
Abstract
Background:
Brain interdependencies can be studied from either a structural/anatomical perspective (“structural connectivity”) or by considering statistical interdependencies (“functional connectivity” [FC]). Interestingly, while structural connectivity is by definition pairwise (white-matter fibers project from one region to another), FC is not. However, most FC analyses only focus on pairwise statistics and they neglect higher order interactions. A promising tool to study high-order interdependencies is the recently proposed O-Information, which can quantify the intrinsic statistical synergy and the redundancy in groups of three or more interacting variables.
Methods:
We analyzed functional magnetic resonance imaging (fMRI) data obtained at rest from 164 healthy subjects with ages ranging in 10 to 80 years and used O-Information to investigate how high-order statistical interdependencies are affected by age.
Results:
Older participants (from 60 to 80 years old) exhibited a higher predominance of redundant dependencies compared with younger participants, an effect that seems to be pervasive as it is evident for all orders of interaction. In addition, while there is strong heterogeneity across brain regions, we found a “redundancy core” constituted by the prefrontal and motor cortices in which redundancy was evident at all the interaction orders studied.
Discussion:
High-order interdependencies in fMRI data reveal a dominant redundancy in functions such as working memory, executive, and motor functions. Our methodology can be used for a broad range of applications, and the corresponding code is freely available.
Impact statement
Past research has showcased multiple changes to the brain's structural and functional properties caused by aging. Here we expand prior work through recent advancements in multivariate information theory, which provide richer and more theoretically principled analyses than existing alternatives. We show that the brains of older participants contain more redundant information at multiple spatial scales—that is, activation in different brain regions is less diverse, compared with younger participants—and identify a “redundancy core” constituted by prefrontal and motor cortices, which might explained impaired performance in the old population in functions such as working memory and executive control.
Introduction
A
Age-related cognitive decline occurs in parallel with well-established variations in brain morphology. During an individual's life span, the total brain volume increases from childhood to adolescence by about 25% on average, remaining constant over the following three decades and, ultimately, decaying toward childhood sizes at later ages (Courchesne et al., 2000). Notably, the atrophy in aged brains is not uniform, but rather some anatomical regions are more strongly affected than others, with the hippocampus (West, 1993) and the prefrontal cortex (Sullivan and Pfefferbaum, 2007) structures known to be targeted for age-related atrophy.
Resting-state functional magnetic resonance imaging or functional MRI (fMRI) has shown that aging affects functional connectivity (FC) in extended brain networks, specifically altering anterior and posterior regions such as the superior and middle frontal gyrus (MFG), posterior cingulate, middle temporal gyrus, and the superior parietal region (Andrews-Hanna et al., 2007; Damoiseaux et al., 2008). Other studies have shown that aging may also alter the FC between several regions, possibly indicating compensation or pathological activation (Betzel et al., 2014; Chan et al., 2014; Onoda et al., 2012; Solesio-Jofre et al., 2014).
An important limitation of such FC studies is that their analysis is restricted to pairwise FC, ignoring possible higher order effects. High-order interactions allow us to distinguish redundancy- and synergy-dominated interactions that play key roles in neural dynamics (Battiston et al., 2020; Erramuzpe et al., 2015; Friston and Price, 2003; Ince, 2017; Ince et al., 2017; Schneidman et al., 2003; Stramaglia et al., 2016, 2021). An initial study of high-order interactions and aging showed significant changes in the synergies and redundancies of triple interactions over the individual's life span (Camino-Pontes et al., 2018), as well as a redundant role of the default mode network (DMN). However, the effects of aging on interactions beyond triple relationships remain largely unexplored to the best of our knowledge.
Here, we built on our previous studies (Camino-Pontes et al., 2018) to examine the effects of aging on higher order interactions in the human brain, paying special attention to the interdependencies between four or more brain regions. We used the recently proposed O-Information (Rosas et al., 2019), which can be considered a revised version of the measure of neural complexity proposed previously in the context of partial information decomposition (Tononi et al., 1994; Williams and Beer, 2010). More specifically, O-Information captures the balance between redundancies and synergies in arbitrary sets of variables, thereby extending the properties of the interaction information of three variables to larger sets (McGill, 1954). Redundancy is understood here as an extension of the conventional notion of the correlation between more than two variables, in which each variable has a “copy” of some common information shared with other variables (Rosas et al., 2018). An example of extreme redundancy is full synchronization, where the status of one signal allows one to predict the status of any other. By contrast, synergy corresponds to statistical relationships that regulate the whole but not the parts (Rosas et al., 2019; Timme et al., 2014; Wibral et al., 2017). Synergy allows local independency and global cohesion to coexist, a condition that has been recently found to be instrumental for higher order brain functions (Luppi et. al, 2020a,b), while redundancy—including highly synchronized situations such as deep sleep stages or epileptic seizures—would make brains less well-suited to this (Tononi and Edelman, 1998; Tononi et al., 1994).
To investigate how the higher order informational organization of the brain changes with age, we studied synergetic and redundant interactions of different orders from fMRI data obtained in the resting state from a cohort of 164 healthy volunteers aged 10 to 80 years. This approach is novel for two reasons. From a methodological perspective, we present the first application of higher order statistical interdependencies in aging, beyond triplet interactions, developing a formalism to apply the metrics previously presented to fMRI data (Rosas et al., 2019). In addition, we investigate how aging alters the higher order interdependencies among different brain regions, as inferred directly from fMRI data. In this way, we obtained novel insights into brain aging, through which the interdependencies in older participants appear to be more redundancy-dominated than those in younger participants, for all interaction orders. When studying how these effects are distributed topographically, we found a “redundancy core” that was composed of brain regions that play key roles in the most redundancy-dominated arrangements for all interaction orders.
Materials and Methods
Participants
A cohort of healthy participants (p = 164) were recruited in this study, aged from 10 to 80 years (mean age 44.35 years, standard deviation 22.14 years). Informed consent was obtained from all the subjects before they were enrolled onto the study. The experimental protocol was approved by the Ethics Committee Research UZ/KU Leuven (project S51615) and was conducted according to the Declaration of Helsinki (1964) and its amendments (World Medical Association 2013).
We grouped the participants into four age groups Ii for i ϵ {1, …, 4}, more specifically: I1 consists of 30 subjects aged 10–20 years; I2, 46 subjects 20–40 years; I3, 29 subjects 40–60 years; and I4 59 subjects 60–80 years.
Image acquisition and analyses
MRI images were acquired with a Siemens 3T MAGNETOM Trio MRI scanner using a 12-channel matrix head coil. The anatomic data were acquired as a high-resolution T1 image with 3D magnetization-prepared rapid acquisition gradient echo (MP-RAGE): repetition time = 2300 ms, echo time = 2.98 ms, voxel size = 1 × 1 × 1.1 mm3, slice thickness = 1.1 mm, field of view = 256 × 240 mm2, with 160 contiguous sagittal slices covering the entire brain and brainstem. Resting-state functional data were acquired with a gradient echo-planar imaging sequence over a 10-min session using the following parameters: 200 whole-brain volumes with TR/TE = 3000/30 ms, flip angle = 90, interslice gap = 0.28 mm, voxel size = 2.5 × 3 × 2.5 mm3 80 × 80 matrix, slice thickness = 2.8 mm, 50 oblique axial slices interleaved in descending order.
Image preprocessing was performed following a similar procedure to that used in Camino-Pontes and colleagues (2018) and subsequently, each subject was represented by 2514 time series of the fMRI signal (Fig. 1). These series were grouped into 20 new time series by spatial-averaging, corresponding to the M = 20 module partition of the Brain Hierarchical Atlas (Diez et al., 2015), the partition that maximizes functional modularity, structural modularity, and the similarity between the functional and structural modules (Diez et al., 2015).
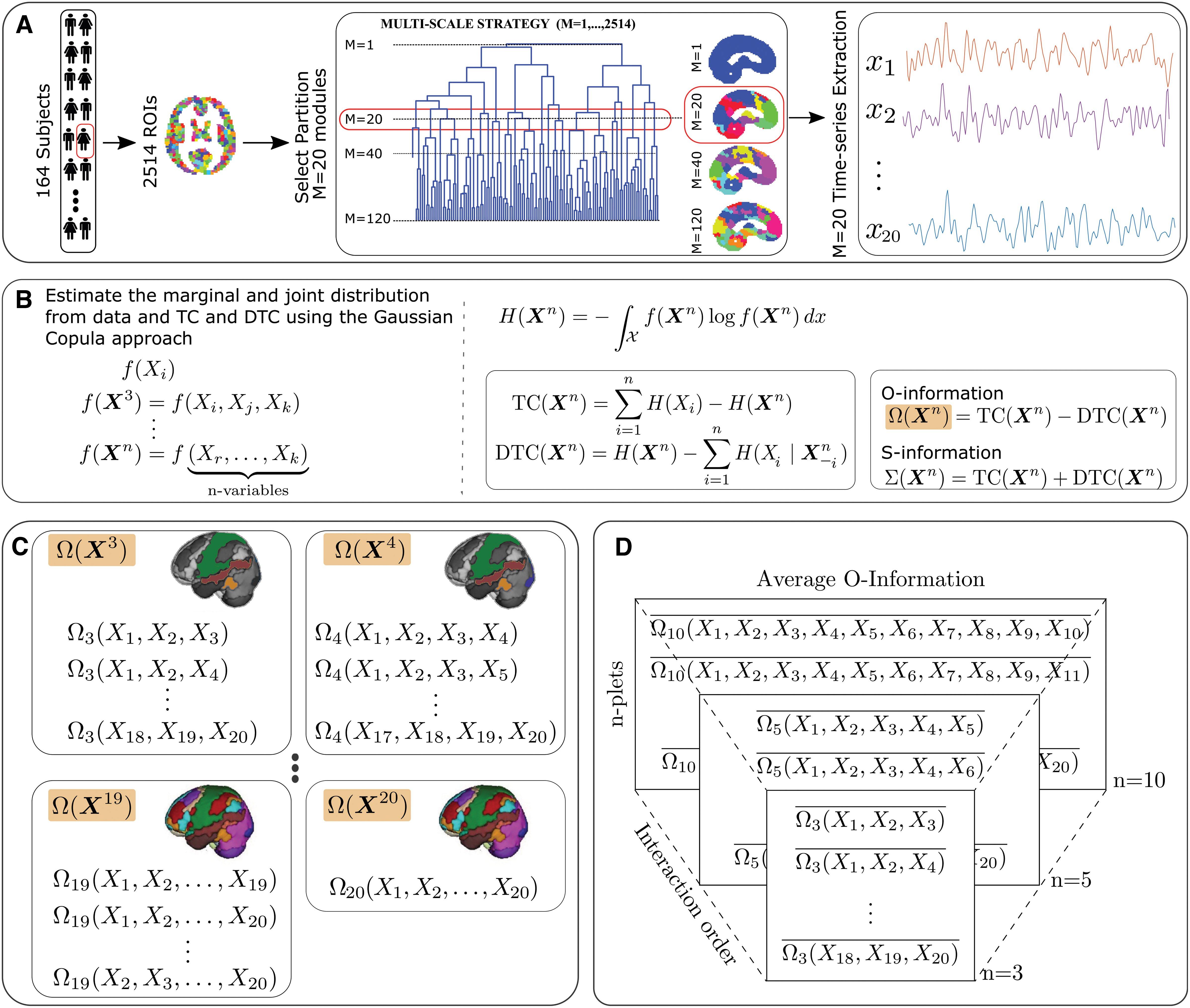
Methodological scheme.
Multivariate information-theoretic metrics
O-Information (Rosas et al., 2019) (shorthand for information about organizational structure) is an attempt to operationalize the original desiderata (Tononi et al., 1994) while overcoming some of the shortcomings of this original proposal (Van Putten and Stam, 2001; van Walsum et al., 2003). Denoted by
In addition to O-Information, we also assessed the S-information (Rosas et al., 2019) denoted by Σ(X n ), to quantify the strength of multivariate correlations. S-Information is a natural complement to O-Information as it quantifies the overall strength of the interdependencies, while the latter determines whether they are synergistic or redundant in nature. Furthermore, following Camino-Pontes and colleagues (2018), we take the positive and negative parts of O-Information as heuristic metrics of redundancy and synergy.
Below, we provide details regarding the main information-theoretic metrics used in this study (Rosas et al., 2019). The O-Information of a set of n random variables
while their S-Information is
In these formulae, TC and DTC correspond to the total correlation (Watanabe, 1960) and dual total correlation (Han, 1978), respectively, given by the following:
Here,
All these quantities were computed for each subject separately using Gaussian Copulas
i
(Ince et al., 2017). The O-Information and S-Information estimated via the data of the kth subject are denoted as
Interaction order, redundancy, and synergy
To estimate the O-Information and S-Information of different brain modules, we calculated the following quantities for each participant:
Above, k is the participant index, m is the module index, n interaction order, and
is the total number of subsets of size
We then split the values of O-Information on positive and negative values using
so that
where
Statistical analyses
In this study, we compared the group of older participants (I4) with the combination of the three other groups (I1, I2, I3). Different information-based measures were compared using a nonparametric statistical Wilcoxon rank-sum test. When appropriate, significance levels for hypothesis testing were corrected for multiple comparisons by controlling the false discovery rate (FDR) following a standard Benjamini/Hochberg procedure (Benjamini and Hochberg, 1995).
Results
We analyzed resting-state fMRI data from 164 healthy subjects, measuring the S-Information and O-Information to quantify high-order interactions between brain areas in terms of their redundancy and synergy. A scheme of the approach adopted is shown in Figure 1. Participants were divided into four age groups (I1–I4) as described in the Materials and Methods section.
In this study, we first measured the S-Information and O-Information per age group, and the interaction order. Significant differences were evident in both the S-Information and O-Information between the old group and the younger groups after correcting with the FDR for multiple comparisons (Fig. 2). The increase in S-Information with age implies an increase of interdependencies between the various brain regions. Interestingly, the increase shown in the older population was significant at all orders, suggesting a widespread effect. The increase in the O-Information detected suggests that the correlations in the older population are generally redundancy-dominated, becoming stronger at higher orders. We split the interactions according to the sign of the O-Information, with the positive values representing a measure of redundancy, and the negative ones, of synergy. Analyzing the O-Information, we identified different patterns of redundancy and synergy associated with increasing interaction order (Fig. 3). While synergy exhibited a bell-shaped curve, redundancy increased linearly with the interaction order. Importantly, the redundancy values were much larger than the synergistic ones, and moreover, the redundancy of I4 showed significant differences from that in the group formed by I1, I2, and I3 for all interaction orders. In terms of synergy, while for some interaction orders the I4 group exhibited significant differences from the rest of the population, these differences did not survive a multiple comparison correction.
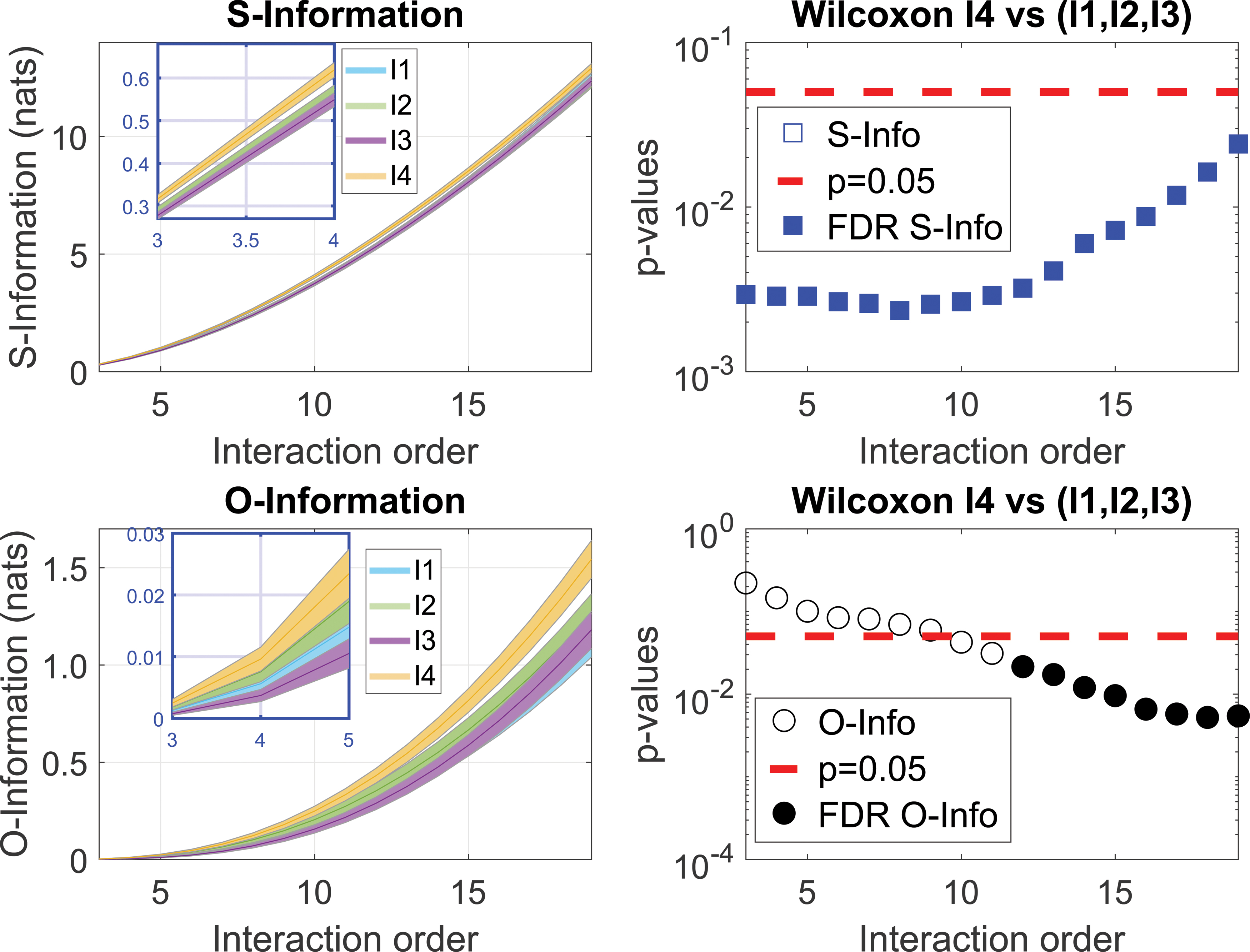
Strong high-order interdependencies in the aging brain. O-Information and S-Information average over all modules
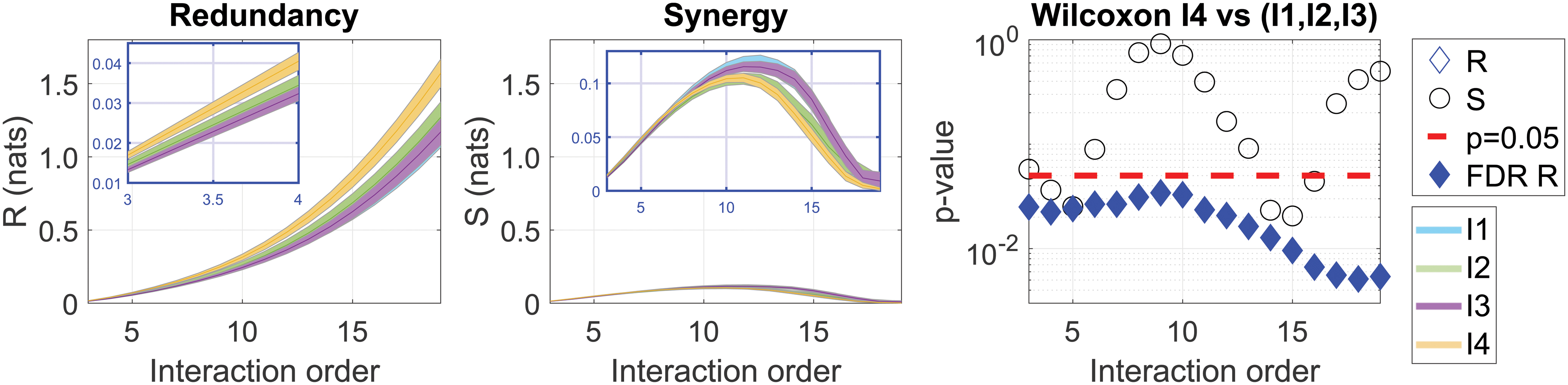
A significant increase in redundancy in older participants across all interaction orders. The average redundancy and synergy were obtained over modules
We quantified the redundancy across brain modules by taking into account all the redundant interactions in which a given module participates. When studying redundancy across the distinct brain areas, modules 1–3, 5, 13–15, 18–20 exhibited significant differences for all interaction orders, while the others only exhibited significant differences for large interaction orders (Fig. 4). This suggests the existence of a redundancy core, as confirmed by later analyses. By contrast, the pattern of synergy across brain areas is very heterogeneous, with only modules 15, 17, 18, and 19 showing significant differences.

Higher order redundant and synergistic interdependencies across brain areas in the aging brain. Differences between I4 and the I1, I2, I3 groups across the different brain modules, each one anatomically represented by three representative views from left to right, sagittal, coronal, and axial. As in Figure 3, group differences in redundancy (diamonds) and synergy (circles) are represented as a function of the interaction order. When the group differences survived correction for multiple comparisons, both the diamonds and circles are filled. In relation to redundancy, there are two “classes” of modules, those where redundancy was significantly different for any order (such as modules 1–3, 5, 13–15, and 18–20) and the remaining modules for which this did not happen. Moreover, redundancy generally differs widely between group I4 and the rest of the population across the different brain areas, yet synergy does not, for which only modules 15, 17, 18, and 19 showed a few interaction orders with values surviving multiple comparisons. Color images are available online.
To confirm the existence of a redundancy core, we studied the extreme O-Information values for various interaction orders. We found that modules 2, 5, and 13 participate in the most redundancy-dominated n-plets for all orders (Fig. 5), suggesting that they might constitute the basis of such a core. While a complete anatomical description of these modules is available (Diez et al., 2015, 2017), it is important to emphasize that these three modules have the MFG and precentral gyrus (PG) in common, two important structures that have a well-differentiated function. Specifically, while the MFG is part of the prefrontal cortex, which mediates executive control and working memory, the PG is part of the primary motor cortex, both structures known to be affected by aging (Grady, 2008, 2012; Pauwels et al., 2019; Salat et al., 2004).
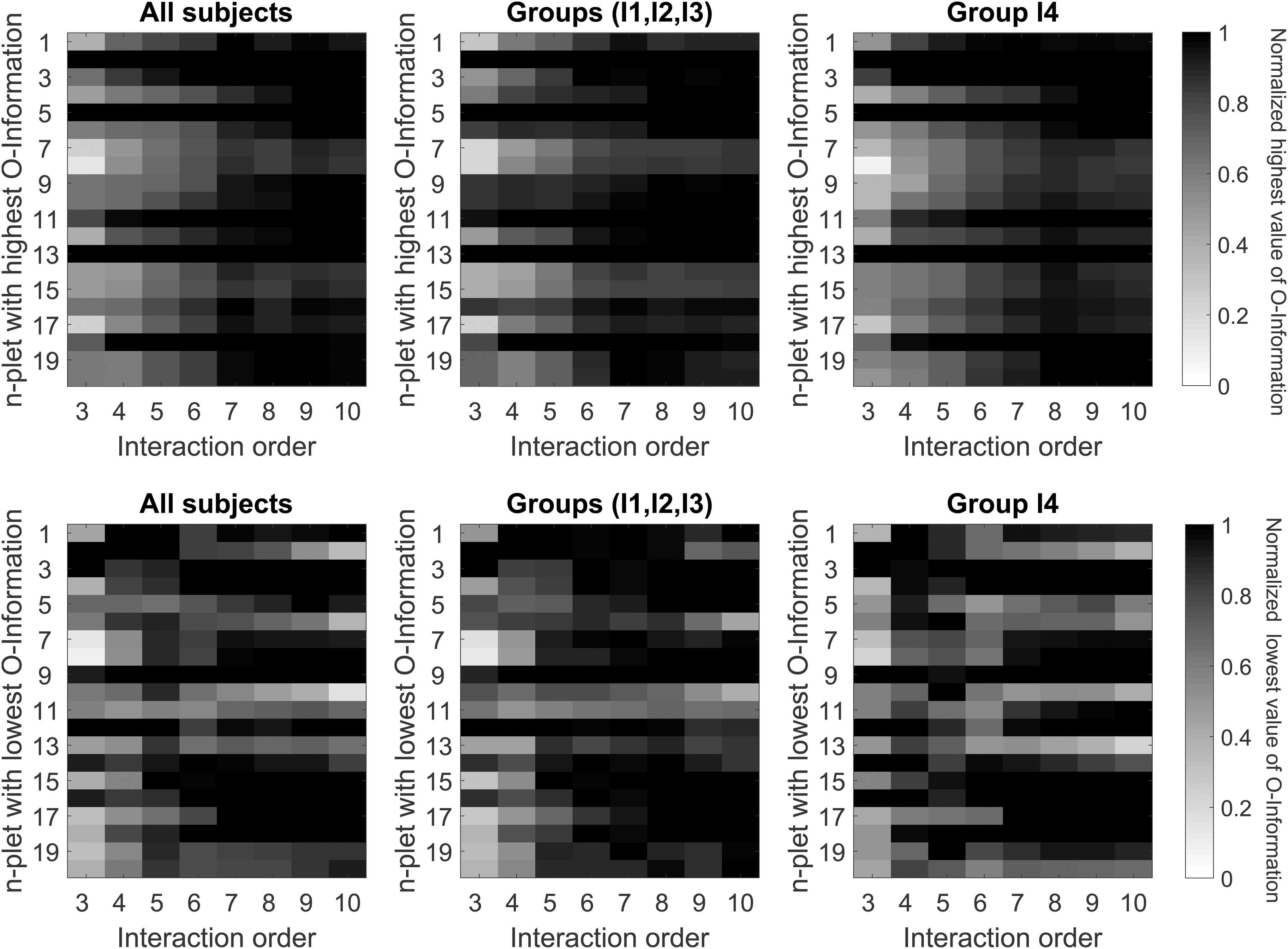
Identification of the redundant and synergistic cores for the different interaction orders. Top row: As a function of the interaction order n, we rank the averaged O-Information values of all the n-plets per group of participants, as detailed in the Materials and Methods section and illustrated in Figure 1D. We plotted the highest O-Information value in which each module participates for each interaction order, normalized to the n-plet with the highest O-Information averaged in different groups of participants. For each interaction order, the n-plets with the highest values of O-Information are in black. The highest O-Information values measure redundancy. Bottom row: As above but plotting the lowest values of O-Information. Because all were negative, they corresponded to synergy. The two rows show similar plots for three different situations: all subjects (left); pooling I1, I2, and I3 together (middle); and for the older group, I4 (right).
It is important to note that although the redundant role found in this study for the interaction between the MFG and PG was obtained from participants at rest, it potentially reflects the fact that older participants typically compensate motor behavioral deficits by additional activation of the prefrontal cortex in synchrony with their associated motor areas, while younger participants only activate motor areas to perform the same tasks (Heuninckx et al., 2008; Rueda-Delgado et al., 2019; Santos-Monteiro et al., 2017). Similarly, it has also been shown that older but not young subjects recruit the prefrontal cortex when performing purely movement tasks, such as inhibitory motor control, thereby relying more on cognitive support for the performance of a motor task, while younger subjects perform such tasks in a more automatic manner (i.e., cognitive penetration into action) (Heuninckx et al., 2005).
Interestingly, the core existing in younger participants (I1, I2 and I3) seemed to be broader than that in the older population, including modules 9, 10, and 16. The brain structures supporting these modules are the middle and superior frontal gyri, posterior cingulate cortex, and the precuneus, all part of the DMN, an important network in the human brain (see Buckner et al., 2008; Raichle, 2015, and references therein). Although a weaker contribution of the precuneus to the DMN has been proposed (Margulies et al., 2009), our data confirmed its participation, as indicated elsewhere (Beckmann et al., 2005; Raichle, 2015).
When comparing these results with those in which a redundant role of the DMN was shown over the individual's life span when
When the modules that participate in synergistic arrangements are considered, that is, in the n-plets with smaller O-Information values, those belonging to the redundant core tended not to be involved in very synergistic arrangements, with the exception of modules 9 and 16 that participated in all age groups. These two modules have the posterior cingulate and precuneus in common, which as indicated above are part of the DMN. Based on these results, we might hypothesize a dual redundant/synergistic role of the DMN, and an impairment of this redundant contribution in older participants.
Conclusions
The present study assesses the high-order redundant and synergistic interactions among brain regions in subjects of various ages. Overall, there was an important increase in the redundant interdependencies in the older population for all interaction orders. In addition, a redundant core of brain modules was observed, the influence of which diminished with age. Together, these findings suggest a change in the balance of differentiation and integration toward more synchronized arrangements.
It is important to note that our analysis goes beyond the traditional brain-network approaches that focus on pairwise interactions, considering higher order interactions that can assess redundant and synergistic effects. In doing so, we follow the seminal ideas introduced previously (Tononi et al., 1994), which postulate that higher brain functions might depend on the coexistence of integration and segregation. Indeed, while the latter enables brain areas to perform specialized tasks independently of each other, the former serves to integrate brain areas to perform goal-directed tasks. A key insight put forward in Tononi and colleagues (1994) is that segregation and integration can coexist, and that this coexistence is measurable by assessing the high-order interactions of neural elements via approaches such as the one used in this study.
In the context of aging, it has been shown that the balanced segregation/integration might breakdown as internetwork connectivity augments in older individuals, thereby compromising segregation. However, this increase is not associated with improved performance, indicating that the limited segregation is related to neuronal dysfunction, possibly due to the weaker inhibitory activity in older adults (King et al., 2018).
It is worth noting that the increased redundancy detected in aging cannot be cast in terms of pairwise interactions. Redundancy not only implies that two areas share information but also that the shared information is the same. Notably, no pairwise network approach can discriminate this relationship. Therefore, more redundancy in the brain does not merely imply higher connectivity, but rather that the same information is present in more than two places at the same time (the number depending on order of redundancy), which in turn suggests a loss of specificity and potentially the suboptimal utilization of resources.
Analyzing the O-Information, we identified different patterns of redundancy and synergy associated with increasing interaction order (n). While synergy exhibited a bell-shaped curve, redundancy increased linearly with n. It is also important to emphasize that the two metrics R and S are built in a similar way, and therefore, the different scaling behavior when increasing n might be due to the differences in the nature of interactions separately for R and S. Although further work is needed to fully clarify the tendency when systematically increasing
The current study is based on the following methodological limitations. First, that the accurate quantification of redundancy in large-scale brain networks remains unresolved (Lizier et al., 2018), although by using the O-information measure to reflect the balance of synergy/redundancy we propose a different metric as a proxy to estimate synergy and redundancy. Second, different brain parcellations can be used to explore higher order functional interactions in the brain, although a finer spatial resolution compromises the calculation of all of the n-plets when assessing the O-Information when the number of combinations become very large. Third, the fact that the O-Information values themselves suggest a predominance of redundancy might be heavily influenced by the nature of the fMRI data, and different results might be obtained if other imaging modalities were used (e.g., electroencephalography or magnetoencephalography). Nevertheless, the increase in O-Information is a clear evidence of a change in the differentiation/integration balance toward more redundant arrangements. Finally, in the context of aging, we have shown major variations of R and S across brain structures occurring in the old population by taking into consideration exclusively fMRI. How much of these variations due to age in R and S might be originated by age variations in the structural connectivity matrix is an unexplored territory that needs to be addressed in future work.
In summary, the framework presented here provides a novel insight into the aging brain by assessing high-order functional interdependencies among brain regions. The data reveal the role of redundancy in prefrontal and motor cortices in older participants, which affects basic functions such as working memory, or executive and motor functions. We believe that this approach may help to provide a better understanding of some brain disorders from an informational perspective, providing distinctive patterns of higher order functional behavior or “info-markers,” which may offer fundamental insights into the healthy and diseased human brain. The code to compute the metrics used in this article has been made publicly available at
Footnotes
Acknowledgments
The authors thank Karine Bertin for her valuable comments on our work. M.G. was partially supported by CONICYT-PFCHA/Doctorado Nacional/2019-21190577. R.C. acknowledges financial support from Fondecyt Iniciación 2018 Proyecto 11181072. P.M. was funded by the Wellcome Trust (Grant No. 210920/Z/18/Z). F.R. acknowledges the support of the Ad Astra Chandaria Foundation. P.O. is funded by Fondecyt Regular grant 1181076 and ANID-Basal Project FB0008. S.P.S. was supported by the FWO Research Foundation Flanders (G089818N), the Excellence of Science funding competition (EOS; 30446199), and the KU Leuven Special Research Fund (grant C16/15/070). J.M.C. acknowledges financial support from Ikerbasque (The Basque Foundation for Science) and from the Ministerio Economia, Industria y Competitividad (Spain) and FEDER (Grant DPI2016-79874-R), as well as from the Department of Economic Development and Infrastructures of the Basque Country (Elkartek Program, KK-2018/00032 and KK-2018/00090). The Centro Interdisciplinario de Neurociencia de Valparaíso (CINV) is a Millennium Institute supported by Grant ICN09-022 (ICM-ANID).
Preprint posted on bioRxiv. Date posted: March 18, 2020 id: 2020.03.17.995886.
Authors' Contributions
M.G.: study conceptualization, methodology, performed the analyses, produced the figures, and drafted the article. R.C.: study conceptualization, methodology, performed the analyses, supervised the information metrics, and drafted the article. P.A.M.M.: study conceptualization, methodology, supervised the information metrics, and drafted the article. F.E.R.: study conceptualization, methodology, supervised the information metrics, and drafted the article. P.O.: study conceptualization and methodology. I.D.: preprocessed the images. S.P.S.: recruited the participants and drafted the article. J.M.C.: study conceptualization, data curation, methodology, supervised the information metrics, and drafted the article. All the authors participated in the writing of the article and agreed on the submission of the final version.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Funding was provided by the following: Agencia Nacional de Investigación y Desarrollo (CONICYT-PFCHA/Doctorado Nacional/2019-21190577), Fondo Nacional de Desarrollo Científico y Tecnológico del Gobierno de Chile (Iniciación 11181072), Wellcome Trust (210920/Z/18/Z), Fondo Nacional de Desarrollo Científico y Tecnológico del Gobierno de Chile (Regular 1181076), Agencia Nacional de Investigación y Desarrollo del Globierno de Chile (Fondo Basal FB0008), Research Foundation Flanders (G089818N), Excellence of Science funding competition (EOS, 30446199), Katholieke Universiteit Leuven Special Research Fund (C16/15/070), Ministerio de Ciencia e Innovación (Gobierno de España) y Fondo Europeo de Desarrollo Regional (DPI2016-79874-R), Department of Economic Development and Infrastructures of the Basque Country (Elkartek KK-2018/00032, KK-2018/00090), Agencia Nacional de Investigación y Desarrollo Milenio del Gobierno de Chile (Instituto Milenio ICN09-022).