
Abstract
Introduction:
Even with the increased access and early initiation of combination antiretroviral therapy, children with perinatally acquired human immunodeficiency virus (CPHIV) continue to demonstrate white matter alterations. Children perinatally HIV-exposed, but uninfected (CHEU) alike show differences in white matter integrity compared with children who are HIV-unexposed and uninfected (CHUU).
Objectives:
Mapping white matter connections that link gray matter regions that form resting-state (RS) functional networks may demonstrate whether structural and functional connectivity alterations in HIV infection and exposure may be related. We hypothesized reduced structural connectivity in CPHIV within the default mode network (DMN), visual, ventral DMN (vDMN), somatosensory, salience, auditory, motor, executive, basal ganglia, and posterior DMN (pDMN). We also hypothesized that CHEU will have increased structural connectivity compared with CHUU in the vDMN, somatosensory, pDMN, dorsal attention, salience, auditory, motor and basal ganglia.
Methods:
Study participants were 61 seven-year-old CPHIV and 46 age-matched children who are HIV uninfected (CHU) (19 CHEU). We used diffusion tensor imaging-based tractography to investigate white matter connections that link gray matter regions within RS functional networks.
Results:
We found altered white matter integrity in the somatosensory, salience, default mode, and motor networks of CPHIV compared with CHU. The superior temporal cortex, superior frontal cortex, and putamen were affected in all four networks and have also been reported to demonstrate morphological alterations in the same cohort. In CHEU, white matter integrity was higher in the visual network, pDMN, and motor network compared with CHUU.
Conclusion:
Our results suggest that altered white matter integrity may influence gray matter morphology and functional network alterations.
Impact statement
The long-term effects of human immunodeficiency virus (HIV) and exposure on the developing brain in the combination antiretroviral therapy era are still not well known. We use diffusion tensor imaging-based tractography to explore these effects on white matter connections that link gray matter regions within functional networks. Our findings provide a context for HIV-associated white matter and connectivity abnormalities.
Introduction
Over 3% of the South African population living with the human immunodeficiency virus (HIV) are children (StatsSA, 2019). Despite increased access to and early initiation of combination antiretroviral therapy (cART), children with perinatally acquired HIV (CPHIV) continue to demonstrate neurologic abnormalities as well as neurocognitive delays and deficits (Boivin et al., 2019; van Arnhem et al., 2013). Further, 20% of children perinatally HIV-exposed, but uninfected (CHEU) are living in South Africa (UNAIDS, 2019) where one in every four children is born to a mother living with HIV (Slogrove et al., 2019). This makes it the country with the highest HIV statistics worldwide. The long-term effects of HIV infection and exposure on the developing child in the era of early treatment are not well known.
Diffusion tensor imaging (DTI) measures microstructural properties of white matter (O'Donnell and Westin, 2011). The DTI outcomes include fractional anisotropy (FA), an indicator of myelin and axonal integrity, mean diffusivity (MD), an indicator of white matter maturation, axial diffusivity (AD), a measure linked to axonal properties, and radial diffusivity (RD), a parameter related to both aspects of myelination and axonal health.
The Children with HIV Early Antiretroviral therapy (CHER) trial examined the clinical outcomes of early treatment regimens in infants (Cotton et al., 2014; Violari et al., 2008). An ongoing longitudinal follow-up study, using neuroimaging and neurocognitive measures, is focused on neurodevelopment in the Cape Town cohort. A voxelwise-DTI study of this cohort at age 5 found lower FA/higher MD in various white matter clusters in CPHIV compared with age and socioeconomic-matched children who are HIV-uninfected (CHU) (Ackermann et al., 2016). At age 7, we found differences in white matter integrity in similar regions as age 5. In addition, we found MD increases in bilateral clusters in the superior corona radiata (SCR) and anterior thalamic radiation (ATR) that were not observed at age 5, suggesting persistent and new damage despite early ART initiation in this cohort (Jankiewicz et al., 2017).
Within the literature, DTI studies on the effects of HIV/cART indicate that although ART provides some protection to the developing brain, white matter abnormalities persist. These studies report white matter alterations, either lower FA and/or higher MD in CPHIV, across similar regions to those we report at 5 and/or 7 in our cohort. Our study as well as others report clusters within the following tracts: corticospinal tract (CST), SCR, inferior fronto-occipital fasciculus (IFOF), inferior longitudinal fasciculus (ILF), cingulum, superior longitudinal fasciculus (SLF), ATR, forceps minor, and uncinate fasciculus (Ackermann et al., 2016; Hoare et al., 2015a; Jankiewicz et al., 2017; Li et al., 2015; Uban et al., 2016). Although the aforementioned studies report regional decreases in FA in CPHIV, a recent study showed regional FA increases in ART treated youth with PHIV (17.9 ± 2.5 years) compared with age-matched uninfected youths (Sarma et al., 2019). The FA reductions and MD increases are largely attributable to greater RD, suggesting an effect of HIV/ART on myelination (Ackermann et al., 2016; Hoare et al., 2012, 2015a; Jankiewicz et al., 2017).
Lower FA and/or higher MD has also been negatively and positively associated with neuropsychological performance (Ackermann et al., 2020; Hoare et al., 2012; Jahanshad et al., 2012; Tran et al., 2016; Yadav et al., 2020). Further, children who began treatment later than 2 years of age demonstrated lower FA, and higher MD and RD in white matter tracts, including the SCR (Hoare et al., 2019). Another study found that ART provided localized white matter protection in CPHIV compared with ART-naive CPHIV (Hoare et al., 2015b). FA was negatively associated with viral load (VL) and positively associated with nadir CD4 Z score in ART-treated youth (Cohen et al., 2016). In summary, these papers link localized white matter alterations to disease severity and neuropsychological performance.
The DTI studies in CHEU populations are limited, and the results are mixed. Our 7-year-old voxelwise-DTI study found increased mean FA in the posterior corona radiata (PCR) and lower mean MD in the CST in CHEU compared with children who are HIV-unexposed and uninfected (CHUU) (Jankiewicz et al., 2017). Three other studies have used DTI in CHEU populations, with one finding no exposure-related differences (Jahanshad et al., 2016), one finding increased FA (Tran et al., 2016), and one reporting decreased FA (Yadav et al., 2020).
The DTI-based tractography, which requires gray matter regions as seeds, incorporates structural information. Therefore, the anatomical restrictions applied by tractography lead to fewer outcomes (e.g., white matter tracts), and a reduced statistical penalty from multiple comparison testing. An additional advantage of tractography is the flexibility to choose seeds based on a specific question or hypothesis. The present study used DTI-tractography at 7 years to examine the influence of HIV and ART exposure. Although we previously published voxelwise-DTI analysis at 7 years, it was challenging to interpret the results in terms of function. In this work, we used tractography to identify tracts within resting state networks (RSNs) to contextualize HIV-related differences in terms of well-defined functional networks. We previously published resting-state (RS)-fMRI findings from a subset of this cohort at 7 years, in which we used independent component analysis (ICA) to define 12 common RSNs.
We used our previously published voxelwise-DTI and RS-fMRI (Toich et al., 2018) work at age 7 years for hypotheses generation. The voxelwise-DTI paper included HIV and exposure-related comparisons, whereas the RS-fMRI paper presented only HIV-related comparisons in a subset of participants at 7 years. We hypothesized that, based on our RS-fMRI results at age 7, CPHIV will demonstrate decreased mean structural connectivity (low FA, high MD) in white matter tracts within the default mode network (DMN) as compared with CHU. Based on our voxelwise-DTI results, we posit decreased mean structural connectivity in CPHIV within RSNs that involve the IFOF, ILF, SCR, ATR, and forceps minor tracts. These networks include: visual, ventral DMN (vDMN), somatosensory, salience, auditory, motor, executive, basal ganglia, and posterior DMN (pDMN). Further, based on our voxelwise-DTI results, we suggest that CHEU will have increased structural connectivity (high FA, low MD) compared with CHUU in RSNs where the PCR and CST are present. These include vDMN, somatosensory, pDMN, dorsal attention (dATT), salience, auditory, motor and basal ganglia. Finally, exploratory analyses examined correlations between altered white matter and neuropsychological outcomes as well as clinical health measures.
Materials and Methods
Participants
A total of 116 seven-year-old children participated in the current study. The 68 CPHIV were recruited from the Cape Town cohort of the CHER trial (Cotton et al., 2014; Violari et al., 2008), whereas the 48 age-matched CHU were recruited from a similar community part of an interlinking vaccine trial (Madhi et al., 2010). In the CHER trial, at age 6–12 weeks, infants were randomized into one of three early treatment groups; two arms involved early (before 12 weeks) time-limited ART and one deferred ART until CD4% <25% in the first year or CD4% <20% thereafter, or if clinical disease progression criteria presented (Cotton et al., 2014). For the present study, all CPHIV were on ART. The ART regimens for all treatment arms consist of ZDV+Lamivudine (3TC, Epivir)+Lopinavir+Ritonavir (LPV/r, Kaletra). The CHU group is further subdivided into exposed and unexposed children.
Image acquisition
Neuroimaging data were acquired at age 7 years according to protocols approved by the Human Research Ethics Committees of the Universities of Cape Town and Stellenbosch. Participants were scanned at the Cape Universities Brain Imaging Centre (CUBIC) adjacent to Tygerberg Hospital, using a 3T Siemens Allegra scanner (Siemens, Erlangen, Germany) with a single-channel receive head coil. Structural T1-weighted images were acquired by using a volumetric navigated multi-echo magnetization prepared rapid gradient echo sequence (voxel size = 1.3 × 1.0 × 1.0 mm3, field of view [FOV] = 224 × 224 × 144 mm3, repetition time [TR] = 2530 ms, inversion time = 1160 ms, echo time [TE] = 1.53/3.19/4.86/6.53 ms, flip angle = 7°), followed by two DTI acquisitions with opposite phase encoding (Anterior-Posterior, Posterior-Anterior; AP, PA) directions using a prospectively motion-corrected twice-refocused spin echo sequence with five reacquisitions (Alhamud et al., 2012). Acquisition parameters for diffusion were: TR/TE 10,100/86 ms, 72 slices, 2 × 2 × 2 mm3, FOV = 224 × 224 × 144 mm3, 30 noncollinear diffusion directions, b = 1000 s/mm2, and 4 nondiffusion-weighted (b0) acquisitions.
Image preprocessing
All data were preprocessed by using a series of in-house scripts in the software package Tolerably Obsessive Registration and Tensor Optimization Indolent Software Ensemble (TORTOISE) version 3.1 (Pierpaoli et al., 2009). For each participant DICOMs were converted to an NIFtI readable format for TORTOISE and T2w anatomical images were created from the T1w images that were acquired during scanning. Subsequently, all diffusion weighted images (DWIs) were visually inspected and distorted volumes were discarded. Subjects with at least 15 diffusion directions (out of the 30), including at least 1 b0 image (out of the 4), were retained for further analyses. TORTOISE's DIFFPREP and DRBUDDI functions were used to correct motion and eddy distortion and DWI distortion, respectively. To estimate diffusion tensors and DTI parameters (FA, MD, etc.), AFNI's fat_proc_dwi_to_dt function was used.
In this study, we were interested in investigating structural connectivity within the 12 RSNs previously identified in a subset of children from this cohort by using group ICA of RS-fMRI data acquired during the same scanning session (Toich et al., 2018). The RSNs included two visual networks (lingual and occipital lobe), DMN, ventral DMN, pDMN, somatosensory, dATT, salience, auditory, motor, executive control, and basal ganglia.
Each subject's DTI data and masks of the 12 RSNs were warped onto the Haskins pediatric brain atlas (Molfese et al., 2011) by using AFNI's 3dNwarpApply (AFNI, 2014). We identified all the anatomical gray matter regions involved in each RSN and then applied the mask of the relevant RSN to the masks of the anatomical regions identified for that network to ensure that only the portions of the anatomical regions involved in that functional network were retained as seeds for tractography. Thus, even if a gray matter region belongs to multiple networks, only the relevant part of it is considered with regards to connections to other gray matter regions within a particular network.
Tractography
Tractography was performed by using the Functional and Tractographic Connectivity Analysis Toolbox (FATCAT) (version 1.1) (Taylor and Saad, 2013) in AFNI. The 3dTrackID function was used to perform probabilistic tractography (Taylor et al., 2016). Within each functional network, we performed tractography to identify white matter tracts that structurally connect pairs of gray matter regions. Each functional network, therefore, had a unique number of possible white matter connections as determined by the number of gray matter to gray matter connections within that network. For tracking we employed 3dTrackID's full probabilistic tracking mode. We used the following algorithm options: FA threshold = 0.2, angle threshold = 60, length threshold = 20 mm, number of seeds per voxel per Monte Carlo iteration = 5, number of Monte Carlo iterations = 1000. Further, for a white matter connection passing through a voxel to be included in the final white matter region of that connection, at least five tracks would have had to pass through the voxel.
Statistical analyses and visualization
For each subject and within each RSN, we extracted mean FA, AD, MD, and RD values in FATCAT for each tract connecting distinct gray matter regions. Using the statistical software R (version 3.4.2), we ran linear regression models, with sex as a possible confounder. The first model examined the possible influence of HIV infection (CPHIV vs. CHU). The second model looked at the effects of HIV exposure (CHEU vs. CHUU children). We used an alpha (α) significance level of p = 0.01 and corrected for multiple comparisons by using the false discovery rate method (p = 0.05) (Benjamini and Hochberg, 1995). The resultant connections were labeled via manual identification in FSLView (version 5.0.9), guided by JHU ICBM-DTI-81 White Matter Labels as well as the JHU White Matter Tractography Atlas (Hua et al., 2008; Mori et al., 2005; Wakana et al., 2007). White matter connections that could not be identified are listed as “WM connection.” For visualization of affected white matter connections (affected in FA and/or MD) and their corresponding gray matter regions, we extracted the gray matter regions and the white matter connection. The 3D volume representations of the gray matter regions and tracts that run between them were generated for a randomly selected control subject by using diffusion spectrum imaging Studio. Further, we present results in the form of graphs (created by using iGraph package in R) representing implicated network and white matter connections. Finally, we used linear regression models to investigate the possible relationships between altered DTI parameters and neuropsychological tests. The neuropsychological assessments were performed at the same age and included the Kaufman Assessment Battery for Children (KABC) and the Test of Variable Attention (TOVA). Our model included sex as a confounder, and interaction terms between DTI parameters and HIV status. We used an alpha (α) significance level of p = 0.05.
Results
Due to motion artefacts present in more than half of the DWI volumes, nine participants (seven CPHIV, two CHU) were excluded. On average, participants had 27 ± 3 DWIs and 3 ± 1 b0s and there were no significant group differences between groups. We present data from a total 61 CPHIV and 46 CHU 7-year-old children (Table 1). The CHU group is further subdivided into 19 CHEU and 27 CHUU (Table 2).
Sample Characteristics
Values are mean ± SD.
Bold p-values are statistically significant (p-value < 0.05).
Baseline CD8 data missing for one subject.
Twelve subjects were not interrupted and received ART continuously.
ART, antiretroviral therapy; CHU, children who are HIV uninfected; CPHIV, children with perinatally acquired HIV; CHU: children who are HIV uninfected; KABC-II, Kaufman Assessment Battery for Children; SD, standard deviation; TOVA, Test of Variables of Attention; VL, viral load.
Sample Characteristics for the Children Who Are Human Immunodeficiency Virus Uninfected
Values are mean ± SD.
CHEU, children perinatally HIV-exposed but uninfected; CHUU, children who are HIV-unexposed and uninfected.
The CPHIV showed lower FA and/or higher MD (Figs. 1 –3) and RD compared with CHU for various connections within the CST, SLF, forceps minor, cingulum, and ATR in the somatosensory, salience, DMN, and motor networks. In the CHEU versus CHUU group comparison, we found higher FA in CHEU compared with CHUU for connections within the cingulum in the pDMN, the forceps major in the visual (occipital lobe) network, and the SLF in the motor network (Figs. 4 and 5). We present the names of connected gray matter regions, affected white matter tracts, and relevant statistics in Tables 3 –5.

Networks with WM connections between GM regions showing significantly lower FA and/or higher MD in CPHIV than CHU. Right hemisphere regions are light blue, whereas left hemisphere regions are coral. The blue connections represent WM connections that showed lower FA, whereas the red ones represent connections that showed higher MD. The gray connections represent the rest of the WM connections that were present in all subjects and unaffected by HIV status. Somatosensory network: FA was lower in CPHIV compared with CHU in the WM connection between L putamen (L_Pu) and L superior temporal cortex (L_STC), CST between L paracentral gyrus (L_ParCG) and postcentral gyrus (L_PostCG), SLF between L_PostCG and L precentral gyrus (L_PreCG), WM Connection between L_STC and L insula (L_Ins), WM connection between R_Pu and R_STC, and CST between R_Pu and supramarginal cortex (R_SPM). The MD was higher in CPHIV compared with CHU in: Cingulum between R superior frontal cortex (R_SFC) and R posterior cingulate cortex (R_PCC). Salience network: The FA was lower in CPHIV compared with CHU in the Forceps Minor between R lateral orbitofrontal cortex (R_lOFC) and R rostral anterior cingulate cortex (R_rACC). The MD was higher in CPHIV compared with CHU in: ATR between L Caudate (L_Cd) and L_Pu, ATR between L_Cd and L lateral orbitofrontal cortex (L_lOFC), ATR between L_Pu and L_lOFC, ATR between L_Pu and L parstriangularis (L_Pars), Cingulum between L caudal anterior cingulate cortex (L_cACC) and L_PCC, SLF between L parsopercularis (L_Parso) and L_Ins, ATR between R_Pu and R_Parso, Forceps minor between L_SFC and R_cACC, Cingulum between L_PCC and R_cACC, and Cingulum between R_cACC and R_PCC. Motor: The FA was lower in CPHIV compared with CHU in the Forceps minor between L_Pu and L_STC. DMN: The MD was higher in CPHIV compared with CHU in: Forceps minor between L_rACC and L_SFC, Forceps minor between L medial orbitofrontal cortex (L_mOFC) and L_SFC, WM connection between L caudal middle frontal cortex (L_cMFC) and L_SFC, and Forceps minor between L rostral middle frontal cortex (L_rMFC) and L_SFC. ATR, anterior thalamic radiation; CST, corticospinal tract; CPHIV, children with perinatally acquired HIV; CHU, children who are HIV uninfected; DMN, default mode network; FA, fractional anisotropy; GM, gray matter; HIV, human immunodeficiency virus; L, left; MD, mean diffusivity; R, right; SLF, superior longitudinal fasciculus; WM, white matter. Color images are available online.
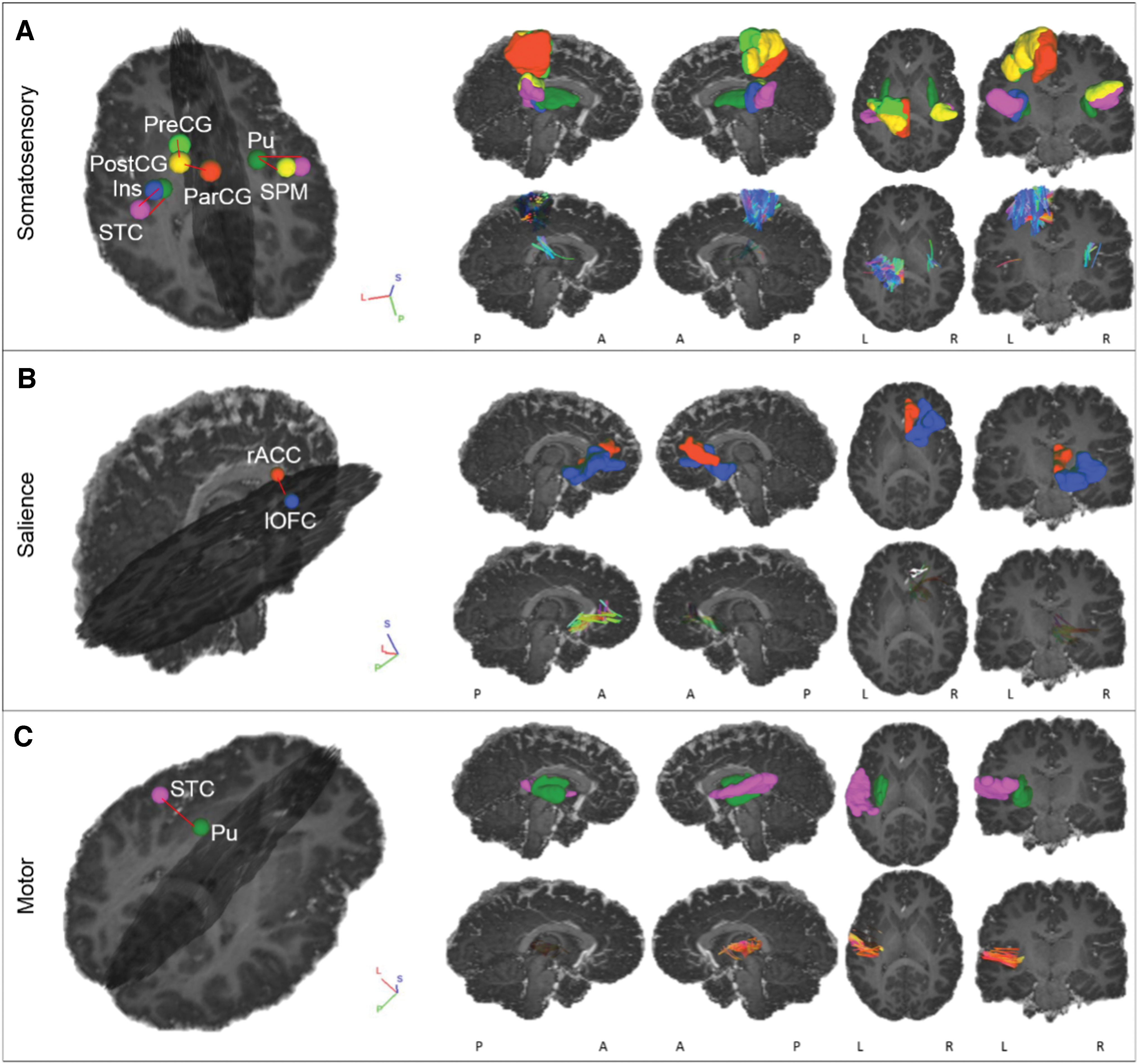
Networks with WM connections between GM regions showing significantly lower FA in CPHIV than CHU. The 3D volume representations were created in DSI studio. From left to right, each column shows: an oblique viewing angle of the GM regions and connections, right and left sagittal views, and axial and coronal views. In rows
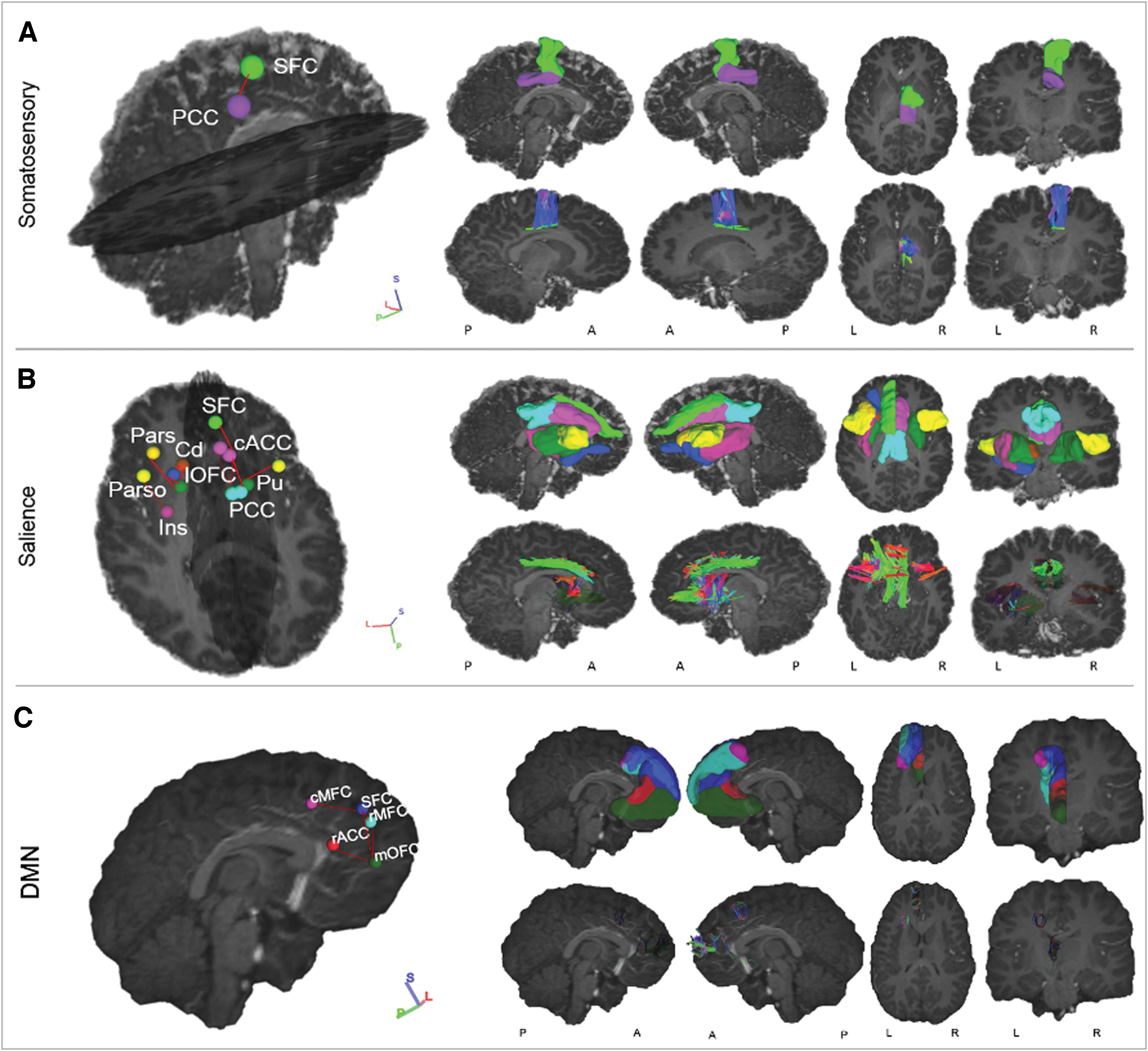
Networks with WM connections between GM regions showing significantly higher MD in CPHIV than CHU. The 3D volume representations were created in DSI studio. From left to right, each column shows: an oblique viewing angle of the GM regions and connections, right and left sagittal views, and axial and coronal views. In rows
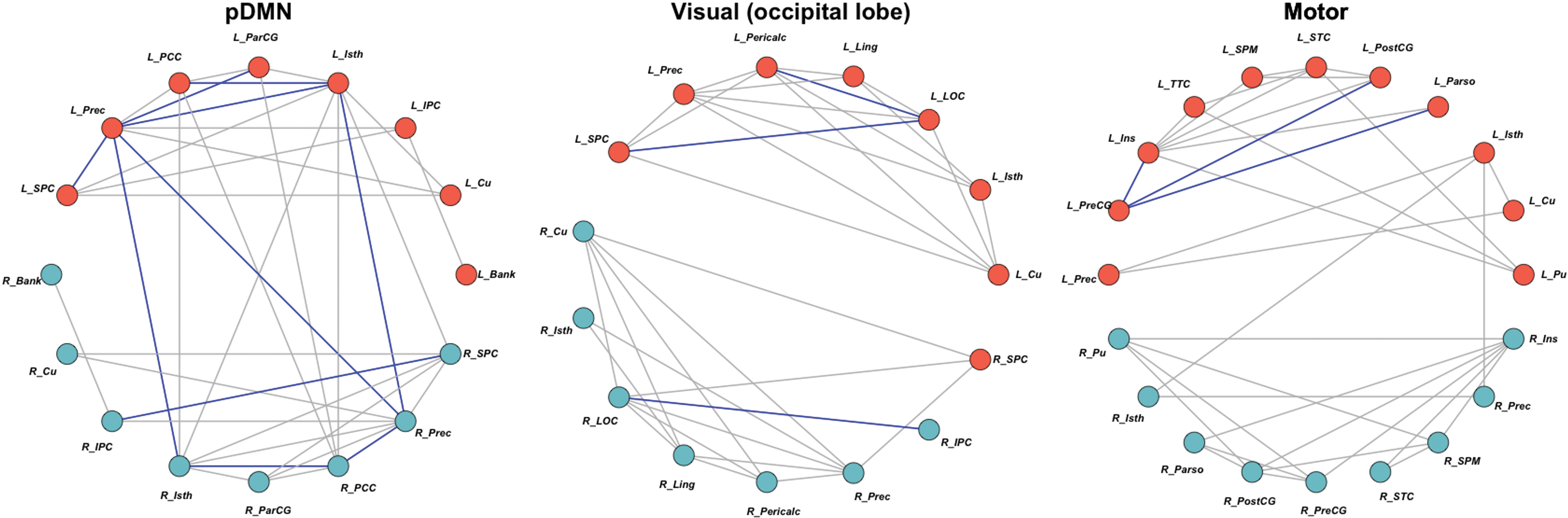
Networks with WM connections between GM regions showing significantly higher FA in CHEU than CHUU. Right hemisphere regions are light blue, whereas left hemisphere regions are coral. The blue connections represent WM connections that showed higher FA, whereas the gray connections represent the rest of the WM connections that were present and unaffected by HIV exposure. pDMN: FA was higher in CHEU compared with CHUU in the Cingulum between: L isthmus cingulate (L_Isth) and L posterior cingulate cortex (L_PCC), L_Isth and L Precuneus (L_Prec), L_Isth and R_Prec, L paracentral gyrus (L_ParCG) and L_Prec, L_Prec and L superior parietal cortex (L_SPC), L_Prec and R_Isth, R inferior parietal cortex (R_IPC) and R_SPC, R_Isth and R_PCC, R_PCC and R_Prec, and L_Prec and R_Prec. Visual (occipital lobe) network: The FA was higher in CHEU compared with CHUU in the Forceps Major between: L lateral occipital cortex (L_LOC) and L pericalcarine (L_Pericalc), L_LOC and L_SPC, and R_IPC and R_LOC. Motor: The FA was higher in CHEU compared with CHUU in the SLF between: L_PreCG and L parsopercularis (L_Parso), L_PreCG and L insula (L_Ins), and L_PreCG and L postcentral gyrus (L_PostCG). CHEU, children perinatally HIV-exposed but uninfected; CHUU, children who are HIV-unexposed and uninfected; pDMN, posterior DMN. Color images are available online.
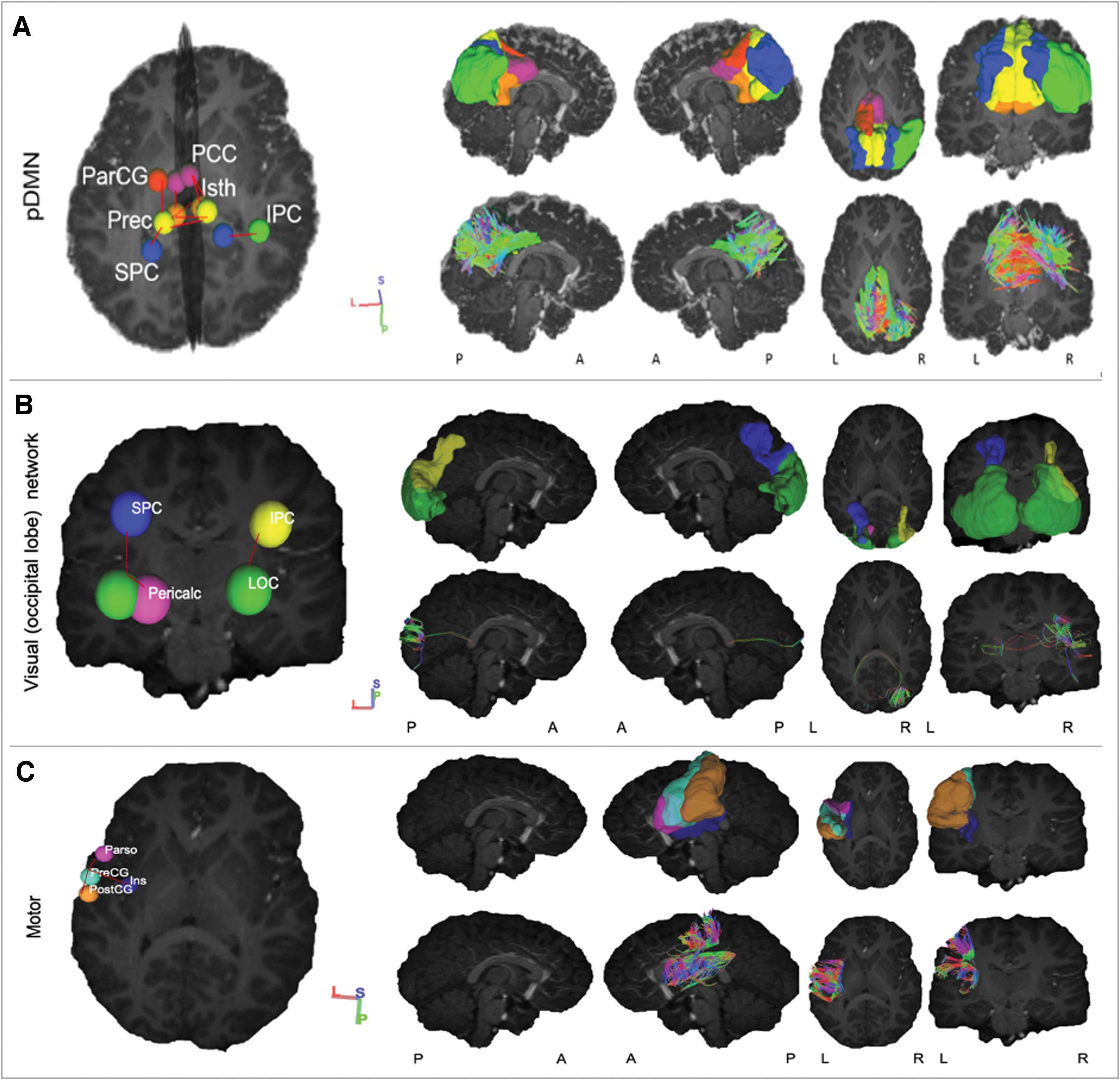
Networks with WM connections between GM regions showing significantly higher FA in CHEU than CHUU. The 3D volume representations were created in DSI studio. From left to right, each column shows: an oblique viewing angle of the GM regions and connections, right and left sagittal views, and axial and coronal views. In rows
Tracts Showing Significantly Lower (at Corrected p ≤ 0.05) Fractional Anisotropy in Children with Perinatally Acquired HIV Compared with Children Who Are HIV-Uninfected: Group Means (μ) and Standard Deviations (σ), Beta (β), Standard Error (Std Error), Standardized Beta (Std β), and Corrected p-Values for Fractional Anisotropy, Axial Diffusivity, and Radial Diffusivity, Respectively
Bold p-values are statistically significant (p-value < 0.05).
AD, axial diffusivity; CST, corticospinal tract; FA, fractional anisotropy; FDR, false discovery rate; GM, gray matter; L, left; RD, radial diffusivity; R, right; SLF, superior longitudinal fasciculus; WM, white matter.
Tracts Showing Significantly Higher (at Corrected p ≤ 0.05) Mean Diffusivity in Children with Perinatally Acquired HIV Compared with Children Who Are HIV-Uninfected: Group Means (μ) and Standard Deviations (σ), Beta (β), Standard Error (Std Error), Standardized Beta (Std β), and Corrected p-Values for Mean Diffusivity, Axial Diffusivity, and Radial Diffusivity, Respectively
Bold p-values are statistically significant (p-value < 0.05).
ATR, anterior thalamic radiation; DMN, default mode network; MD, mean diffusivity.
Tracts Showing Significantly Higher (at Corrected p ≤ 0.05) Fractional Anisotropy in Children with HIV Exposure But Uninfected Compared with Children Who Are HIV-Unexposed and Uninfected: Group Means (μ) and Standard Deviations (σ), Beta (β), Standard Error (Std Error), Standardized Beta (std β), and Corrected p-Values for Fractional Anisotropy, Axial Diffusivity, and Radial Diffusivity, Respectively
Bold p-values are statistically significant (p-value < 0.05).
Correlation to neuropsychological test scores
We found no significant correlations between DTI parameters of implicated tracts and neuropsychological test scores. Further, group comparisons (CPHIV vs. CHU and CHEU vs. CHUU) of the neuropsychological test scores showed lower scores for the CPHIV group compared with CHU only in the learning sub-scale of the KABC-II, as well as both the preferred and nonpreferred hand sub-scales of the Purdue pegboard test. There were no differences on any of the tests between CHEU and CHUU.
Discussion
To our knowledge, this is the first study to use tractography to quantify the properties of white matter within functional networks in pediatric HIV. An investigation of the properties of white matter tracts within RSNs provides a context for HIV-associated white matter and FC abnormalities. We found lower mean structural connectivity in CPHIV compared with CHU in a subset of the RSNs that we hypothesized—the CST and SLF within the somatosensory network, and forceps minor within the salience and motor networks, demonstrated lower FA; however, the cingulum in the somatosensory network, ATR, cingulum, SLF, and forceps minor in the salience network, and forceps minor in the DMN demonstrated higher MD. Only the ATR and forceps minor were among the tracts hypothesized. The observed networks and white matter tracts that fit our hypotheses provide evidence of a link between HIV-associated structural and functional abnormalities in this cohort.
In addition, CHEU demonstrated higher mean FA in the cingulum in the pDMN, forceps major in the visual occipital network, and SLF in the motor network. Although the pDMN and motor network were among the networks we predicted, we did not observe higher structural connectivity in the PCR nor the CST as hypothesized. Our results suggest that HIV- and HIV-exposure-related white matter abnormalities are focused within a few specific RSNs.
CPHIV versus CHU
A noteworthy aspect of our results is the recurrence of a few gray matter regions among particular RSNs. Seeds in the superior temporal cortex, superior frontal cortex, putamen, and posterior cingulate cortex are involved in connections within at least two of the four RSNs that demonstrated HIV-associated differences. The findings suggest that these regions are vulnerable to HIV and either influence or are influenced by white matter alterations. We have previously reported HIV-related structural differences in these regions within this cohort. At age 5, CPHIV demonstrated increased cortical thickness and decreased gyrification in the superior frontal and temporal cortex compared with CHU (Nwosu et al., 2021). At age 7, lower gyrification was reported bilaterally in the posterior cingulate cortex, and right superior frontal and temporal cortices (Nwosu et al., 2018). Further, we found putamen volume increases at age 5 (Randall et al., 2017), but volume reductions in this region compared with CHU at age 7 (Nwosu et al., 2018).
The HIV-related morphological changes previously observed may be related to the localized myelin reductions presented. The theory of tension-based morphogenesis hypothesizes that developing white matter produces localized fiber tension that brings densely interconnected regions closer together (Van Essen, 1997). Drawing regions of greater connectivity closer leads to shorter connections and faster transmission of action potentials, making that specific white matter connection more efficient. Decreases in myelination may contribute to reductions in fiber tension, and it subsequently increased thickness and less folding in cortical regions. This is a possible explanation for how the trajectory of white matter development influences the shape of gray matter regions. Reductions in myelin, which translate into the higher MD and RD values seen in the present study, could therefore have resulted in lower tension and ultimately increased thickness and less folding on gray matter regions.
The observed changes in FA and MD were largely attributable to increases in RD, which has been shown to be related to localized myelin reductions (Yan Aung et al., 2013). In healthy subjects, RD declines significantly with age whereas AD undergoes very little to no change with age (Lebel et al., 2017; Tamnes et al., 2018). Thus, because of the changes that perpendicular diffusivity is undergoing, any effect of HIV/ART can alter the growth trajectory. Consequently, the white matter deficits reported are most likely due to complications in myelination or axonal packing.
Somatosensory and motor networks
We found bilateral alterations between the superior temporal cortex and putamen in the somatosensory network. This connection was also affected in the motor network in the left hemisphere. As these two networks work together, these may be linked. Successful planning and efficient execution of movement by the motor cortex requires sensory input of the body surface, which is facilitated by the primary somatosensory cortex (Lee et al., 2013). Both motor and somatosensory projections connect to the putamen, which plays a role in modulating motor control of the limbs (Lanciego et al., 2012). The cortico–subcortical connections between the superior temporal cortex and putamen contribute to spatial awareness, and damage has been associated with spatial neglect (HO, 2001).
Though we were unable to label the connections between the putamen and superior temporal cortex in the somatosensory network, the connection between these regions in the left hemisphere in the motor network was identified as being part of the forceps minor. The forceps minor has not been described within somatosensory or motor function. Another major tract identified to be connected to the putamen in the somatosensory network is the CST. The somatosensory function of the CST is in relation to the spinal cord, where it receives and regulates input from the peripheral somatosensory receptors (Van Wittenberghe and Peterson, 2020), allowing it to control voluntary movement (Van Wittenberghe and Peterson, 2020). This further validates somatosensory-motor network interaction. Motor impairment has been associated with low AD in the CST of adults (Bernard et al., 2013). At age 5, we reported reduced FA in a region of the CST in the CPHIV compared with CHU (Ackermann et al., 2014). Studies on ART-naive CPHIV have reported poor motor and executive functioning (Hoare et al., 2012) as well as no improvement of motor function after 6 months of treatment (Smith et al., 2002). Further, a tractography study in older patients living with HIV and on cART showed compromised motor connections (Jahanshad et al., 2012). This suggests not only particular vulnerability of these regions but also a potential disruption in between-network communication among these networks.
In addition, we have previously reported lower FC in CPHIV compared with CHU between a seed in the left paracentral lobule in the somatosensory network and clusters in the salience and/or basal ganglia networks, including left and right cingulate gyri, left medial frontal gyrus, and the right anterior cingulate (Toich et al., 2018). In the current study, we observe alterations in the CST between the paracentral gyrus and the postcentral gyrus. In the MD results, we found the cingulum connecting the right posterior cingulate cortex to the right superior frontal cortex, suggesting a vulnerability of the cingulate cortex. Notably, our seed-based FC analyses at age 7 similarly demonstrated lower FC in CPHIV compared with CHU between the left posterior cingulate gyrus in the DMN and inferior and medial frontal regions.
Salience network
The salience network is a higher-order cognitive network that undergoes significant maturation during early adolescence. An important role of the salience network is modulating the activities of other functional networks (Bonnelle et al., 2012; Kumfor et al., 2015; Seeley et al., 2007; Walsh et al., 2016). It has been shown to demonstrate lower intra-network connectivity in adults living with HIV despite VL suppression (Chaganti et al., 2017) but not in children. Although previously we did not find altered FC within the salience network, both higher and lower FC between this and other networks were found in CPHIV (Toich et al., 2018). Notably, many of the gray matter regions in the salience network showing altered FC in that study overlap with gray matter regions showing altered white matter connections here, namely the anterior cingulate cortex, insula, and posterior cingulate cortex. Further, FC in the salience network (right inferior frontal gyrus and insula) was negatively associated with CD4% in infancy (Toich et al., 2018). The fact that white matter connections to the right insula did not demonstrate alterations in the present study suggests that the observed FC alteration may not be due to changes in structural connectivity to this region.
Connections involving the posterior cingulate cortex, putamen, and caudal anterior cingulate cortex were repeatedly implicated. The salience network interacts with the somatosensory network in that afferent signals from sensory receptors travel to the brain, where the salience network is recruited for conscious perception of the signal (Walsh et al., 2016). The connection in the somatosensory network with higher mean MD involved the posterior cingulate cortex. The overlapping regions within network connectivity may point to changes between networks.
The DMN also shows between-network relationship with the salience network. This network is involved in episodic memory processing and consists primarily of the posterior cingulate cortex, the medial temporal lobe, and the medial prefrontal cortex. Coordinated activity between these two higher-order cognitive networks allows for one to behave efficiently. Salience network structural connectivity has been reported to predict DMN function (Bonnelle et al., 2012), and damage to tracts in the salience network has been associated with weaker FC in the DMN (Kumfor et al., 2015). In the current study, we found a connection between the superior frontal gyrus and the right caudal anterior cingulate to be altered in the salience network of the CPHIV and out of all four of the implicated connections in the DMN, the superior frontal gyrus was involved in each one. This gray matter region has been reported to have higher cortical thickness and decreased gyrification in CPHIV of the same cohort at age 5 compared with CHU (Nwosu et al., 2021), suggesting that these may be ongoing changes.
Default mode network
The DMN showed decreased intra-network FC using seed-based FC analysis in CPHIV in this cohort at age 7 (Toich et al., 2018) and associations of FC with clinical measures of health in another cohort of perinatally infected youth (Herting et al., 2015). In this analysis, we find altered structural connectivity (higher MD) within the forceps minor in the DMN in the CPHIV. Notably, in both the functional analysis and the current structural analysis, the medial frontal cortex was involved, which suggests that the effects of HIV infection in this region have both functional and structural consequences.
CHEU versus CHUU
The FA increases in CHEU compared with CHUU are consistent with findings of higher FA from our voxelwise-DTI analysis in CHEU from the same cohort in a region of the SCR (Jankiewicz et al., 2017) and by Tran et al. (2016) in middle cerebellar peduncles of HEU neonates. Though higher FA is usually associated with healthier white matter, some studies in adolescents have also reported increased FA in other neurodevelopmental disorders, including attention deficit/hyperactivity disorder and autism (Andrews et al., 2019; Davenport et al., 2010; Peterson et al., 2011). A 2018 study reported a more than twofold increase in odds for autism, disturbance of emotions, hyperkinetic syndrome, and developmental delay in HEU compared with matched HUU children (Piske et al., 2018). Further investigation within our cohort is needed in relation to possible diagnoses of neurodevelopmental disorders.
FA as an indicator of axonal integrity is a measure of axonal diameter, axonal packing density, and axonal branching (Hoeft et al., 2007). An alteration in any of these will lead to either an increase or a decrease in FA. This, together with the understanding that FA in childhood increases with age, at a certain rate over a certain time (with rates and times varying regionally) (Lebel et al., 2017), means that premature increases in FA may disrupt typical development. In the implicated tracts in the motor network, CHEU also had lower RD compared with the CHUU. Higher FA and lower RD could mean increased myelination, resulting in a more dense axonal packing and ultimately increased anisotropy/higher restriction of diffusion.
Posterior default mode network
The pDMN had the most affected connections within this part of the analysis. Notably, all the affected white matter connections were identified to be part of the cingulum. Further, most affected connections were either to the precuneus or to the isthmus cingulate. The precuneus is involved in reflective self-awareness, episodic memory, and visuospatial processing (Cavanna and Trimble, 2006). The isthmus cingulate is involved in memory processing (Nielsen et al., 2005), whereas connectivity and structure alterations have been associated with depression (Grieve et al., 2013; Korgaonkar et al., 2014). The fact that the pDMN was affected in this analysis and the DMN within the CPHIV versus CHU analysis further speaks to the functional and structural vulnerability of this network to HIV and exposure.
Visual (occipital lobe) network
In this network, all implicated white matter connections were part of the forceps major and involved the lateral occipital cortex. This structure, which is involved in visual attention, has been noted to have HIV-associated reduced FC in adults as early as within a year of infection (Wang et al., 2011).
Motor network
We found all affected white matter connections in this network to be part of the SLF and to involve connections to the left precentral gyrus. All the connections were in the left hemisphere, where the SLF is involved in verbal working memory (Peters et al., 2012). This tract only reaches maximum maturity between the age of 19 and 25 (Lebel et al., 2017; Tamnes et al., 2018), making age 7 a vulnerable age. Results from the FC analysis of CHEU versus CHUU showed increased FC in clusters within the DMN, pDMN, executive, and motor networks in CHEU compared with CHUU (Toich, 2015). Though the sample size was small (8 CHEU, 10 CHUU) compared with the size of the current study (19 CHEU, 27 CHUU), results from the present study suggest that altered FC due to HIV exposure in pDMN and motor network may be related to altered structural connectivity.
Correlation to neuropsychological test scores
Groups showed no differences on the neuropsychological tests except for the learning sub-scale of the KABC-II, and both the preferred and nonpreferred hand sub-scales of the Purdue pegboard test, which were lower in CPHIV than CHU. Studies correlating DTI metrics with neuropsychological scores in CPHIV and CHEU are few but collectively report an association between brain microstructural integrity and neurocognitive function (Ackermann et al., 2020; Hoare et al., 2012; Jahanshad et al., 2012; Tran et al., 2016; Yadav et al., 2020). Lower FA and/or higher MD was associated with poor neuropsychological performance in cART-naive adolescents living with HIV (Hoare et al., 2012). In another study, lower FA and/or higher MD in CPHIV and CHEU were both negatively and positively associated with neuropsychological scores (Yadav et al., 2020). Though Tran et al. (2016) reported higher FA and lower MD in WM tracts of CHEU compared with CHUU, they too report associations with neuropsychological scores. Further, at age 5, the CPHIV in the current study reported a negative correlation between RD and neuropsychological scores in WM, which included the SLF and ILF (Ackermann et al., 2020). The association between DTI metrics and neuropsychological scores also seems to be present even where there are no group differences between CHEU and CHUU (Jahanshad et al., 2016). Some of the white matter tracts reported in the current study have been associated with cognitive behavior (Frye et al., 2011; Grundy et al., 2017; Tzourio-Mazoyer, 2016). The SLF integrity has been associated with reading and attention-related skills (Frye et al., 2011). In addition, the forceps minor is involved in cognitive function (Tzourio-Mazoyer, 2016), and reduced FA has been suggested as a predictor of cognitive impairment (Grundy et al., 2017). Given these reported links between specific white matter tracts and cognitive function, we explored relationships between DTI outcomes in all our affected tracts (in both CPHIV vs. CHU and CHEU vs. CHUU) and neuropsychological measures. We found no statistically significant correlations. The findings suggest that the white matter differences observed may be either too subtle to translate into neuropsychological alterations or relate to different neuropsychological domains. It is also possible that the functional implications of altered white matter integrity may only become apparent at later ages. Further, though participants in the study come from similar communities and backgrounds, we cannot rule out the possibility of environment-induced white matter alterations. White matter integrity has been associated with anxiety (Lu et al., 2018), and HIV-related maternal stress and anxiety has been negatively associated with parenting skills (Murphy et al., 2010). More information pertaining to settings in which the children are growing up may shed more light on the causes of the white matter differences we see here.
Conclusions
In conclusion, as hypothesized, we found altered structural connectivity in the somatosensory, salience, default mode, and motor networks of CPHIV compared with CHU. The default mode, salience, and somatosensory networks had been implicated in the same cohort at the same age in a previous study of FC, which suggests that damage or developmental delay in white matter may be related to or responsible for the HIV-associated functional abnormalities. Further, the regions implicated seem to recur in all four implicated networks and have been implicated in morphometry studies of the same cohort. Together, these results suggest that white matter alterations influence gray matter morphology and between-network FC. Both FA and MD differences were largely attributable to RD increases, suggesting demyelination/dysmyelination due to HIV/ART. In addition, we examined the effects of HIV exposure and found increased structural connectivity in the visual network, pDMN, and motor network. Higher FA is consistent with the previous voxelwise-DTI study performed on the same cohort and suggests that HIV/ART exposure may result in premature axonal maturation in the developing brain.
Footnotes
Acknowledgments
The authors thank the participants and their parents for being willing to take part in this study, research assistants Lungiswa Khethelo and Thandiwe Hamana for their expertise in supporting the children during neuroimaging, and the radiographers at CUBIC.
Authors' Contributions
Joanah Madzime and Marcin Jankiewicz were involved in designing and performing data analyses. Ernesta Meintjes, Andre J.W. van der Kouwe, Barbara Laughton, and Mark Cotton conceived, designed, and obtained funding for the study. Joanah Madzime, Martha Holmes, and Marcin Jankiewicz drafted the article and all other authors provided a critical revision of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by NIH grants R01HD099846, R01DC015984, R01HD071664, R21MH096559, and R21MH108346; South African National Research Foundation (NRF) grants 48337, 99069, and 78737; UCT VC Interim Funding (2011/2012); and the South African Medical Research Council (SAMRC). The research and writing of the article were supported by the NIMH and NINDS Intramural Research Programs (ZICMH002888) of the NIH/HHS, USA. Support for the CHER study, which provided the infrastructure for the neurodevelopmental substudy, was provided by the US National Institute of Allergy and Infectious Diseases through the CIPRA network, Grant U19 AI53217; the Departments of Health of the Western Cape and Gauteng, South Africa; and GlaxoSmithKline/Viiv Healthcare. Additional support was provided with Federal funds from the National Institute of Allergy and Infectious Diseases, National Institutes of Health, United States Department of Health and Human Services, under Contract No. HHSN272200800014C. Permission to conduct the substudy on this cohort was granted by Doctors Avy Violari, Shabir Madhi, and Mark Cotton and the CHER steering committee.