SPSS Software by using an analysis of variance (ANOVA). Only in case of a statistically significant effect obtained in the global F test, post hoc t tests were performed for multiple pairwise comparisons. To account for multiple testing, an alpha adjustment was carried out by using Tukey-Kramer correction.
Results
Neurocognitive performance
All results of the neurocognitive tests are given in Table 1. Following the Hochberg adjustment for multiple testing, almost all tests correlated significantly negative with age, except the following variables: TAP-visual scanning-omissions (p = 0.706), TAP-Alertness-index of phasic alertness (p = 0.465), TAP-Alertness-SD (p = 0.1), WCS-perseverative errors (p = 0.051), and TAP-divided attention-omissions (p = 0.025). The p-value for the sixth ranked test Digit Span-backward (p = 0.007) was lower than the corresponding adjusted Hochberg significance level (0.0083), so that other, smaller p-values could be considered as significant.
Group means (N = 74) in the Different Neurocognitive Test Parameters (Row Values) As Well As Their Correlations with Age
Column 1 contains the mean values of the corresponding parameters or in case of TAP battery the median values. Asterisks (**) mark correlations with age, which remained significant after Hochberg adjustment for multiple testing.
RT, reaction time; SD, standard deviation; TAP, Tests of Attentional Performance; TMT, Trail Making Test; WCST, Wisconsin Card Sorting Test; WMS, German version of the Wechsler Memory Scale—revised.
RS-fMRI measures—identification of the three core rsNWs
The three core rsNWs are depicted in Figure 1. Very concretely, DMN comprised the precuneus, the posterior cingulate cortex (PCC), the medial prefrontal cortex (mPFC), and the angular gyrus. The SN included the anterior cingulate cortex (ACC) and the frontal insular cortex (FIC). The subregions of the CEN were the dorsolateral prefrontal cortex (DLPFC) and the lateral posterior parietal cortex (LPPC). ALFF, ReHo, and DC measures across the whole brain are shown in Figure 2.
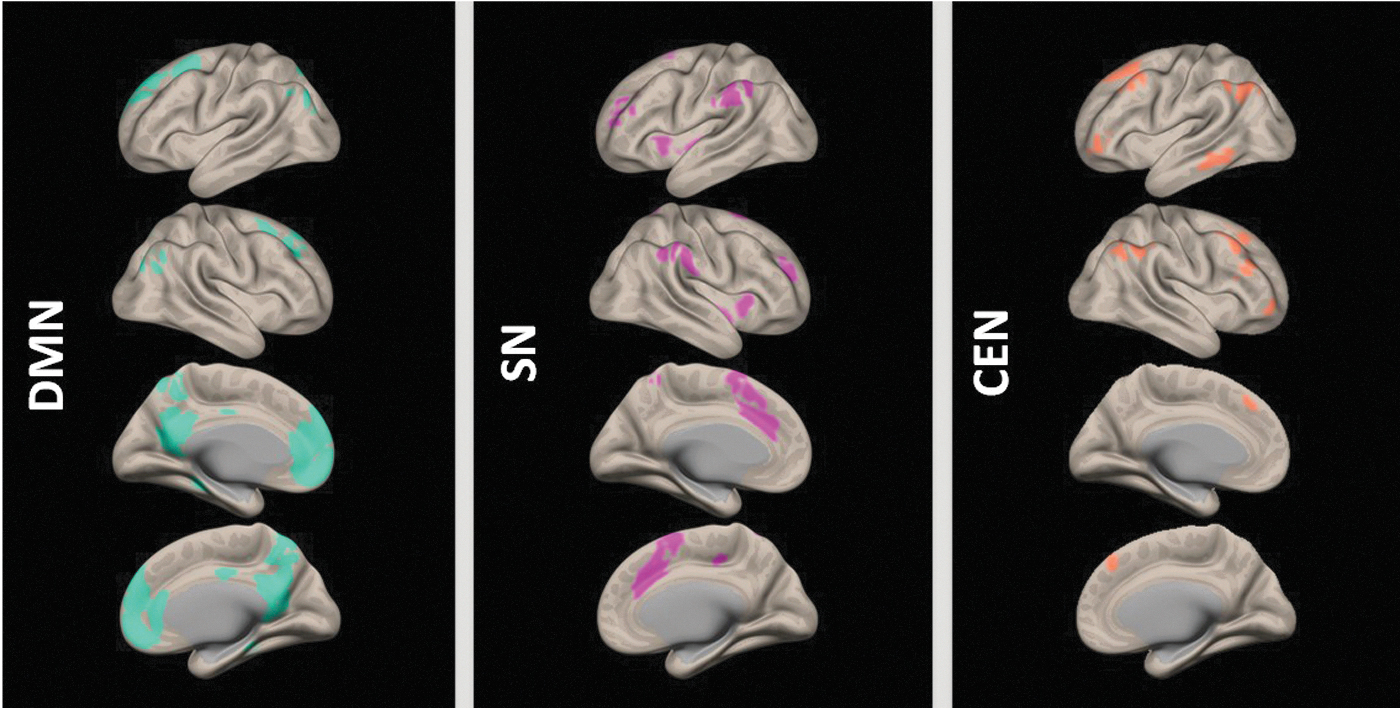
The three core resting-state networks (rsNW): DMN - default mode network, SN - salience network, and CEN - central executive network.

Resting-state measures across the whole brain: ALFF - amplitude of low-frequency fluctuations, ReHo - regional homogeneity, and DC - degree of network centrality.
RSA (ALFF) across the three core rsNWs
The ALFF values within the three core rsNWs were compared between the networks. The one-way ANOVA for three groups revealed a significant group difference in ALFF values between the networks [F(2, 219) = 14.363; p < 0.001]. The post hoc analysis revealed significantly lower values in the SN compared with DMN and CEN (both: p < 0.001) (Fig. 3).
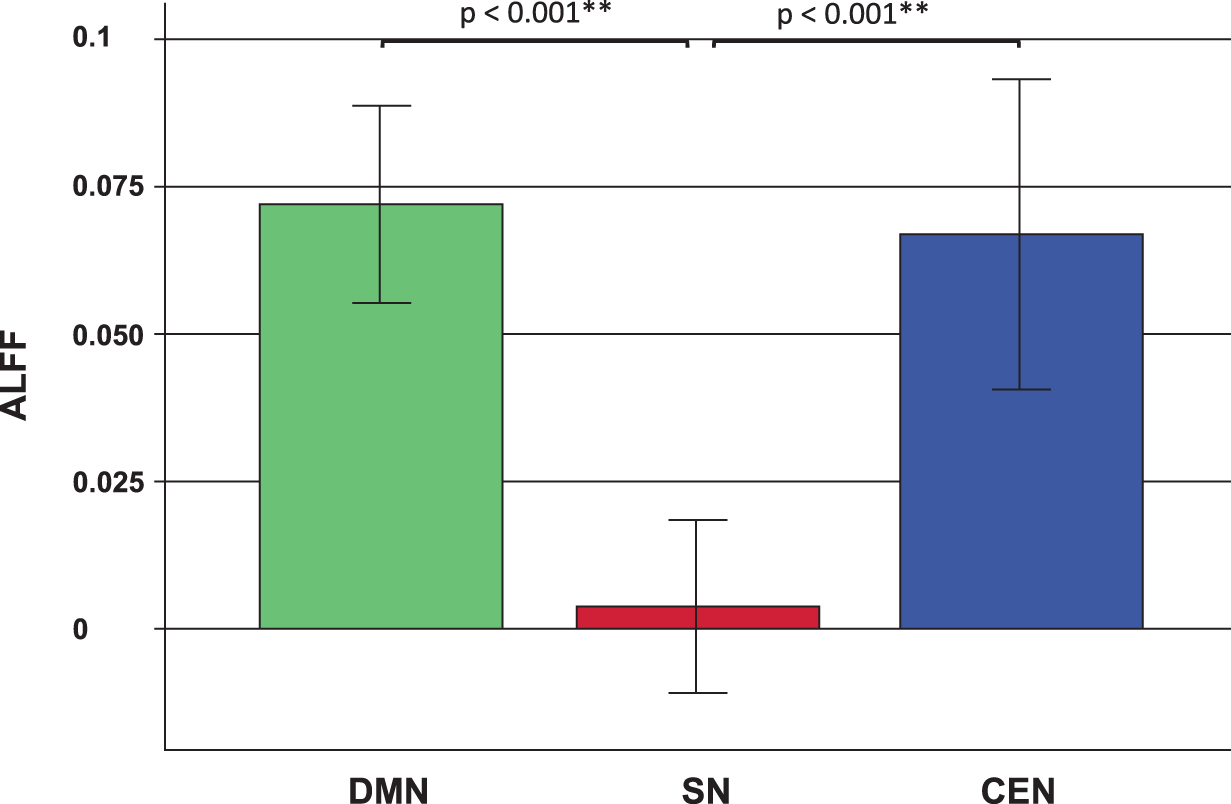
Amplitude of low frequency fluctuations (ALFF) values in the three core resting state networks (rsNW): Default mode network (DMN), the salience network (SN), and the central executive network (CEN). The error bars show the 95% confidence intervals. The calculated measures were linearly standardized into Z-values for each subject as described in the Materials and Methods section. The negative ALFF values within the SN arise from the circumstances that they are lower than the whole brain values. Asterisks (**) mark statistically significant differences (p < 0.01).
Local connectivity (ReHo) across the three core rsNWs
With respect to ReHo, the three networks differed significantly [F(2, 219) = 24.36; p < 0.001]. The post hoc analysis revealed significantly higher ReHo values in the DMN than in SN (p < 0.001) as well as in the CEN (p = 0.008). Further, the values in SN were significantly lower than in the CEN (p < 0.001) (Fig. 4).

Regional homogeneity (ReHo) within the default mode network (DMN), the salience network (SN), and the central executive network (CEN). The error bars show the 95% confidence intervals. Asterisks (**) mark statistically significant differences (p < 0.01).
Global connectivity (DC) across the three core rsNWs
Regarding DC, the one-way ANOVA also revealed a significant overall difference between the three groups [F(2,219) = 11.38; p < 0.001] (Fig. 5). In the post hoc analysis, DMN showed the highest DC values, differing significantly from SN (p < 0.001), but not from the CEN (p = 0.367). The DC values in the SN were significantly lower than in the CEN (0 = 0.006).

Degree of network centrality (DC) within the default mode network (DMN), the salience network (SN), and the central executive network (CEN). The error bars show the 95% confidence intervals. Asterisks (**) mark statistically significant differences (p < 0.01).
Influence of age on RSA and FC
The RSA and FC parameters mainly showed a decrease with increasing age in all three core rsNWs. Following the Hochberg adjustment for multiple testing, the following linear regressions remained significant: Within the DMN: ALFF (r = −0.624, r 2 = 0.390, **p < 0.001) and DC (r = −0.395, r 2 = 0.156; p < 0.001); in SN for ALFF (r = −0.397; r 2 = 0.158, **p < 0.001) and ReHo (r = −0.411, r 2 = 0.169, p < 0.001). None of the fMRI parameters in the CEN was significantly associated with age (Fig. 6).
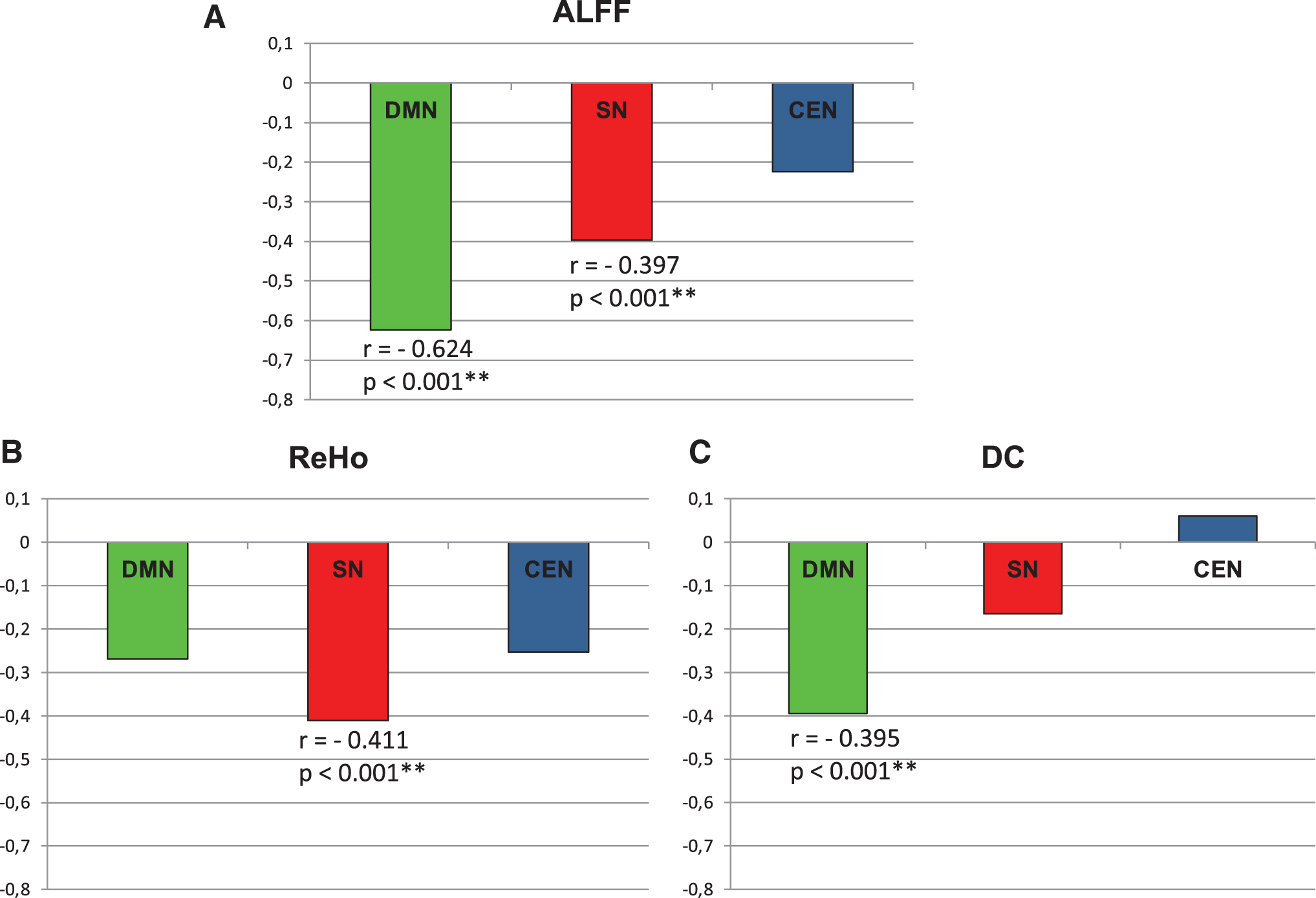
Age-related decrease of the values of ALFF
The curvilinear regression analysis has not revealed a significant r 2-value change for the age dependence of any of the fMRI parameters. A slight, though statistically nonsignificant, change of the r 2 value could only be observed for the ALFF value in CEN [linear regression: F(1,72) = 3.819; r = −0.224; r 2 = 0.05; p = 0.055; quadratic regression: F(1,71) = 0.753; r 2 = 0.096; p = 0.028; r 2-change = 0.045, significance of F-value change: p = 0.064] and for the ReHo value in CEN [linear regression: F(1,72) = 4.928; r = -0.253; r 2 = 0.064; p = 0.03; quadratic regression: F(1,71) = 3.961, r 2 = 0.1, p = 0.023, r 2-change = 0.036, significance of F-value change: p = 0.095]. Since both observed F-value changes were above the 5% significance level, these findings do not support the conclusion that there is a curvilinear relationship between the two measures and age. However, the observed trend might encourage looking for the possible occurrence of such associations in larger groups.
Details from the curvilinear regression analysis are shown in Supplementary Table S1, and the corresponding scatter plots for each parameter are shown in Supplementary Figures S1–S3.
Association between cognitive performance and RSA and FC
The intercorrelations between the different cognitive variables and the three rsfMRI measures in the three core rsNWs are shown in Figure 7. Due to the significant effects of age on neurocognitive performance, as well as on the fMRI metrics, a partial correlation (with age as a control variable) was calculated.
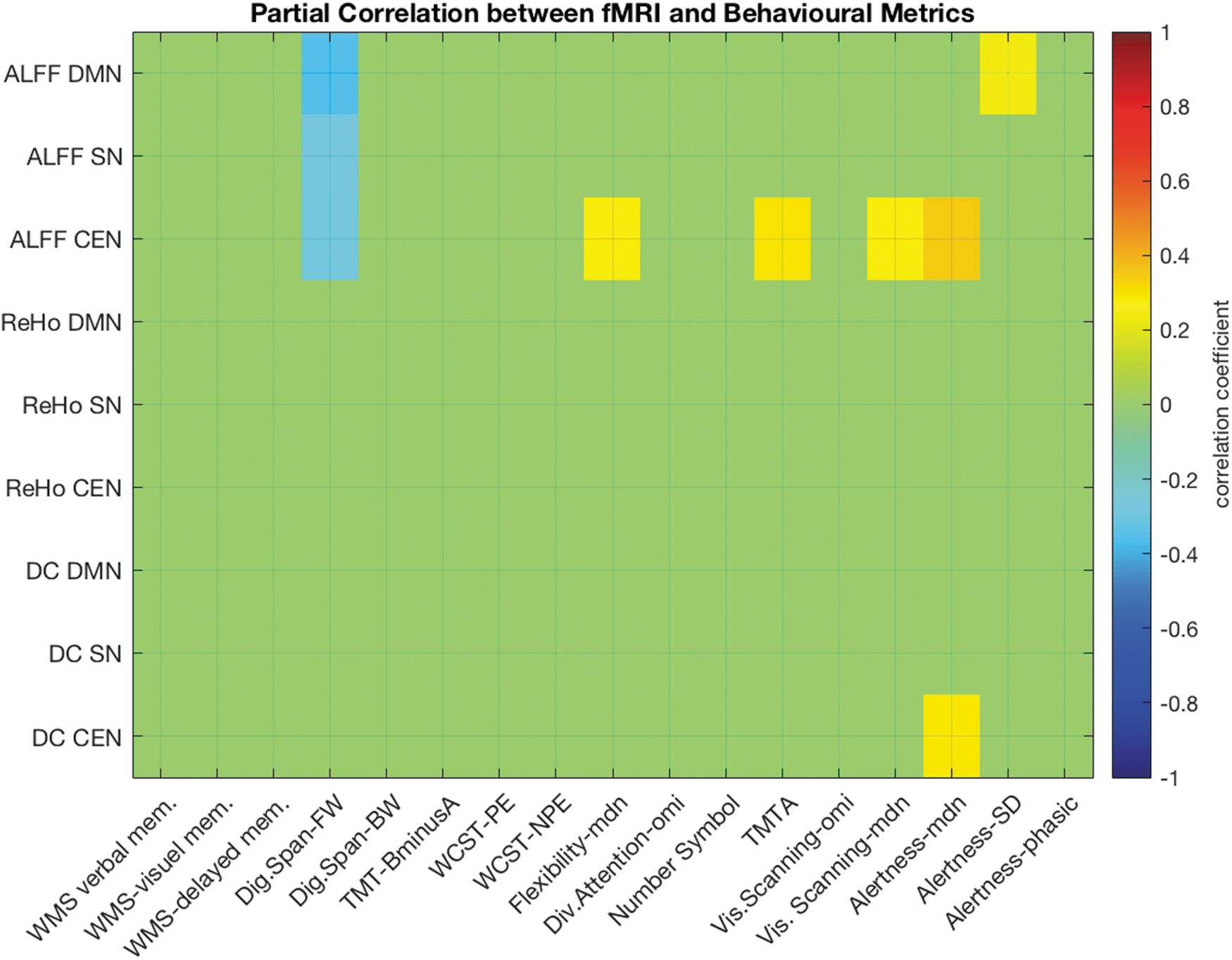
Partial correlation (corrected for age) between the ALFF, the ReHo, and degree of network centrality (DC) in the three core rsNW (DMN, SN, and central executive network [CEN]) and the results of the cognitive test-battery. Only significant (p < 0.01) correlation values are colored in the image matrix. BW, backwards; Dig.Span, digit Span; FW, forwards; NPE, nonpereseverative errors; mdn, median; omi, omissions; PE, perseverative errors; RT, reaction time; SD, standard deviation; TMT, Trail Making Test; WCST, Wisconsin Card Sorting Test; WMS, German version of the Wechsler Memory Scale—revised.
In the DNM, the ALFF correlated significantly positively with the SD of reaction times in the alertness test (alertness-SD, r = 0.24, p = 0.045), revealing more consistent simple reaction times (and thus less attentional fluctuation) in people with a lower level of RSA in the DMN. Further, a significant negative correlation between working memory (digit span) and the ALFF values in DMN (r = −0.353; p = 0.002) was observed.
In the SN, only a significant negative correlation between the ALFF values and working memory (r = −0.273, p = 0.02) was observed.
In the CEN, higher ALFF values were associated with lower working memory scores (r = −0.28; p = 0.016), lower attentional flexibility (r = 0.28; p = 0.017), lower performance in TMT-A (r = 0.29; p = 0.013), lower performance regarding visual scanning (r = 0.28; p = 0.016), and alertness (Alertness median r = 0.35; p = 0.002). Further, higher DC values in the CEN were also associated with lower alertness (r = 0.282, p = 0.016). All reported values include an age correction, but not a correction for multiple testing.
Discussion
The aim of the study was to explore the relevance of RSA and short- and long-range FC of the three core rsNWs for cognition in healthy individuals over a long age span. Previous investigations suggest that aberrant internal organization within the individual functional networks or disrupted interconnectivity between them is a characteristic of many psychiatric and neurological disorders, including schizophrenia, depression, dementia, and autism (Wu et al., 2016). Thus, a deeper understanding of what constitutes healthy patterns of network properties, as well as the age-associated changes in these three networks and their effects on cognition, is of high scientific interest.
Age dependence of cognitive performance
In accordance with the well-known cognitive decline experienced during normal aging, even in the absence of disease (Nashiro et al., 2017), the performance in almost all of the neurocognitive tests conducted in this study decreased significantly with age, with only a few exceptions. However, in contrast to previous robust findings showing that age has a significant influence on executive functions (Rhodes, 2004), we did not find age-related changes for the number of perseverative errors in the WCST. It is important though to keep in mind the relevance of education for the WCST (Boone et al., 1993), as this result may be a consequence of the high educational level of the participants in our sample.
Further, fluctuation of alertness and phasic alertness were also found to be age-independent in our study. This is in line with some earlier findings indicating the relative stability of attention (Ortiz et al., 2014; Staub et al., 2014). Moreover, we could not observe an age-associated increase in omission rates in either the divided attention task or the visual scanning test. These findings confirm the previously reported preservation of some aspects of attention (Thomson and Hasher, 2017) and visual scanning accuracy (Madden et al., 1999) over the lifespan.
Patterns of the rsfMRI metrics in healthy volunteers
The fMRI part of our study addressed the RSA and FC within the three core rsNWs. In distinction from many previous publications, which primarily investigated the properties of individual brain subregions to draw conclusions about the networks as a whole, our study followed a network-based approach in determining the RSA and FC parameters for the complete networks. In this approach, the temporal dynamics of the intrinsic brain networks is more robust compared with the consideration of singular voxels or regions. This is due to a lower susceptibility to noises caused by motion artefacts, vascular pulsatility, and respiration (Qi et al., 2018; Zuo et al., 2010). Further, the whole-network approach enables a statement to be made on the overall function of the networks and not merely on the function of individual subregions; most of these cannot be exclusively assigned to (just) one network. Thus, the comparison of acquired fMRI measures between the three networks revealed a pattern, with the highest values in the DMN and lowest values in the SN, for all three observed parameters.
Our finding reveals that the highest spontaneous RSA in the DMN during rest corresponds with established knowledge about DMN as a task-negative network that is mainly active at rest and typically deactivated during most stimulus-driven cognitive tasks (Greicius et al., 2003; Raichle, 2015). Our results also show not only the highest levels of RSA in DMN, but also the highest local connectivity (ReHo) compared with the other core rsNWs. This confirms previously reported highest ReHo values in the DMN (Tomasi and Volkow, 2011). ReHo reflects a widely used, most efficient, and reliable index of local FC (Zuo et al., 2013; Zuo and Xing, 2014), which is defined by the temporal coherence or synchronization of the BOLD time series within a set of a given voxel's nearest neighbors (Jiang and Zuo, 2016). An increase in ReHo indicates an increased local synchronization of spontaneous neural activity (Tommasin et al., 2017). Further, ReHo correlates with measures of functional segregation, such as local efficiency and clustering coefficients (Lee and Xue, 2017). Thus, high ReHo in the DMN may reflect the high levels of functional segregation (Respino et al., 2020) within the DMN during rest. Further, high signal coherence within the network ensures a tight functional coupling of the sub-components of the network, possibly resulting in more distinct functional specialization (Dennis and Thompson, 2014; Sternberg, 2011; Wig et al., 2011).
Interestingly, structures belonging to the DMN also showed significantly higher levels of global FC (DC) when compared with the SN and the CEN. As a graph-based measure of network organization, DC mirrors the number of instantaneous functional connections between a region and the reminder of the brain, within the entire connectome (Takeuchi et al., 2015). Thus, it shows how much of the node influences the entire brain areas and is involved in integrating information across functionally segregated brain regions (Hagmann et al., 2008). High levels of DC in the DMN suggest that the DMN has a strong influence on the other brain regions during the resting state.
We observed the lowest values for all three fMRI measures (ALFF; ReHo, DC) in the SN. Initially, the function of the SN was proposed to mainly include the identification of the homeostatically most relevant stimuli from the multiple internal and external stimuli that constantly accost the brain (Seeley et al., 2007). Any event that reorients the individual and redirects his/her attention is salient, whether it affects the individual physically (as in pain) or psychologically (as with empathy). In addition, the SN (particularly the anterior insula) has been found to mediate “dynamic switching” between the DMN and the CEN, facilitating the focusing of attention on external stimuli, which adds significance or saliency to them (Menon and Uddin, 2010). The neurobiological correlates of several brain disorders have been tied to the SN (Uddin, 2017). Among others, increased activity and connectivity in the SN may underlie not only the hypervigilance and hyperarousal symptoms commonly observed in patients with PTSD (Fenster et al., 2018) but also symptoms of depression and anxiety (Hamilton et al., 2012; Peterson et al., 2014). Our results indicate that in the absence of salient signals, the spontaneous activity of the SN, considered as a whole, as well as its local and global connectivity, are significantly below the DMN and CEN. This could be a pattern of a physiological balance between the three networks in a state where the SN is neither processing relevant stimuli nor regulating the activity of other networks.
Age effects on the fMRI metrics
The examination of age effects on the fMRI metrics in our study revealed significant findings for the DMN and SN, but not for the CEN. When interpreting our results, the specifics of our methodology have to be taken into account.
Our results confirm the previously reported finding of age-related decrease in spontaneous brain activity (ALFF) in the DMN and SN (Hu et al., 2014; Lopez-Larson et al., 2011). Interestingly, however, we did not observe a significant age-related ReHo decrease in the DMN, which is divergent from a previous consistently reported decrease of connectivity in the DNM (Ferreira and Busatto, 2013), but may be attributable to our network-based approach. Although in previous studies, the observed decrease in ReHo values for the DMN was derived from data from individual subregions of the DMN, our results are based on the joint consideration of all subregions that were identified as part of the DMN.
Conversely, in our study, the DMN in its entirety was the only network where the mean number of connections per voxel (DC) decreased significantly with increasing age during rest. This finding indicates an age-dependent decrease of the influence of the DNM on the entire brain and its decreasing involvement in integrating information across functionally segregated brain regions (Hagmann et al., 2008). This result deviates somewhat from a number of findings, indicating a generally increased FC between networks in elderly subjects (Chan et al., 2014; Geerligs et al., 2015; Song et al., 2014). However, none of the aforementioned studies used the same approach as applied here. Further, a recent large population-based study (Zonneveld et al., 2019) reported variable patterns of both increases and decreases of (anti-)correlations in approximately equal proportions between different brain networks. In this study, the strongest age-dependent connectivity decrease was related to the DMN, as also seen in our study.
In the SN, we observed a significant age-dependent decrease for ALFF and ReHo but not for DC. To date, studies of age-related changes in the SN have shown mixed results, including decreased age-related connectivity (He et al., 2014; Langner et al., 2015; Roski et al., 2013), but also preserved or increased connectivity in other cases (Sakaki et al., 2016; Xiao et al., 2018). However, a closer look shows that the focus in these previous studies was placed on different structures of the SN, which are known to have distinct atrophy rates due to the cytoarchitectonic properties of each particular subregion (McGinnis et al., 2011). Indeed, in their recent work, Touroutoglou and colleagues (2018) found that connectivity within the ventral subsystems of the SN remained stable (preserved or even increased with age), whereas the connectivity within the dorsal subsystem decreased with age. Examining the age effects on the whole SN, we could demonstrate an age-dependent decrease of the spontaneous RSA and ReHo. This is in line with the report by Lopez-Larson and associates (2011), who observed the main ReHo decrease in the ACC and the temporal lobe, which also make up the SN. In support of this, the highest correlation of the ReHo values with age in the SN indicates that the temporal synchronization of the nearest neighbor nodes (Jiang and Zuo, 2016) decreases mostly pronounced within the SN during the lifespan.
Conversely, we observed stable long-range connectivity in the SN during aging. This is in agreement with the fact that SN forms an anatomically tightly coupled network, ideally placed to integrate information from several brain regions (Critchley et al., 2004). Thus, the stability or even strengthening of long-range network connectivity concurrent with a parallel weakening of short-range FC with increasing age (Uddin et al., 2010) appears to be particularly important in the SN. This is likely due to its unique wide range of functions in integrating information and coordinating the activity of other networks (Uddin, 2017).
None of the fMRI parameters in the CEN showed significant age dependency. This is in agreement with some previous works that reported an absence of age-dependent connectivity decrease for the regions summarized here as the CEN (Onoda et al., 2012). However, visual inspection of the scatter plots has raised a question about the possible curvilinear age dependence of the parameters ALFF and ReHo in CEN. An additional curvilinear regression analysis revealed a slight although not statistically significant increase in r 2-values as a goodness-of-fit measure in a quadratic model. Thus, a curvilinear relationship with age would explain 9.6% of the variability in ALFF values (4.5% increase over the linear model) and 10% (3.6% increase over the linear model) of the variability in ReHo values in CEN. As this is only a statistical trend (in terms of the significance of the changes of r 2), these results do not allow any conclusive statements to be made, but they do indicate a possible relationship that should be investigated in further studies.
Age-independent associations between the rsfMRI metrics and cognition
As both the fMRI metrics and cognitive performance generally decrease significantly with age, we investigated partial correlations between the cognitive scores and the fMRI metrics, accounting for the effects of age on both variable sets. This approach revealed several significant associations.
Higher spontaneous RSA in the DMN was related to higher alertness variability and lower working memory. Defined as a task-negative network, DMN is associated with mind-wandering and self-referential thoughts (Lin et al., 2017), and previous investigations show that the DMN has an important role in spontaneous cognition, daydreaming, or stimulus-independent thoughts (Andrews-Hanna et al., 2010). Studies have also demonstrated its role in the support of emotional processing, self-referential mental activity, and the recollection of prior experiences (Raichle, 2015).
The majority of investigations suggest that cognitive performance is positively correlated with DMN FC (Mevel et al., 2013; Persson et al., 2014; Sambataro et al., 2010; Yang et al., 2013). In addition, a recent meta-analysis supports the view that changes in FC and DMN activation might be explained by a combination of age effects and cognitive abilities (Mak et al., 2017). However, despite numerous previous investigations, the cognitive role of the DMN is still not sufficiently understood. Indeed, it is still unclear whether the (co)activity of the DMN nodes reflects the participant's mental content/inner experience during the scan, or rather represents the base functioning of the brain, independently from the inner experience during the scan but potentially related to cognitive abilities (Mevel et al., 2013).
In this study, the properties of the DMN were investigated independently from the assessment of cognitive performance. After controlling for age, we observed that higher spontaneous activity within the DMN predicts lower working memory and lower attentional stability. In contrast to our findings, Gui and associates (2015) found a more stable performance during a sustained attention test in subjects with higher pre-test resting baseline ALFF values in two core DMN regions (the PCC and the MePFC). However, here we report the ALFF values for the whole DMN and not only for selected regions of maximum activity. Moreover, contrary to the cited work, we found associations after adjustment for the age effects, revealing a negative influence of higher DMN activity on alertness stability and working memory.
A previous investigation suggests that increased connectivity of the DMN could be considered to be a neural substrate of depressive rumination (Hamilton et al., 2015). Thus, the higher level of self-referential processes during rest seems to be predictive for lower attention and working memory.
In the SN, we only observed a significant negative correlation between the ALFF values and the working memory. The SN plays a pivotal role in determining the biological significance of external stimuli (Seeley et al., 2007). In contrast to the DMN, which mainly assigns valence to internally represented stimuli, the main function of the SN is to complement the function of the DMN by assessing the significance of external stimuli (Hamilton et al., 2015). The SN is responsible for the integration of emotional, visceromotor, autonomic, and interoceptive responses (Menon, 2011) and seems to mediate emotional, somatic, and vegetative aspects of several psychiatric disorders (Groenewold et al., 2013). Indeed, increased activity in the SN, in particular of the anterior insula, has been reported in relation to anxiety, depression, and insomnia (Liu et al., 2015, 2018). Our results extend this knowledge by showing a correlation with a poorer working memory performance.
In terms of the CEN, we observed a significant negative association between spontaneous RSA and performance in several cognitive domains. In detail, higher RSA in the CEN was associated with lower performance in working memory, TMT-A, visual scanning, alertness, and also lower attentional flexibility. CEN is a task-positive network that is engaged in higher-order cognitive and attention control as well as in decision making, working memory, and goal-directed behavior (Bressler and Menon, 2010; Knight, 2006). In addition to its direct involvement in cognitive processes, the CEN also supports the cognitive regulation of behavior, emotion, and thought. It is activated during efforts to exert self-control, suppress intrusive, unpleasant thoughts, and reappraise threatening stimuli (Gagnepain et al., 2017; Turner et al., 2019). Our results indicate that higher involvement of the CEN in such activities during rest results in lower performance in several cognitive domains.
Conclusion
Our investigation of network properties within the three core rsNWs revealed a pattern with low RSA and FC in the SN in a sample of healthy volunteers aged 20 to 79 years. In terms of aging, both RSA and FC remained stable in the CEN, while decreasing with age in the DMN and SN.
Further, higher RSA, particularly in the central executive network, seems to have had negative effects on cognition in our sample. This relationship was shown here, following correction for age-related changes, which are known to be important for both network properties and cognition. These findings present a new perspective, particularly when seen against the backdrop of the somewhat conflicting findings regarding the usefulness of RS studies for understanding the neurobiological basis of cognition (Campbell and Schacter, 2017; Davis et al., 2017; Grady, 2017). It is anticipated that the whole-network approach applied here, with a focus on the large rsNWs and a more precise characterization of their resting-state properties, could provide valuable new insights into the effects of age on the neurobiological correlates of cognition.
Footnotes
Acknowledgments
The authors would like to thank the Dr. Jorge Arrubla for his help in data acquisition, Petra Engels for technical assistance and support during the scans, and Claire Rick for proofreading. This study is deemed to be part of the doctoral thesis (Dr. med.) of Laura Amort, Faculty of Medicine, RWTH Aachen University, Germany.
Authors' Contributions
T.V:. conception of the statistical design and analysis, data analysis, interpretation of the results, article writing (original draft) and revision, and critical revision of the work for important intellectual content. R.R.: analysis of the fMRI data and conception of the methodology, creation of codes for the analysis, revision of the article, and critical revision of the work for important intellectual content. L.A. and J.J.: data acquisition and analysis, revision of the article, and critical revision of the work for important intellectual content. N.J.S.: funding acquisition, supervision, provision of resources, revision of the article, and critical revision of the work for important intellectual content. B.F.: compilation of the cognitive test battery, conception of the statistical analysis, revision of the article, supervision, and critical revision of the work for important intellectual content. I.N.: funding acquisition, study design, supervision, data acquisition, article writing and revision, and critical revision of the work for important intellectual content.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.