
Abstract
Background:
Aging is accompanied by a gradual deterioration in multiple cognitive abilities and brain structures. Both cognitive function and white matter (WM) structure are found to be associated with neurodegeneration diseases and correlated with sex during aging. However, it is still unclear whether the brain structural change could be attributable to sex, and how sex would affect cognitive performances during aging.
Materials and Methods:
Diffusion magnetic resonance imaging (MRI) scans were performed on 1127 healthy participants (age range: 21–89) at a single site. The age trajectories of the WM tract microstructure were delineated to estimate the turning age and changing rate between sexes. The canonical correlation analysis and moderated mediation analysis were used to examine the relationship between sex-linked WM tracts and cognitive performances.
Results:
The axon intactness and demyelination of sex-linked tracts during aging were multifaceted. Sex-linked tracts in females peak around 5 years later than those in males but change significantly faster after the turning age. Projection and association tracts (e.g., corticospinal tracts and parahippocampal cingulum) contributed to a significant decrease in visuospatial functions (VS) and executive functions (E). We discovered that there is a stronger indirect effect of sex-linked tracts on cognitive functions in females than in males.
Conclusion:
Our findings suggest that the vulnerable projection and association tracts in females may induce negative impacts on integrating multiple functions, which results in a faster decrease in VS and E.
Impact statement
By recruiting a healthy population for diffusion magnetic resonance imaging (MRI) scan, we demonstrated that the age-related changes of white matter (WM) integrity were manipulated by sex. Sex-linked tracts in females reached the turning age 5 years later, but change faster than those in males after the turning age. The most significant sex effect was found in projection and association tracts. Our results indicated that sex affected both WM microstructure and cognitive functions and was further involved in the mediation of the age-WM-cognition relationship. The vulnerability of projection and association tracts in females may induce negative impacts on visuospatial and executive functions.
Introduction
Aging is accompanied by a gradual deterioration in cognitive abilities, especially in the working memory and visuospatial functions (VSs) (Cansino et al., 2013; Pauls et al., 2013; Li et al., 2017; Voyer et al., 2017). Previous studies have indicated that sex differences in brain structural features may mediate how males and females perform cognitively (Gur et al., 1999; Chou et al., 2011; Dunst et al., 2014; Tunç et al., 2016). In general, females excel in both verbal fluency and verbal memory (VM) tasks (Rilea et al., 2004; Jones and Healy, 2006; Munro et al., 2012), whereas males show better working memory capacity in both numbers and visuospatial elements (Dadin et al., 2009; Longenecker et al., 2010).
The interaction between sex and age is also evident that, for example, the verbal superiority in females diminishes due to the increasing cognitive deterioration after the age of 55 (Maitland et al., 2004); and the verbal-based cognitive functions in males decline acceleratively after the age of 50 (Lundervold et al., 2014; McCarrey et al., 2016).
Aging is also affected by the white matter (WM) tracts beneath the interregional signal transmission. Brain shrinking in WM was also found in the advanced age, with a mixed consequence of multiple microstructural changes, such as demyelination and axonal damage (Miller and Corsellis, 1977; Peters, 2006; Salat et al., 1999; Tang et al., 1997). The WM tracts that connect distinct functional cortices are associated with specific cognitive performances (Roberts et al., 2013). For example, the corpus callosum links to executive function (E) and processing speed (Frederiksen, 2013; Prendergast et al., 2015); and the superior longitudinal fasciculus (SLF) is correlated with a language function (L) (Chen et al., 2009).
Imaging and postmortem studies have shown that age-associated brain change occurs predominantly in the WM (Allen et al., 2005; Jernigan et al., 2001; Piguet et al., 2009), and the rate of WM shrinkage accelerates precipitously at the fifth decade of life (Raz and Rodrigue, 2006). In terms of sex effect, males showed higher WM directivity, while females showed higher WM complexity (Hemmen et al., 2017; Menzler et al., 2011; Ritchie et al., 2018); however, the role of sex effect on the age trajectories of WM tracts and whether it may cause differences in cognitive abilities are still under debate. One study indicated that females showed a faster decline in corpus callosum (Sullivan et al., 2010), while others stated that sex only marginally affects the aging model (Lebel et al., 2012; Slater et al., 2019; Westlye et al., 2010). Altogether, the age-associated change of WM is corresponding to the onset timing of cognitive decline (Salthouse, 2009; Singh-Manoux et al., 2012; Thomas et al., 2016).
To clarify the role of sex in the age-WM-cognition relationship, we established the sex-linked aging model of specific WM tracts. By analyzing 1331 single-site diffusion magnetic resonance imaging (MRI) data in the age range of 20 to 89, we evaluated 20 major cerebral changes of WM tracts with diffusion indices across the life span. A canonical correlation analysis (CCA) was used to identify the relationship between the tract that most associated with sex-linked cognitive functions, including VS, E, L, and VM. Mediation analysis was established to examine whether the age- and sex-linked cognitive functions are mediated by the WM tracts and moderated by sex.
This study aims to investigate the sex-linked model for cognitive aging with its neural basis and test the following hypotheses: (1) the sex effect affects both age-related cognitive and WM microstructural changes, (2) where males and females show different aging patterns in turning ages, changing rates, and magnitudes. (3) The changes of brain WM mediate the cognitive aging process in which sexes contribute unequally in this relationship.
Materials and Methods
Participants
Two cohorts of 1331 right-handed participants were enrolled. The first cohort included 972 participants, community-based aged residents (age range: 50–87) recruited through the household registrations in I-Lan County of Taiwan (Liu et al., 2014). The second cohort included 359 participants (age range: 21–89), recruited through advertisements in local communities and universities from northern Taiwan. All participants were self-reported with good visual and auditory abilities, no medical history of major neurological or psychiatric diseases, and were able to take the cognitive tests.
The intactness of global cognitive performance for each participant was defined by Mini-Mental State Examination (MMSE) (Folstein et al., 1975; Shyu and Yip, 2001). Raw scores are considered the standard criterion, where well-educated people (education ≥6 years) showed a score greater than 24, whereas less-educated people (education <6 years) showed at least 14 (Sun et al., 2014). In particular, the participants of the first cohort received extra cognitive tests for the evaluation of their detailed cognitive function in four axes, including VM, L, E, and VS (Wu et al., 2015). The delayed-free recall in the Chinese Version Verbal Learning Test determined the performance of VM (Chang et al., 2010). Boston Naming Test (Mack et al., 1992) and the category (animal) Verbal Fluency Test were used for assessing the L. E was estimated by the backward digit-span (Hester et al., 2004) and the Clock Drawing Test (Rouleau et al., 1992). VS was assessed by the copy test of the Taylor Complex Figure Test (Taylor, 1969).
Experiments of the two cohorts were approved, respectively, by the Institutional Review Board of Taipei Veterans General Hospital (2013-06-021AY) and the Institutional Review Board of National Yang-Ming University (2018-05-003B).
Image acquisition
MRI scans of the two cohorts were performed with identical imaging protocols on the same 3T Siemens MR scanner (Siemens MAGNETOM Tim Trio, Erlangen, Germany) equipped with a 12-channel head coil at the National Yang-Ming University, Taipei, Taiwan. All participants received identical pulse sequences, including a set of diffusion tensor images (DTI) and a high-resolution T1-weighted (T1W) image. T1W image was scanned with the magnetization-prepared rapid gradient-echo protocol with TR/TE/TI = 3500/3.5/1100 ms; flip angle = 7°; voxel size = 1 × 1 × 1 mm3 without interslice gap; 192 sagittal slices; and FOV = 256 × 256 mm2. For the DTI data, single-shot spin-echo echo-planar diffusion-weighted images were utilized with the following parameters: TR/TE = 11,000/104 ms; voxel size = 2 × 2 × 2 mm3; 70 contiguous axial slices; FOV = 128 × 128 mm2; 30 noncollinear gradient directions with b value of 1000 s/mm2, and three additional null images (b = 0 s/mm2) as reference images with NEX = 3. All the images were first visually examined by two analysts (C.-C.H.H. and C.-C.H.) to exclude the data with (1) brain lesion (unreported tumor, stroke, and cyst) and (2) severe motion in T1W and DTI, which would generate ill registration and tensor fitting.
Image preprocessing
MRI data were all processed using FSL V5.0.9 (Functional Magnetic Resonance Imaging of the Brain Software Library). For each participant, the preprocessing procedures included skull removal, tissue segmentation for T1W images, motion correction, and eddy current correction for diffusion-weighted images. The diffusion tensor model was reconstructed and the quantitative indices of fractional anisotropy (FA), axial diffusivity (AD), mean diffusivity (MD), and radial diffusivity (RD) were calculated.
Twenty Johns Hopkins University (JHU) WM tracts with the probability distribution of projection tracts (anterior thalamic radiation [ATR] and corticospinal tract [CST]), association tracts (inferior longitudinal fasciculus [ILF], inferior fronto-occipital fasciculus [IFO], uncinated fasciculus [UNC], SLF and its temporal projection [tSLF], cingulum in cingulum cortex [CgC], and parahippocampal cingulum [CgH]), and commissural tracts [forceps minor (Fmi) and forceps major] (Hua et al., 2008).
The JHU tract probabilistic map was transformed from standardized Montreal Neurological Institute (MNI) space (Jenkinson and Smith, 2001) to diffusion native space to estimate the microstructures with the diffusion indices of the tracts of interest. First, the diffusion null image was coregistered to the corresponding T1W image of each participant. Second, the T1W image was nonlinear registered and spatially normalized to the MNI space. By multiplying the two transformation matrices from the above steps, the JHU tract probabilistic map was applied to the backward transformation matrix and transformed from MNI space to each individual's diffusion native space. The diffusion indices for each tract were extracted with an FA threshold of 0.2 to excluded the portion of gray matter or cerebral spinal fluid. The diffusion indices (including FA, AD, MD, and RD) representing the microstructure states for each tract were then weighted by the probabilistic distribution and normalized by the total sum of the probability (see Supplementary Methods for more details).
Curve fitting and model selection
Four curve models, linear, quadratic, Poisson (Lebel et al., 2012; Yeatman et al., 2014), and piecewise, were applied to model the trajectories of sex differences in age-related change of cognitive functions and WM microstructures. The models are defined as follows: Linear:
Quadratic:
Poisson:
Piecewise: a three-parameter piecewise linear model was utilized to fit the transition plateau after the development and the change during aging. The first two parameters are the intercepts and the time point, which indicated when the measurement started to change. The third parameter is the slope of change during aging (see Supplementary Methods for more details).
For all models, the dependent variable, y, is the vector of measurements (cognitive performances and diffusivities) and each
For the WM microstructure, the total intracranial volume (TIV) and education year were used as covariates to control the effects (Dufouil et al., 2003; Takao et al., 2011b). The best-fitted model was evaluated by the goodness of fit with maximal adjusted coefficient of determination (R adj 2 > 0.1) (Draper and Smith, 2014), and the majority in 20 best-fitted trajectories would be selected to apply to all cognitive functions and tracts. After the R adj 2 estimation of WM microstructure fitting trajectories, the quadratic model was selected as the best-fitted model and applied to all diffusivity indices, which had higher goodness of fit for trajectories (R adj 2 > 0.1, Supplementary Tables S2-1, S2-2, S2-3, and S2-4).
The sex difference in an age-related change of WM microstructure was observed in three different views: (1) the sex main effect, (2) the turning age (the derived vertex), and (3) the degree of the curvature. The degree of the curvature was controlled by the quadratic coefficient of a quadratic function; a larger magnitude corresponded to a sharply curved trajectory, which may indicate a faster change of diffusion indices over time. The significance of each fitting coefficient was tested by the t-statistic hypothesis that determines whether it is equal to zero or not. For the derived parameter, turning age and the significance of sex differences were measured by a permutation test for 5000 times.
CCA and mediation analysis
The correlation of sex-linked WM trajectories and the cognitive performance were evaluated by CCA and mediation analysis. We modified the CCA code released by Smith and colleagues (2015) for our data set. The relationship between sex-linked WM measures and cognitive functions was initially identified and generated as the optimal linear combination that maximally correlated with each other, while the effects of TIV and education were controlled. The correlation between the variables and the opposite canonical covariate was achieved, and the significance was estimated via 10,000 permutations.
After the CCA model was trained, moderated mediation analysis was achieved using a moderation model (model 1) and a moderated mediation with conditional indirect effect (model 8) in PROCESS macro (v.2) on SPSS (v.20) statistical software (Hayes, 2013). The bootstrapping method was adopted by the PROCESS to assess the significance of the indirect effects by 95% confidence intervals (CIs). The correlated components of WM tracts and cognitive functions identified by the CCA steps were applied to the mediation analysis. Model 1 tested the relationship modulated by the sexes between age and cognitive functions. Model 8 tested the mediation of the component of WM tracts to age and cognitive functions, and sex acts as the moderator in the model.
Experimental design and statistical analyses
The fitting models of cognitive functions and WM diffusivity both included an additional sex term to each variable and added education year as covariate (Dufouil et al., 2003). For the WM microstructure, TIV was included as an additional covariate due to the evidence that head size would affect the value of diffusion indices (Takao et al., 2011b). The significance of the sex main effect and changing rate was tested by the t-statistic hypothesis to investigate whether it is equal to zero or not. For the derived parameter, turning age and the significance of sex differences were measured by a permutation test for 5000 times. The sex term acts as the moderator in the mediation analysis. For the mediation analysis, 95% of a bias-corrected bootstrap model with CIs was resampled 10,000 times.
Results
Demographics
After excluding impaired global cognitive function and poor image quality, we analyzed MR data from 1127 participants, with case matching of age between sex groups. Among the participants, 819 were from the first cohort (mean age: 62.7 ± 8.1 years; range: 50–87 years; 49.3% males), and 308 participants were from the second cohort (mean age: 49.6 ± 19.1 years; range: 21–89 years; 49.0% males). All the participants were defined as intact global cognitive performance by MMSE. The group differences between males and females were found in education year, MMSE score, and TIV (p < 0.001). In the cognitive performances evaluated with the first cohort (819 participants were included), males had significantly better E, VS, and L than females. Although females' performance of VM surpassed that of the males in general, however, there was no significance between the groups in our study (p = 0.333). The demographic details are shown in Table 1.
Demographic Characteristics of the Study Groups
The assessment of cognitive functions was only acquired from the first cohort (N = 819).
MMSE, Mini-Mental State Examination; SD, standard deviation; TIV, total intracranial volume.
Cognitive performance in sexes
The age-related cognitive trajectories were limited to people older than 50 years (first cohort) who had cognitive assessment. The linear model was selected with maximal R adj 2 in E and L (Supplementary Table S1). Cognitive performances decreased along the aging process (R adj 2[VS] = 0.35, R adj 2[E] = 0.55, R adj 2[L] = 0.47, and R adj 2[VM] = 0.18). Visual spatial function and E showed significant sex main effect and age/sex interaction, in which the findings indicated that females revealed a significantly larger decrease ([VSage] = −0.02, [VSsex] = 1.52, [VSsex×age] = −0.03; [Eage] = −0.01, [Esex] = 1.50, [Esex×age] = −0.03; p < 0.01). For language and VM, there was no difference in either age/sex interaction or the sex main effect (Fig. 1).

Age-related cognitive change. The cognitive task including VS, E, L, and VM. Fitting curves with a 95% CI are separated into two colors representing different sexes (blue: male; red: female). Red and blue dots represent females and males. CI, confidence interval; E, executive function; L, language function; VM, verbal memory; VS, visuospatial function. Color images are available online.
Age-related WM trajectories
The quadratic model was selected as the best-fitted model (R adj 2 > 0.1, Supplementary Tables S2-1, S2-2, S2-3, and S2-4). Overall, the FA showed quadratic inverted U-shaped patterns that decreased with advanced age. Other indices (AD, RD, MD) showed opposite patterns with quadratic U-shaped trajectories (Fig. 2A and Supplementary Fig. S2). By observing the age trajectories, we discovered that a flat-fitted line was corresponding to small R adj 2 values (Table 2).
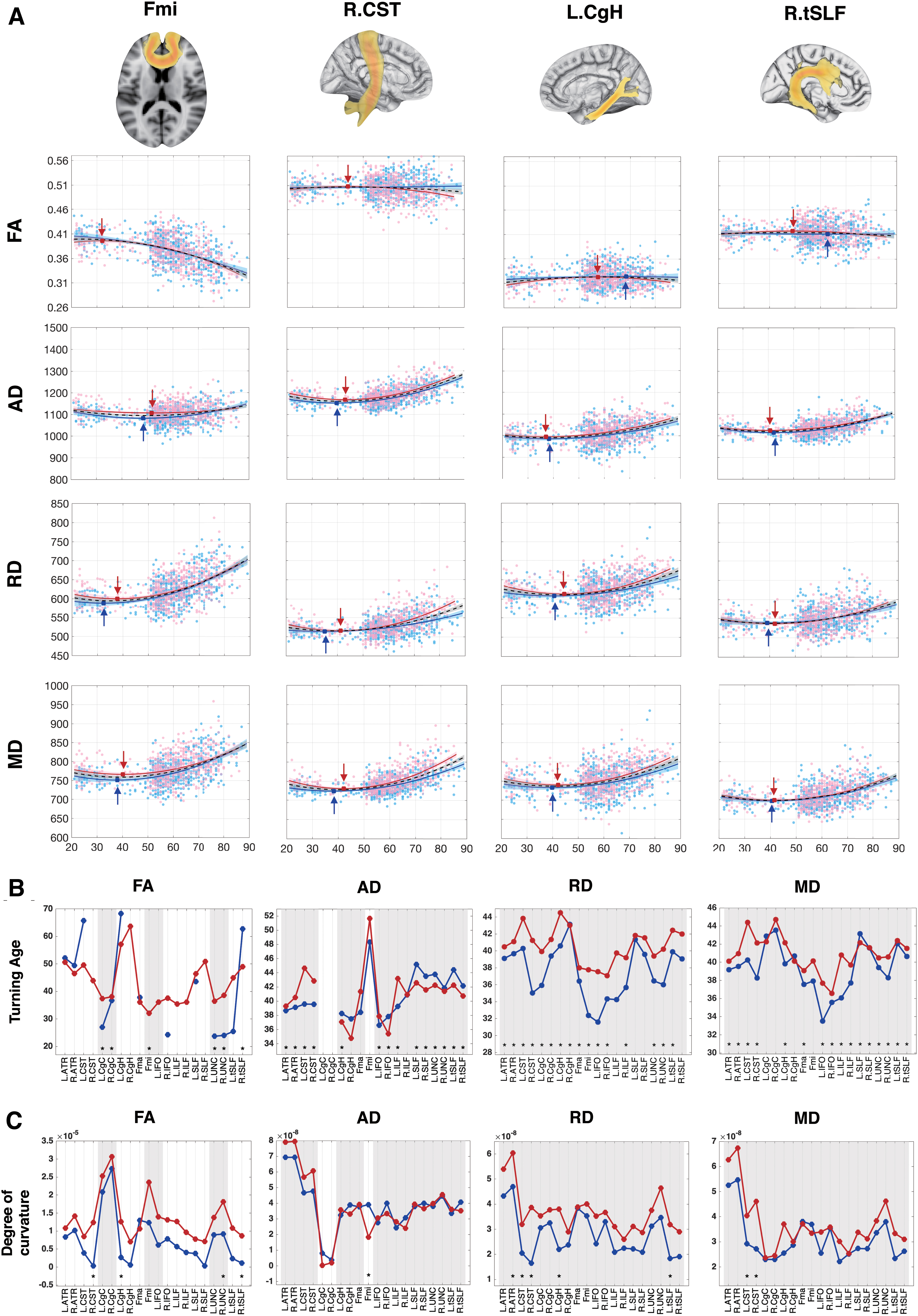
Regional fitting curve of diffusivity indices and derived turning ages and changing rates.
Adjusted R Square Value from Fitting Model of Each Tract
Tracts average diffusivity (FA, AD, MD, and RD) was regarded as dependent, and sex, age, age2, age × sex, age2 × sex, TIV, and education year as independent variables.
Represented not significant fitting, where no symbols mean p value smaller than 0.000625 (0.05/80, Bonferroni corrected).
AD, axial diffusivity; FA, fractional anisotropy; Fma, forceps major; Fmi, forceps minor; MD, mean diffusivity; RD, radial diffusivity; TIV, total intracranial volume.
Sex effect
The WM trajectories showed significant differences between sexes in left CgH (t = −2.31, p = 0.021), Fmi (t = −2.00, p = 0.046), and right UNC (t = −2.04, p = 0.042) in FA. Bilateral CST (Left: t AD = 2.67, p = 0.008, t MD = 2.78, p = 0.006; Right: t AD = 2.5, p = 0.012; t MD = 3.43 p < 0.001) and left ILF (t AD = 2.04, p = 0.042; t MD = 2.03, p = 0.042) had significantly greater AD and MD in females. For RD, significant sex main effects were found in right CST (t RD = 3.00, p = 0.003) and left CgH (t RD = 2.13, p = 0.033). The fitting results with significant nonzero effect on sex terms are summarized in Table 3, where sex effects were found in Fmi, right tSLF, UNC, and ATR, left CgH and ILF, and bilateral CST and tSLF.
Results from Fitting Model of 17 Sex-Linked White Matter Age Trajectories
Tracts average diffusivity (FA, AD, MD, and RD) was regarded as independent, and sex, age, age2, age × sex, age2 × sex, TIV, and education year as dependent variables. For the sex term, positive t value indicates that females were higher than males. For the quadratic term of age, negative in FA and positive in AD, RD, and MD of t value represented a faster change rate in females.
p < 0.05.
False discovery rate-corrected significance (q < 0.05).
T, t value; F, f value; R 2, coefficient of determination.
Turning age
The turning ages of AD, RD, and MD were in the range from 30 to 45, while FA varies from 20 to 70 (Fig. 2B and Supplementary Table S3). There was no specific trend in turning ages between males and females in AD. In RD, all tracts in males reached the turning ages earlier than females (male: 31.6–43.1; female: 37.1–44.5). All MD trajectories of males met the peak earlier (male: 33.5–43.5; female: 36.6–44.7), except left CgC (male: 42.9; female: 42.3) and right CgH (male: 40.7; female: 40.1). All significant differences of turning ages between males and females were less than 5 years in AD and MD. Differences greater than 5 years were only shown in RD, which were right CST (male<female = 6.2 years), Fmi (male<female = 5.4 years), and left IFO (male<female = 6.0 years). For FA, significantly later turning ages in females were found in bilateral CgC (male/female, R: 36.7/38.1; L: 27.0/37.4), UNC (male/female, R: 24.0/38.6; L:23.7/36.5), and Fmi (male/female: 9.6/32.1). Multiple derived turning ages showed extremely small or large value, which is outside the cohort age range (smaller than 21 or larger than 89). Those extreme values were derived by the trajectories that are relatively stable and with small R adj 2 values across the life span, including the right-sided CgH, right CST in FA, and bilateral CgC in AD (Table 2 and Supplementary Fig. S2).
Changing rate
Females showed faster changes than males (T age 2 ×sex) in FA, MD, and RD (Fig. 2C and Table 3). Significant accelerated decrease of FA in females was found, including right CST (β = −1.27 × 10−5, p = 0.009), UNC (β = −8.95 × 10−6, p = 0.041), tSLF (β = −9.80 × 10−6, p = 0.037), and left CgH (β = −9.94 × 10−6, p = 0.028). On the contrary, females also showed faster increase in MD and RD, including left tSLF (β = 1.35 × 10−8, p = 0.038), CgH (β = 1.60 × 10−8, p = 0.026), right ATR (β = 1.34 × 10−8, p = 0.040) of RD, and bilateral CST (L/R: βRD = 1.15 × 10−8/2.22 × 10−8, pR D = 0.041/<0.001; βMD = 1.12 × 10−8/1.89 × 10−8, pM D = 0.023/<0.001) of both RD and MD. The AD of Fmi was the only tract that showed accelerated increase in males (β = −2.08 × 10−8, p = 0.017).
CCA result
The sex-linked WM trajectories and cognitive performances were found to be correlated via CCA. The first CCA component showed a striking correlation between WM microstructure status and cognitive performances (r = 0.493, p < 0.001), while the second component was insignificant (r = 0.137, p = 0. 892) and was not included in the further mediation analysis. Right tSLF was the only tract out of the 17 sex-linked tracts that showed no significant contribution through bootstrapping. CST has contributed most across four indices, FA, AD, RD, and MD. See Figure 3 for the relationship between WM microstructure and cognitive performance, and Table 4 for the canonical coefficient and permutation p value details of the first CCA component.
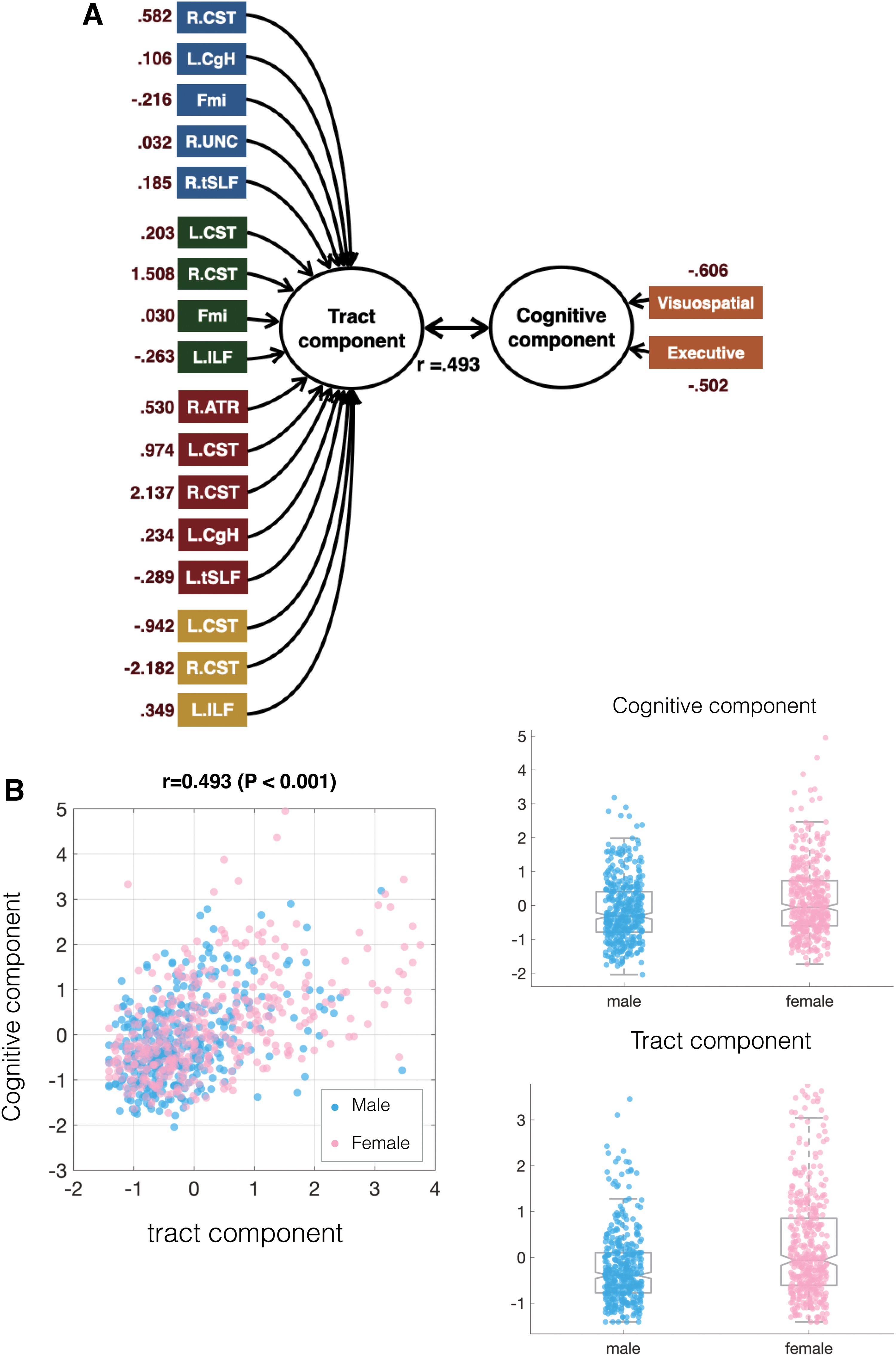
The construction of CCA.
Canonical Correlation Analysis
Mediation analysis
Model 1 indicated a significant age/sex interaction where females showed greater effect between age and cognitive scores (b = 0.233, p < 0.001). Via the moderated mediation analyses, both sexes showed notable indirect effects. The indirect effects of males and females, respectively, are 0.0617 (CI = [0.019–0.107]) and 0.0706 (CI = [0.022–0.122]). Females showed a significantly larger effect (equality test: differences = 0.0090, CI = [0.006–0.012]). Males showed the full mediation effect, whereas females showed both direct effects of age and indirect effects that are mediated by WM (Fig. 4).
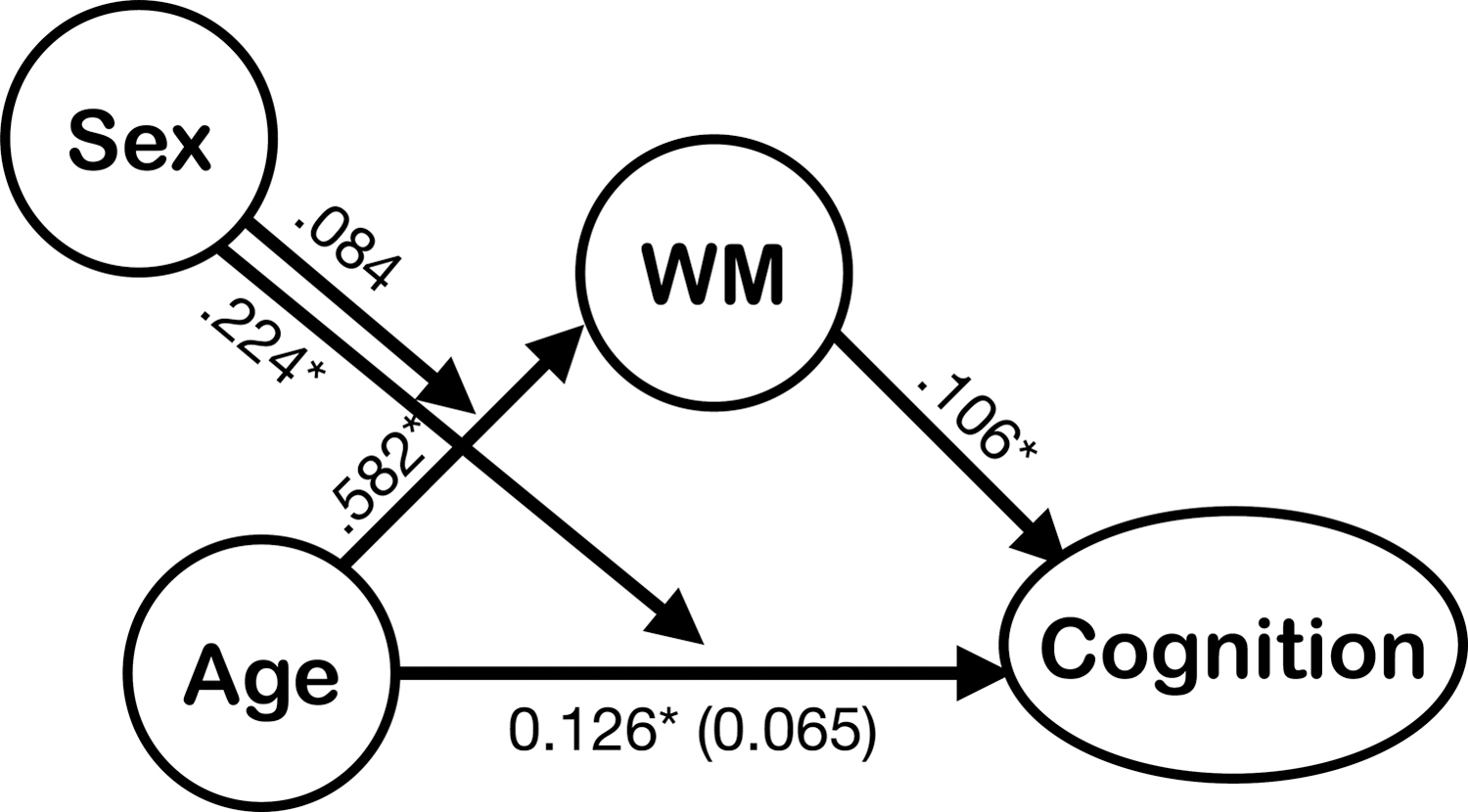
Moderated mediation model for the relationship between age and canonical cognitive z scores mediated by the canonical WM scores that were moderated by sex. The numbers above the arrow indicated the standardized regression coefficients. The indirect effects of males and females are, respectively, 0.0617 (CImale = [0.019–0.107]) and 0.0706 (CIfemale = [0.022–0.122]). After moderated mediation, the direct effect of males became insignificant (b = 0.065, p = 0.118). Females showed both indirect and direct effects (b = 288, p < 0.001) in this relationship. IE 95% CI, 95% confidence interval for the indirect effect with 10,000 bootstrapped samples; WM, white matter.
Discussion
We analyzed the cognitive decrease in four domains with advanced age from a large single-site life span data set, and discovered that females showed considerably faster deterioration than male participants in VS and E. The WM changes accelerated after age 40. These changes were revealed by the trajectories of AD and RD, which implied the degeneration in axon and myelin. Specifically, females had lower AD than males in the projection tracts (CST and ATR) and the association tracts (ILF, CgH, and tSLF) with a faster and more prominent decrease of RD. In contrast, Fmi, the anterior part of the commissural tract, was the only sex-linked tract that showed accelerated decrease in AD during aging in male participants. Those sex-linked tracts were closely associated with cognitive performances and partially mediated the cognitive aging process. Our results indicate that sex affects both cognitive function and WM microstructure, and further involves in the mediation of the age-WM-cognition relationship.
The gradual cognitive deterioration during aging was well-documented, however, there is a discrepancy of studies on sex effects in the cognitive decline. For example, Maitland and his colleagues reported that females have a steeper age-associated decline in verbal-based functions, while Lundervold's team demonstrated the opposite result (Lundervold et al., 2014; Maitland et al., 2004); McCarrey's group remarked that male participants showed a steeper rate of decline of performance in visuospatial ability with age; on the contrary, León found a pronounced decrease in females (León et al., 2016; McCarrey et al., 2016). Conducted with a larger sample size, our study demonstrated the age-related decrease across all cognitive domains, in which only females showed a steeper age-associated decrease in VS and E, corresponding to the predominant part in cognitive aging (Buckner, 2004; Fjell et al., 2016; Harada et al., 2013).
Previous histopathological studies had found several features that occurred with WM aging, including loss of myelinated fiber, malformation of the myelin sheath, and axonal damage (Adalbert and Coleman, 2013; Bartzokis, 2004; Pakkenberg and Gundersen, 1997; Peters and Sethares, 2002). DTI quantifies the diffusion in WM fibers and myelin sheath, and may be able to capture those microstructural changes in the brain. Previous studies on rats have indicated that demyelination may be revealed by the change of RD values, while axonal damage may be reflected by AD values (Song et al., 2002; Sun et al., 2006). FA measure has been shown to be insensitive to evaluate the WM microstructure during the aging process (Acosta-Cabronero et al., 2010; Alexander et al., 2007), while the AD and RD showed clear neural changes across the life span (Bennett et al., 2010; Kumar et al., 2013; Sexton et al., 2014; Winklewski et al., 2018; Wu et al., 2011). Relatively flat FA trajectories accompanied with change in RD or AD were observed, including ILF, CgH, tSLF, CST, and ATR (Cox et al., 2016; Sullivan et al., 2010). Current findings demonstrated that those sex-linked tracts in female not only had higher AD values, but also increased more quickly in RD, which indicated that the deterioration of WM was more severe in females.
Another important finding of this study was that the microstructural conditions of sex-linked tracts in females showed considerably greater mediation in the performance of VS and E. Association tracts, including ILF, tSLF, CgH, and ILF, were devoted to visual-related function (Latini, 2015; Voineskos et al., 2012); particularly, tSLF and CgH were responsible for improving the function of selective spatial attention and visuospatial working memory (Burks et al., 2017; Metzler-Baddeley et al., 2017). CST and ATR were the projection tracts that passed through and extended from the subcortical areas to the cerebral cortex, and integrated multiple functions across different cognitive domains (Benito-León et al., 2017; Moeller et al., 2015). Lesions and microstructural change in projection tracts were associated with the reduction in global cognitive function, E, processing speed, attention, and visual memory function (Cremers et al., 2016; Jiang et al., 2018; Mamiya et al., 2018; Smolker et al., 2018). More importantly, the degeneration in specific projection tracts may result in Alzheimer's disease (Douaud et al., 2011; Takahashi et al., 2002; Torso et al., 2015). In this study, the most significant sex effect was found in projection tracts, where tracts deteriorated more severely in females and had the closest association with decreased visuospatial and executive performances. This specific pattern may be associated with a higher incidence of Alzheimer's disease in females.
Compared with males, females had a larger indirect effect on the relationship among age, WM microstructure, and cognitive performances. Faster change in most WM tracts was revealed in females than males in this study. Previous postmortem studies demonstrated that males usually had more dense and larger sized axons in the brain, while female brains had more unmyelinated neuropil/neuronal process regions (Courten-Myers, 1999; Souma et al., 2008). In vitro studies also indicated that smaller axons and thinner myelin sheath in females may be more susceptible to age-related degradation (Bartzokis, 2004; Dollé et al., 2018). In contrast, overproduced and larger axons in male brains not only could compensate for the pathological neuronal loss but also could prevent accelerated aging (Rabinowicz et al., 1999).
Specifically, Fmi, the anterior part of the commissure tract for interhemispheric communications, was the only tract in males that showed a faster change in axons than that in females. Corresponding to a previous cross-sectional DTI study, it had also reported that males showed a significant increase in RD in genu but not splenium (Sullivan et al., 2010). It was evident that females had stronger interhemispheric connections and higher apparent fiber density in the corpus callosum, especially in the anterior part (Genc et al., 2018; Shiino et al., 2017). These may suggest that dense axonal fibers in Fmi may be one of the possible reasons for compensating the accelerated neuronal loss in females (Dollé et al., 2018; Rabinowicz et al., 1999).
Sex differences in the magnitude of diffusion measures have been constantly reported in WM-related studies (Menzler et al., 2011; Ritchie et al., 2018). On the contrary, the reports of sex differences in age-related change were mixed (Cox et al., 2016; Inano et al., 2011; Sullivan et al., 2010). Although the age-related patterns in this study were in line with both cross-sectional and longitudinal studies (de Groot et al., 2016; Sexton et al., 2014), we have to be cautious that some cross-sectional studies reported age and sex interaction (Hsu et al., 2008; Kochunov et al., 2012; Sullivan et al., 2010), and some others reported that the age/sex interaction was trivial or insignificant (Cox et al., 2016; Inano et al., 2011). With a longitudinal design, Sexton and colleagues (2014) had reported that there was no significant difference in annual change between males and females.
The major strengths of our study include large sample size, wide age range, and single MR site scanning, however, the caveat was that what we have observed may come from the period and cohort effect (Palmore, 1978). Including longitudinal data with multiple time points and a long interval may be a key area for future research.
We noticed that sex-linked tracts, tSLF, CgH were only shown in the left hemisphere in this study. A leftward asymmetry had been reported in arcuate fasciculus and cingulum in previous studies (Gong et al., 2005; Takao et al., 2011a). It has been reported that the asymmetric reduction was related to some development of neurodegenerative disorders (Irwin et al., 2018). A detailed study focus on sex effect in brain asymmetries during aging may be possible to provide insights into observing disease states.
There are some limitations. First, although the data set covered the entire adulthood and the aging process, the cognitive tasks were selectively administered for the first cohort (age older than 50 years, 819 participants were included). Therefore, the imaging/cognitive relationship can only be estimated after the age of 50. Second, the education level between males and females was different in these two data sets (see Supplementary Fig. S1 for details). To minimize the discrepancy, we have controlled the education level for evaluating the sex differences in visual function and E. However, the effect of the education may not be fully excluded. Third, the DTI protocol in this study only has limited directions and one fixed b value. Although DTI can be acquired and has been widely used to study brain WM, its capacity to capture the microstructure was limited (Jones et al., 2013). We have tried to interpret the aging pattern with full DTI measurements instead of reporting a single value. Fourth, although it is beneficial to have a large cross-sectional sample size on the same scanner, it is important to note that the cross-sectional study may not be identical to the age-related changes within individuals from longitudinal data. While the diffusion MRI technology and protocols advance rapidly, it would be necessary but also challenging to consider using higher order models on a longitudinal data set in a future study.
Conclusion
Sex has pronounced implications for the human brain throughout the life span, especially in the aging process. Both males and females aged in the same way, but the deterioration accelerated in the major projection tracts among females. The vulnerability of projection tracts in females during aging creates a negative impact on integrating multiple functions, which may be the key to a faster decrease in visuospatial and executive performance in females. By providing the aging trajectories of cognitive function and WM structure, we highlighted the importance of sex in the process of brain aging and its association with cognitive functions. Taking sex effect into account in the studies of the aging population is crucial for extended researches that are related to brain-based diseases and cognitive impairment in the aging process.
Footnotes
Acknowledgment
The authors acknowledged the support from the MRI Core of National Yang-Ming University in Taiwan.
Authors' Contributions
C.C.H. (Chih-Chin): study design and concept, MRI acquisition, image processing, data analysis and interpretation, bibliography research, and drafting of the article. C.C.H. (Chu-Chung): study design and concept, MRI acquisition, image processing, statistical analysis, data interpretation, and revising the article. S.-J.T., L.-K.C., and H.C.L.: study concept, revising the article, and approved the submitted version. C.-P.L. and C.-Y.Z.L.: study design and concept, data interpretation, bibliography research, critical revision of the article, and approved the submitted version.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported as follows. C.-Y.Z.L. was partially supported by the National Key Research and Development Program of China (No. 2018YFC0910503), the Young Scientists Fund of the National Natural Science Foundation of China (No. 81801774), and the Natural Science Foundation of Shanghai (No. 18ZR1403700). C.-P.L. was supported by the Ministry of Science and Technology (MOST) of Taiwan (MOST 110-2321-B-010-004, MOST 110-2321-B-010-007, MOST 110-2634-F-010-001, and MOST 108-2321-B-010-010-MY2). This work was supported, in part, by the Brain Research Center, National Yang-Ming University from The Featured Areas Research Center Program within the framework of the Higher Education Sprout Project funded by the Ministry of Education (MOE) in Taiwan.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2-1
Supplementary Table S2-2
Supplementary Table S2-3
Supplementary Table S2-4
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.