
Abstract
Introduction:
Synchronized oscillatory brain activity is considered a basis for flexible neuronal network communication. However, the causal role of inter-regional oscillatory phase relations in modulating signaling efficacy in cortical networks has not been directly demonstrated in humans so far.
Aim:
The current study addresses the causal role of transcranial alternating current stimulation (tACS)-induced oscillatory cross-network phase relations in modulating signaling efficacy across human cortical networks.
Methods:
To this end, concurrent tACS, transcranial magnetic stimulation (TMS), and electroencephalography (EEG) were employed to measure the modulation of excitability and signaling efficacy across cortical networks during externally induced neural oscillations. Theta oscillatory activity was introduced through tACS in two nodes of the human frontoparietal network: the dorsolateral prefrontal cortex (DLPFC) and the posterior parietal cortex (PPC). Six Hertz tACS was applied to the DLPFC and PPC simultaneously in an in-phase or antiphase manner. In addition, single-pulse TMS was administered over the DLPFC at four different phases of tACS and the propagation of TMS-evoked neuronal activity was measured with EEG.
Results:
We show that tACS-induced theta oscillations modulate TMS-evoked potentials (TEPs) in a phase-dependent manner, and that the induced oscillatory phase relation across the frontoparietal network affects the propagation of phase-dependent TEPs within as well as beyond the frontoparietal network.
Conclusion:
We show that the effect of tACS-induced phase relation across the frontoparietal network on signal transmission extends beyond the frontoparietal network. The results support a causal role of inter-nodal oscillatory phase synchrony in routing cortico-cortical information flow.
Impact statement
Theoretical models have proposed that phase relations of cross-network neural oscillations control communication efficacy across human cortical networks. The current study introduced concurrent transcranial alternating current stimulation-transcranial magnetic stimulation-electroencephalography (tACS-TMS-EEG) to experimentally study the theoretical framework. Dual-site in-phase or antiphase 6 Hz tACS was applied to the frontoparietal network. Synchronized tACS was shown to affect signaling within as well as beyond the targeted network. The study demonstrates how inter-regional oscillatory coherence supports the control of brain network signaling.
Introduction
Understanding the mechanisms of flexible neuronal information transfer across distributed brain networks is indispensable for understanding cognition. Neuronal oscillations have been considered a key candidate for enabling flexible routing of neuronal information flow in brain networks (Buzsáki and Draguhn, 2004; Engel et al., 2001; Fries, 2005; Varela et al., 2001). In recent years, new approaches to non-invasively modulate oscillatory activity in humans (Herrmann et al., 2015; Thut et al., 2011) have enabled us to directly experimentally address the role of oscillatory brain activity in neuronal communication and cognition. Transcranial alternating current stimulation (tACS) is one such promising approach, involving a weak alternating (sinusoidal) current being non-invasively applied through scalp electrodes (Paulus, 2011). tACS can modulate the excitability of the underlying neuronal tissue and drive neuronal population-level excitability fluctuations of a specific frequency band, and it allows to address their causal role in neuronal functioning and cognitive operations (Helfrich et al., 2014a,b; Kanai et al., 2008; Neuling et al., 2013; Polanía et al., 2012; Zaehle et al., 2010).
A prominent model of the role of neuronal oscillatory activity in neuronal communication (Fries, 2005, 2015; Varela et al., 2001; Womelsdorf et al., 2007) states that oscillations in neuronal excitability dictate responsiveness to neural inputs, and that oscillatory activity hence modulates the efficacy of neuronal communication in a periodic manner. Thereby, the dynamically established relation of the oscillatory phase across brain networks is proposed to gate signals propagating across the networks, flexibly rendering the inter-nodal communication more or less efficient. The modulation of oscillatory inter-nodal phase relations in humans via tACS has resulted in behavioral effects that suggest alterations in signaling efficacy across the targeted network nodes (Alekseichuk et al., 2017; Bächinger et al., 2017; Helfrich et al., 2014a; Polanía et al., 2012, 2015; Strüber et al., 2014; Tseng et al., 2018; Violante et al., 2017). Further, compared with antiphase tACS, in-phase tACS on the frontoparietal network has been shown to increase the correlation of fMRI signals across the network (Violante et al., 2017). However, an influence of oscillatory inter-regional phase relations on neural transmission has not been directly demonstrated in human brain networks so far.
With the aim of providing causal evidence for the role of oscillatory inter-regional phase relations in inter-regional neural communication in humans, we employed our recently introduced concurrent transcranial alternating current stimulation-transcranial magnetic stimulation-electroencephalography (tACS-TMS-EEG) method (Fehér et al., 2017). The rationale of the method is as follows. While introducing oscillatory activity with tACS, tACS-dependent modulation of neural transmission in cortical networks is measured as TMS-evoked potentials (TEPs) in the EEG. We have demonstrated that 6 Hz tACS modulates TMS-induced neural transmission in a phase-dependent manner (Fehér et al., 2017). In this way, we can address the causal influence of oscillatory neural dynamics on signal transmission.
In the present study, we targeted theta-frequency (6 Hz) phase relations between two nodes of the frontoparietal network: the dorsolateral prefrontal cortex (DLPFC) and posterior parietal cortex (PPC). Long-range functional connectivity across the frontoparietal networks, serving as a basis for higher-order cognitive processing (Womelsdorf and Everling, 2015), has been shown to be mediated by theta activity during sustained attention (Sellers et al., 2016) as well as working memory tasks (Alekseichuk et al., 2017; Polanía et al., 2012; Sauseng et al., 2005; Violante et al., 2017). Further, cortical theta oscillations have been shown to be generally associated with long-range cortico-cortical interactions (Cohen, 2014; Mizuhara and Yamaguchi, 2007; Narayanan et al., 2013). As EEG potentials are considered to reflect the synaptic inputs to a neuronal population, rather than the firing outputs of the neuronal population, TEPs observed in the parietal area would reflect inputs from the DLPFC (Buzsáki et al., 2012). Therefore, resulting changes due to tACS-controlled oscillatory phase relations in the frontoparietal network could be observed as changes in TEPs beyond the frontoparietal network. We, therefore, hypothesized that the spread of phase-dependent TEPs to brain areas connected to the left parietal cortex would depend on the induced theta-band phase relation between the frontoparietal nodes, in agreement with the proposed role of oscillatory phase coherence in modulating inter-nodal information flow.
Materials and Methods
Subjects
Twenty-four healthy human subjects participated in the experiment after providing written informed consent (16 females, mean age across subjects 24.1; range 19–38; all subjects right-handed). Three subjects were excluded due to problems during the recording. Two additional subjects were excluded due to insufficient data quality. The reasons for rejecting their data were the presence of residual tACS-induced and/or TMS-induced artifacts. The insufficient removal of tACS-induced artifacts was caused by excessive blinking artifacts, muscle artifacts, or a combination of these. Finally, data from 19 subjects were subjected to further analysis (11 females, mean age across subjects 24.5; range 20–38). Experimental procedures conformed to the Declaration of Helsinki and were approved by the ethics committee of Canton Bern (KEK-BE 007/14). All participants were screened for contraindications to TMS (Rossi et al., 2011), as well as for a previous history of mental illness or neurological disorders before enrolment.
Concurrent tACS-TMS-EEG recording
Single-pulse TMS with a biphasic pulse configuration was administered with a figure-eight-shaped coil with an outer diameter of 75 mm (MCF-B65 Butterfly Coil; MagVenture A/S) connected to a MagPro R30 magnetic stimulator (MagVenture A/S). The intensity of single-pulse TMS during the concurrent tACS-TMS-EEG recordings was set to 40% of the maximal output intensity of the stimulator. This corresponded to an average of 78.5% of the active motor threshold, as measured by finger twitch before the main measurements. Subjects wore the EEG cap during the measurement of the individual motor threshold to be consistent with the additional space between the coil and the scalp caused by the EEG cap in the experiment proper.
During the concurrent tACS-TMS-EEG recordings, the TMS coil was positioned tangentially to the scalp, centered above electrode F3 [which corresponds to the middle frontal gyrus (Koessler et al., 2009)] at an angle of 45° to the sagittal plane (Fig. 1B). Electrophysiological data were obtained via a 32 Ag/AgCl electrode cap (waveguard; ANT Neuro), recorded with a 24-bit EEG amplifier (eego sports; ANT Neuro), and digitized at a sampling rate of 2048 Hz. The recording reference was located at CPz, and the ground electrode was located at AFz. The EEG electrodes were placed according to the international 10–20 system for EEG electrode positioning (Jasper, 1958). Skin resistance at EEG electrodes was kept below 5 kΩ.
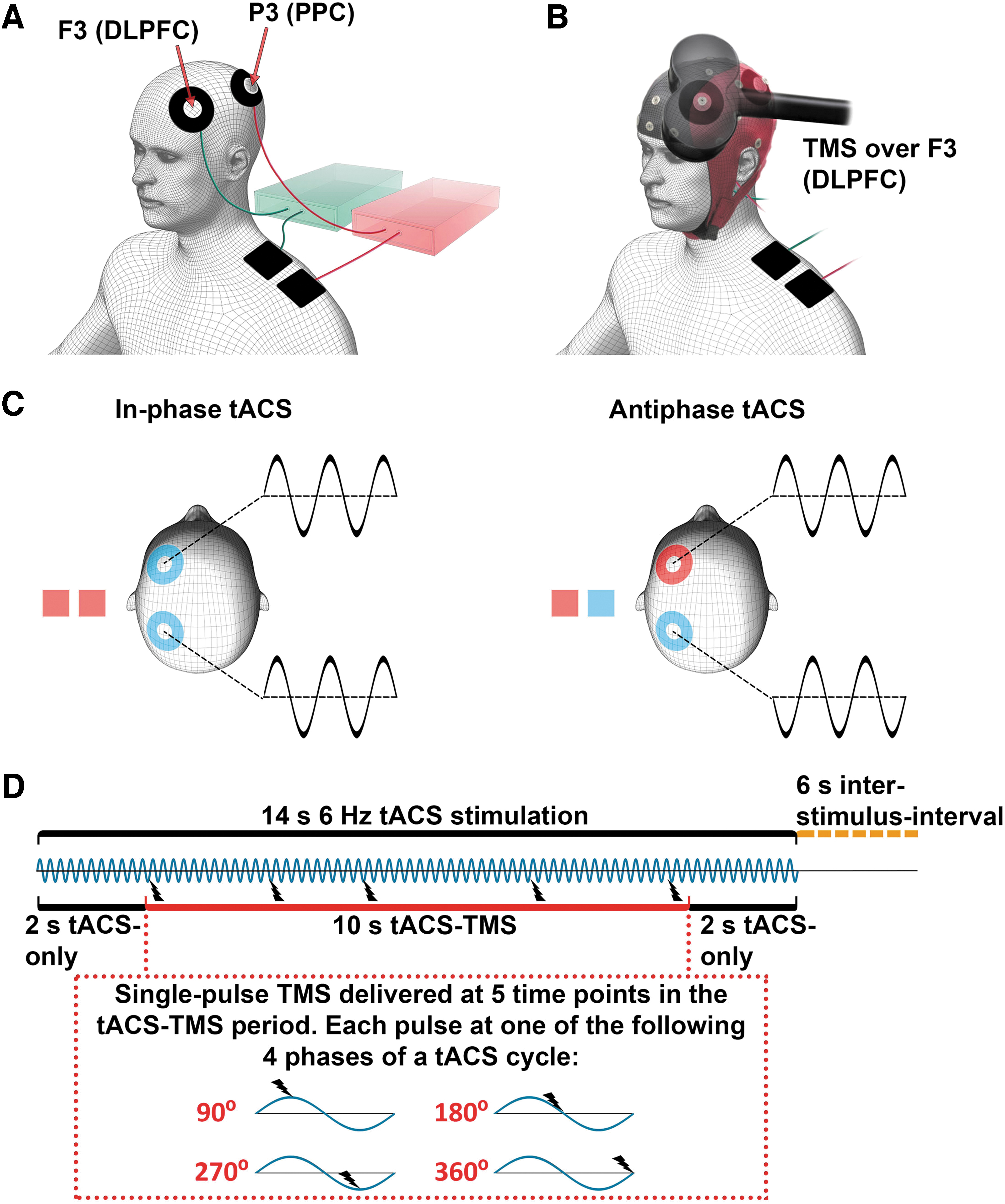
Experimental setup for concurrent tACS-TMS-EEG recording.
All subjects were comfortably seated in a dimly lit room. They were instructed to relax, fixate on a point in front of them, and keep still during the recording and the stimulation. Subjects were also instructed to wear earplugs during measurements, to reduce the auditory click noise produced by the TMS coil. A chin-rest was used to restrict head movements, and the position of the TMS coil over the scalp was fixed with a tripod to ensure constant coil placement throughout the experiment.
tACS was applied by two battery-driven stimulators (DC-Stimulator plus; NeuroConn GmbH). Each stimulator was connected to one stimulating electrode placed on the scalp and one extra cephalic return electrode on the ipsilateral shoulder, made of conductive rubber (NeuroConn GmbH). The two doughnut-shaped scalp electrodes (outer diameter, 60 mm; inner diameter, 25 mm; surface area, 23.4 cm2) were placed on the left hemisphere centered at channels F3 (DLPFC) and P3 (PPC) (Fig. 1A), respectively. The return electrodes (50 × 50 mm; surface area, 25 cm2) were placed on the ipsilateral shoulder of the participant. The tACS electrode montage was selected by means of electric field modeling (Supplementary Fig. S1). The tACS electrodes were attached to the scalp and to the shoulder with EEG gel, as described in a previous protocol article (Fehér and Morishima, 2016). The combined skin resistance for each tACS electrode pair was kept below 10 kΩ.
We recorded eight sessions of concurrent tACS-TMS-EEG, during which tACS with a sinusoidal waveform was applied with a peak-to-peak intensity of 0.9 mA at a frequency of 6 Hz. Each of the tACS-TMS-EEG sessions lasted for ∼6 min, between which the subjects were allowed to take a short break. Within each of the eight recording sessions, tACS was delivered in blocks of 14 sec. Each recording session contained 16 tACS-blocks, interleaved by 6-sec inter-stimulus intervals (Fig. 1D). tACS was delivered to the DLPFC and the PPC in an in-phase manner for half of the stimulation blocks and in an antiphase manner (i.e., 180° shifted) for the other half of the stimulation blocks (Fig. 1C). The order of in-phase and antiphase tACS-blocks was randomized.
Within each tACS-block, single pulses of TMS (at 40% of maximal output intensity) were applied over the DLPFC at five time-points. The inter-TMS-interval within a tACS-block was jittered between 2 and 4 sec (i.e., 12–24 tACS-cycles). The first and last pulse of each tACS-block was delivered 2 sec after the onset and before the offset of tACS, respectively. Each TMS-pulse was applied at one of four different phases of a tACS-cycle (90°, 180°, 270°, 360°; Fig. 1D). Thus, each tACS-block contained TMS pulses delivered at four different phases, with one delivery phase occurring twice (counterbalanced across blocks). The order of TMS-delivery phases within a tACS block and within a recording session was randomized. A total of 80 TMS-pulses was delivered during each recording session, and the occurrence of TMS-delivery phase within a session was balanced. Across all 8 sessions, a total 640 pulses was delivered, with 80 TMS trials per condition (in-phase tACS 90°, 180°, 270°, 360°, antiphase tACS 90°, 180°, 270°, and 360°). An analogue output board (model NI PCI-6723; National Instruments) was used to control the tACS stimulators and the TMS device and to send triggers to the EEG system. The output board was controlled through the data acquisition toolbox for MATLAB (MathWorks, Inc., Natick, MA).
EEG data preprocessing and analysis
EEG data preprocessing was performed by using MATLAB and the EEGLAB toolbox available online (Delorme and Makeig, 2004). The tACS-related and TMS-related artifact were removed as detailed in the Supplementary Methods section in Supplementary Data, and as further elaborated in our previous publication (Fehér et al., 2017). After their removal, traces were visually inspected and trials affected by prominent artifacts were rejected. On average across subjects, 94.56% of the trials (i.e., 75.65 trials per TMS-delivery condition) were retained for further processing (Supplementary Table S1). The artifact-cleaned data were subsequently re-referenced to a common average of all electrodes, excluding channels Fp1, Fpz, Fp2, F3, and the mastoids, which had been previously removed. The mastoids were excluded, as tACS-related artifacts on EEG data were not effectively removed due to muscle artifacts across subjects. At channel F3, we observed a longer decaying time of the TMS artifact, preventing the complete removal of tACS-related artifacts. Channel F3 as well as the frontal channels Fp1, Fpz, and Fp2 were furthermore highly contaminated with random noise unrelated to the application of TMS. The channels were, therefore, excluded from further analyses. Baseline correction was performed by subtracting the average of the −70 to −10 msec period with respect to the TMS onset. We also excluded the first 20 msec after TMS from the analysis to avoid TMS artifacts (Supplementary Figs. S2 and S3).
Statistical analyses were performed on TEPs in MATLAB. Phase-dependent TEPs differences were analyzed separately for the in-phase and antiphase tACS data through cluster-based permutation tests for one-way repeated-measures analysis of variance (ANOVA), calculated for mean TEPs across time windows of 20 msec after TMS. Due to the noisy nature of the data, we reasoned that an identification of clusters across channels and time-bins would remove isolated, noise-related findings. In addition, channel-wise phase-dependent TEPs differences were analyzed, separately for the in-phase and antiphase tACS data, through one-way repeated-measures ANOVAs, for mean TEPs across time windows of 20 msec after TMS. We also addressed global modulation of TEPs across channels, by calculating the global mean field power (GMFP) over all channels (Lehmann and Skrandies, 1980). Phase-dependent differences in GMFP were analyzed separately for the in-phase and antiphase tACS data through one-way repeated-measures ANOVAs, calculated across time windows of 10 msec after TMS in the time-period 20–250 msec after TMS. The interaction between the TMS-delivery phase and the phase relation of tACS (in-phase and antiphase) on the GMFP was analyzed through two-way repeated-measures ANOVAs, calculated for GMFP across time windows of 10 msec after TMS in the time-period of 20–250 msec after TMS.
Finally, we performed voxel-wise statistics on individual source localized data (standardized low-resolution brain electromagnetic tomography [sLORETA]) (Pascual-Marqui, 2002) from mean TEPs per condition. The interaction effects between phase relation of tACS (in-phase and antiphase) and TMS delivery phase on TEPs were analyzed by two-way repeated-measures ANOVAs, calculated voxel-wise within the time windows of 110–130 msec after TMS. We also performed planned pair-wise comparisons of TEPs of the 90° and 180° phases and the 270° and 360° phases of tACS, calculated for the mean region of interest voxels in the time-window 120–130 msec after TMS. A significance threshold of 0.05 was used for all of our statistical analyses.
Results
Phase dependence of TEPs
First, we addressed the condition-specific global propagation of TMS-induced activity, by calculating the GMFP for TEPs per TMS-delivery phase obtained during in-phase and antiphase tACS. The condition-specific GMFPs were compared in the time-period 20–250 msec after TMS (Fig. 2A). The one-way repeated-measures ANOVA yielded significant differences in GMFPs between TMS-delivery phases during in-phase tACS, in the time windows of 90–220 msec after TMS (p < 0.05) (Fig. 2A). No further phase-dependent changes in GMFPs manifested beyond 210 msec after TMS. During antiphase tACS, we found significant differences in GMFPs between TMS-delivery phases in the time windows 150–230 msec after TMS (p < 0.05) (Fig. 2B).
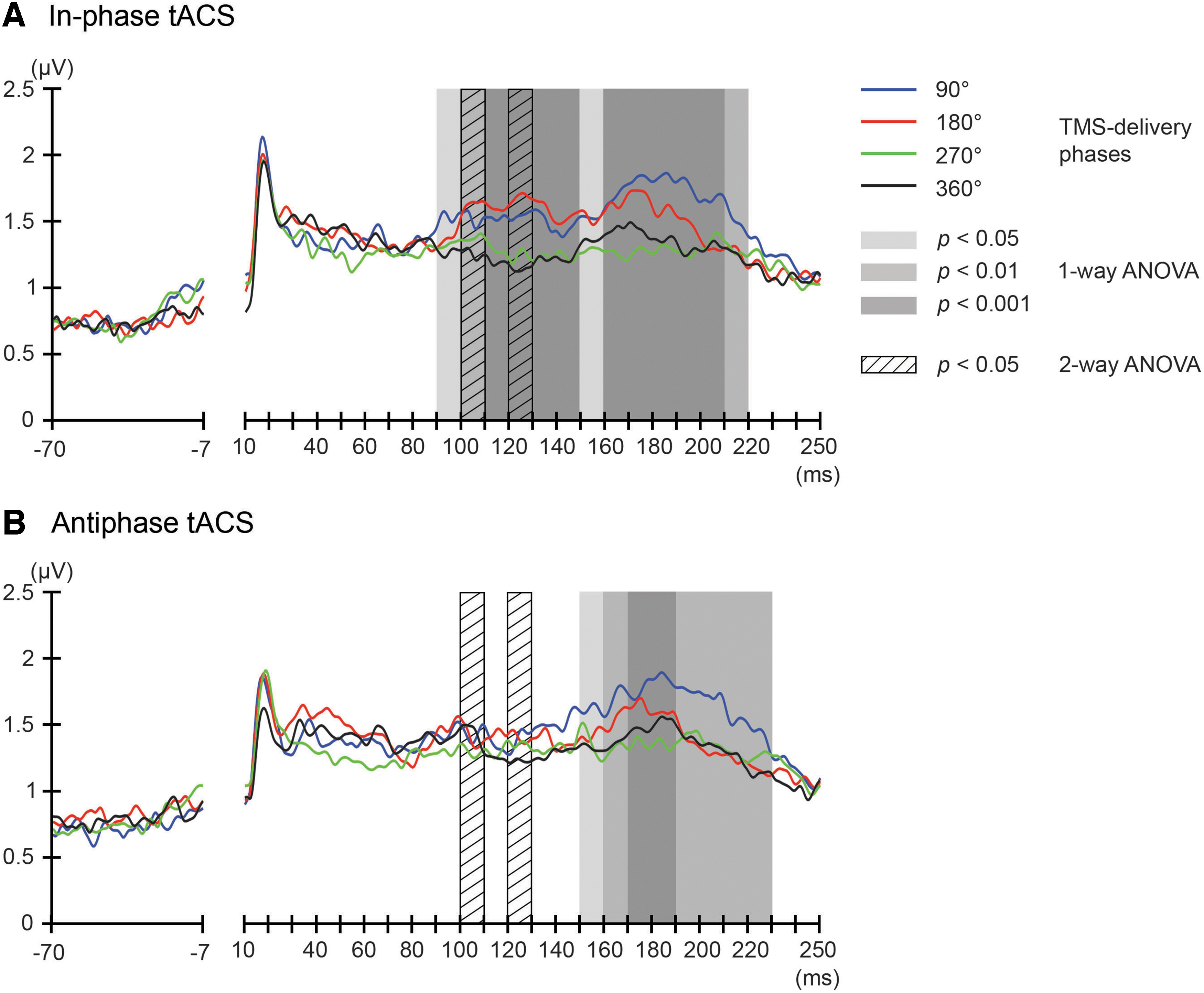
Phase-dependent modulation of excitability during in-phase and antiphase tACS as measured by GMFP. The onset of TMS corresponds to 0 msec of the plot. The shaded areas mark time windows with significant effect (p < 0.05 to p < 0.001) of TMS-delivery phase on GMFPs during in-phase
The analysis of GMPFs suggests that a tACS-induced oscillatory phase relation (in-phase or antiphase) between the DLPFC and PPC influences the degree of phase-dependent GMPFs modulations. To formally test this, we first performed a two-way repeated-measures ANOVA for the within-subject factors TMS-delivery phase and tACS condition on our GMFP data in time windows of 10 msec. We observed a significant interaction effect of tACS condition and TMS-delivery phase in the time-period of 100–110 and 120–130 msec after TMS (p < 0.05) (Fig. 2).
We proceeded to address the local modulation of excitability. A channel-wise one-way repeated-measures ANOVA yielded significant differences between TMS-delivery phases for the in-phase tACS dataset from the time-window 20–40 msec after TMS, at parietal channels P3 [F(3,18) = 3.65; p = 0.018] and CP1 [F(3,18) = 4.138; p = 0.01], and at contralateral channels FC6 [F(3,18) = 2.8; p = 0.049] and CP6 [F(3,18) = 3.1; p = 0.034] (Fig. 3, fifth row). From the time-window 100–120 msec after TMS, significant phase-dependent differences in TEPs (p < 0.05) emerged in posterior as well as contralateral regions within the same time-window. For the antiphase tACS dataset, the channel-wise one-way repeated-measures ANOVA yielded significant differences in TEPs between TMS-delivery phases at central channel Cpz, beginning from the time window of 20–40 msec after TMS [F(3,18) = 6.231; p = 0.001], and at contralateral channel T8 [F(3,18) = 3.069; p = 0.03] (Fig. 4, fifth row). In later time windows, significant phase-dependent differences in TEPs (p < 0.05) emerged in contralateral regions 100 msec after TMS and in posterior regions 160 msec after TMS.
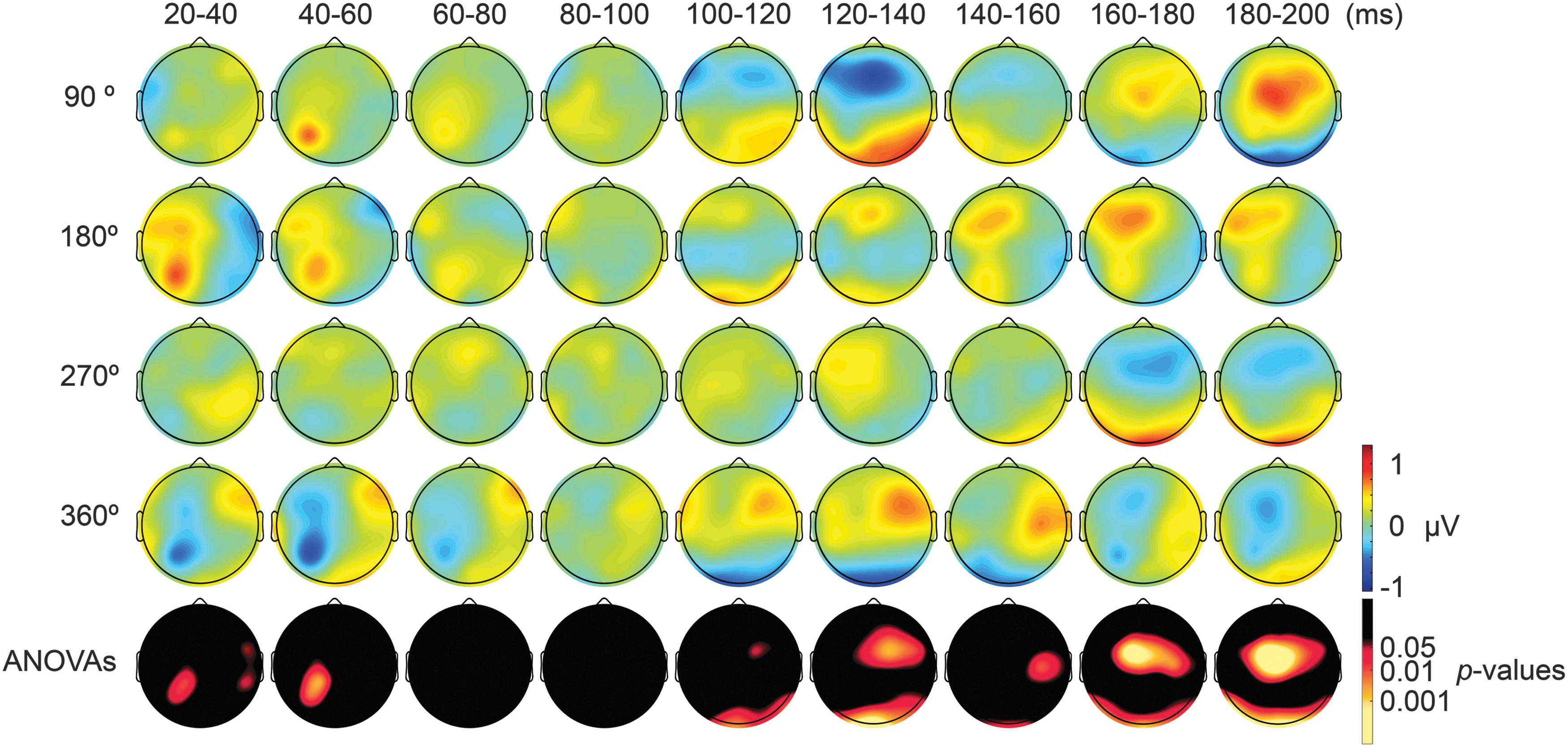
Scalp maps of TEPs during in-phase tACS showing mean across participants, and statistical analyses. Top four rows: scalp topographies of TEPs between 20 and 200 msec after TMS during 6-Hz tACS applied to the DLPFC and to the PPC in an in-phase manner. The average TEP across the four TMS-delivery phase conditions is subtracted from each TMS-delivery phase condition. The TMS was applied at 90°, 180°, 270°, or 360° of the tACS phase. Channels Fp1, Fpz, Fp2, F3, as well as the mastoids were excluded from the plot, and the potentials were interpolated from the surrounding channels. Fifth row: scalp topographies of p-values across time based on channel-wise one-way repeated-measures ANOVA for the effect of the delivery phase of the TMS. The p-value of each electrode is shown in color only when it reaches significance (p < 0.05). TEPs, TMS-evoked potentials.

Scalp maps of TEPs during antiphase tACS showing mean across participants, and statistical analyses. Top four rows: scalp topographies of TEPs between 20 and 200 msec after TMS during 6-Hz tACS applied to the DLPFC and to the PFC in an antiphase manner. The average TEP across the four TMS-delivery phase conditions is subtracted from each TMS-delivery phase condition. TMS was applied at 90°, 180°, 270°, or 360° of the tACS phase. Channels Fp1, Fpz, Fp2, F3, as well as the mastoids were excluded from the plot, and the potentials were interpolated from the surrounding channels. Fifth row: scalp topographies of p-values across time based on channel-wise one-way repeated-measures ANOVA for the effect of the delivery phase of the TMS. The p-value of each electrode is shown in color only when it reaches significance (p < 0.05).
Together, these results confirm that 6-Hz tACS can modulate cortical excitability in a phase-dependent manner, consistent with our previous study (Fehér et al., 2017). In addition, phase-dependent difference of TEPs propagation is more efficiently preserved during in-phase tACS.
Spread of phase-dependent TEP modulation to posterior brain regions
From our channel-wise one-way repeated-measures ANOVA, we observed significant phase-dependent differences in TEPs between TMS delivery phases during in-phase tACS in this channel cluster within the time-window 100–200 msec (p < 0.05) (Fig. 3, fifth row). However, the channel-wise comparisons of TEPs suffer from two problems. First, it suffers from a multiple comparisons issue. This is an issue in particular as, due to the noisy nature of our data, it may lead to the identification of isolated, noise-related differences. These issues may be circumvented by cluster-based non-parametric permutation tests. However, it should be noted that while providing a more robust test, caution should be taken when interpreting the precise “when” and “where” of the identified significant channel and time-bin clusters (Sassenhagen and Draschkow, 2019). We, therefore, tested the condition-specific effect of the delivery phase of the TMS based on cluster-based permutation tests [as implemented in the FieldTrip analysis toolbox (Oostenveld et al., 2011)]. For the in-phase tACS dataset, the cluster-based permutation tests yielded two significant clusters from the time-window 120–130 msec after TMS, with the first one at posterior channels (p < 0.05; Supplementary Fig. S4). For the antiphase tACS dataset, we found three significant clusters from the time-window 180–200 msec after TMS, with the first one at central channels (p < 0.05; Supplementary Fig. S4). No clusters extended to posterior channels. These results confirm the main conclusions from the time- and channel-wise tests.
Phase-relations shape phase-dependent TEPs in parieto-occipital areas
The results cited earlier indicate that the phase dependence of TEPs was maintained beyond the frontoparietal network, and that the phase-dependent TEPs transmitted from the PPC were affected by the manipulation of oscillatory phase relation between the DLPFC and the PPC. Therefore, to assess the area at which the tACS-induced oscillatory phase relation affected the spread of signals, we performed source localization on the data. We converted condition-specific TEPs to sLORETA data and performed voxel-wise two-way repeated-measures ANOVAs for the time windows 100–110 and 120–130 msec after TMS, which were chosen based on the observed interaction effect between the tACS condition and TMS-delivery phase on the GMFP. In accordance with our prediction, we observed a significant interaction effect of phase relation of tACS and TMS-delivery phase in the time-window 120–130 msec after TMS in a bilateral posterior cluster [peak MNI coordinate at the right inferior parietal lobule: x = 45, y = −55, z = 40, F(3,18) = 0.079, p ≤ 0.0001; Fig. 5].
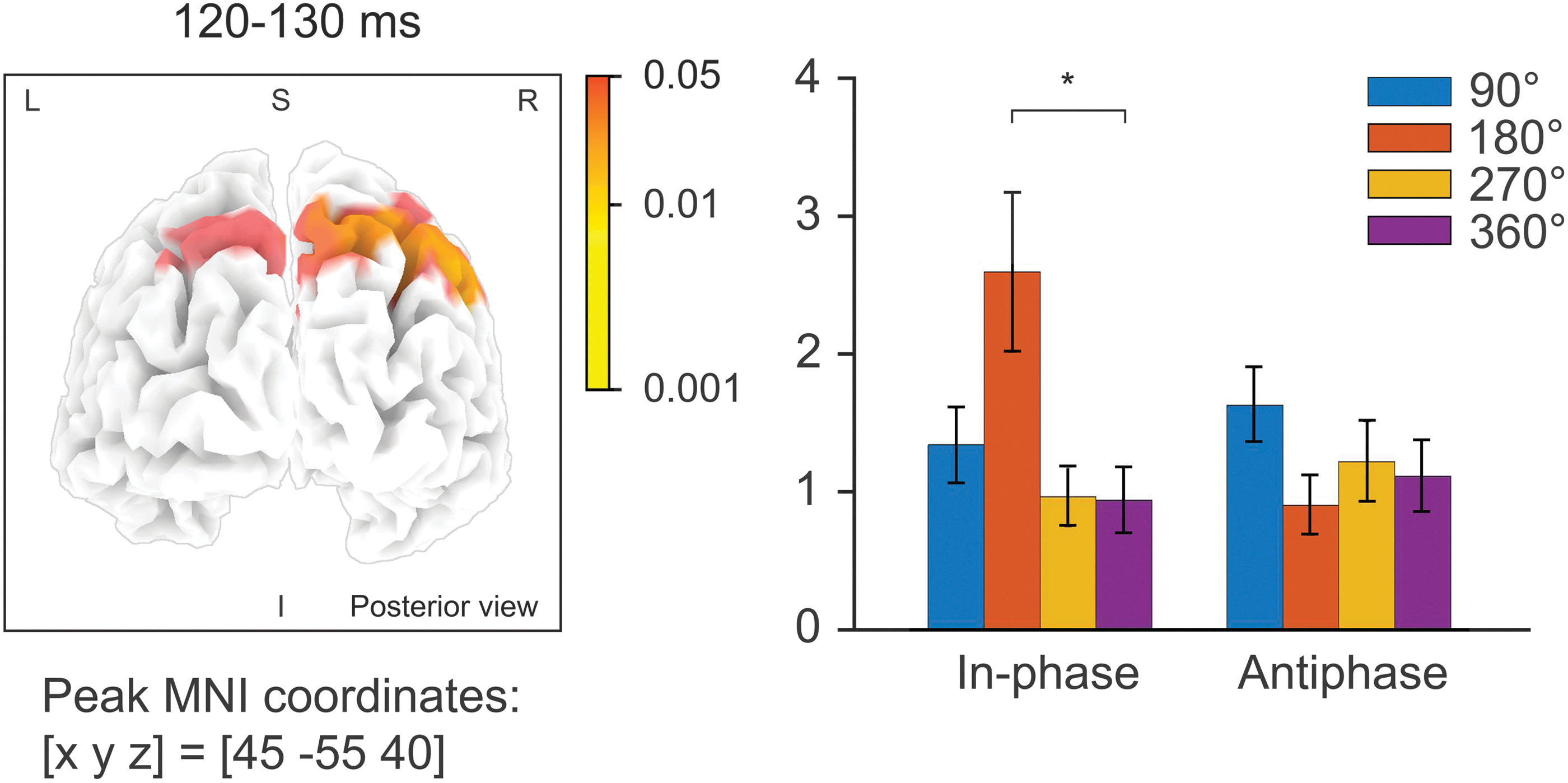
p-Values calculated on sLORETA source estimates across participants from TEPs during in-phase and antiphase tACS, 120–130 msec after TMS. The left panel shows the interaction effect of tACS phase relation (in-phase and antiphase) and TMS-delivery phase, calculated voxel-wise. The p-value of each electrode is shown in color only when it reaches significance (p < 0.05). The peak coordinate of the posterior cluster in the time-window 120–130 msec after TMS was located at the right inferior parietal lobule [peak MNI coordinate: x = 45, y = −55, z = 40, F(3,18) = 0.079, p ≤ 0.0001]. For visualization purposes, the right panel shows source activation at the peak coordinate of the plotted for each tACS phase relation and each TMS-delivery phase. Error bars signify standard error. *Signifies a p-value <0.05. Pairwise comparisons were corrected for multiple comparisons (Bonferroni correction). sLORETA, standardized low-resolution brain electromagnetic tomography.
As exploratory post hoc analyses of TMS-induced source activity, we compared condition-specific sLORETA activity with respect to TMS phase relations during in-phase and antiphase tACS. Condition-specific one-way ANOVAs were significant at the peak voxel only for the in-phase condition [120–130 msec: F(3,18) = 6.44, p ≤ 0.0008; 130–140 msec: F(3,18) = 4.305, p ≤ 0.009], but not for the antiphase. Post hoc pairwise comparisons showed a significant difference between the 180° and 360° TMS-delivery phase in the time-window 120–130 msec after TMS (p = 0.001, Bonferroni corrected), at the peak MNI coordinate (Fig. 5). The results suggest that in-phase tACS enhances neural transmission from the DLPFC in the time window 120–130 msec after TMS.
Discussion
We recently introduced the concurrent tACS-TMS-EEG method to measure the immediate effects of tACS on local excitability and neuronal communication efficacy across cortical networks (Fehér et al., 2017). In the present study, we employed this method to address the question whether tACS-induced synchronization between the DLPFC and the PPC nodes of the frontoparietal network can modulate signaling efficacy within the frontoparietal network, as well as beyond the frontoparietal network. The main finding of the current study is that modulation of the oscillatory phase relation between the DLPFC and the PPC not only affects the maintenance of phase dependence of the induced signal within the frontoparietal network, but it also modulates phase-dependent transmission beyond the synchronized networks.
Models of the role of neuronal oscillations in the brain can be classified into two larger categories, focusing on the routing of neuronal signaling and on the informational content of neuronal signaling, respectively. The influential communication-through-coherence (CTC) model belongs to the former category. Our results lend support to two of its major principles. First, replicating the findings of our previous study (Fehér et al., 2017), we once again demonstrate that tACS-induced theta oscillations modulate cortical excitability in a phase-dependent manner. This supports the phase-dependent modulation of neuronal signaling efficacy by regional endogenous oscillatory activity. The CTC then proposes that the added periodicity to the efficacy of signal transmission can be capitalized upon by modulating the inter-regional oscillatory phase relation. By modulating the inter-regional phase relation, the state of excitability in a downstream region at which a signal will arrive can be systematically controlled. We show that the propagation of the maintained phase-dependent TEPs to the posterior regions depends on the tACS-induced oscillatory phase relation between the DLPFC and the PPC. This lends support to the role of inter-regional phase relation of endogenous oscillatory activity in modulating inter-regional signal transmission. The second category of models on the role of neuronal oscillations suggests that oscillatory activity adds another dimension to neuronal information coding, proposing that information is coded in the timing of the neuronal spikes relative to the phase of neuronal oscillations (Buzsáki and Chrobak, 1995; O'Keefe and Recce, 1993). The maintenance of phase dependence is a prerequisite for the communication of phase-coded information between network nodes. Although we do not directly address further predictions specific to phase-coding models of oscillatory activity, our results demonstrate how the tACS-induced inter-nodal oscillatory phase relation affects the maintenance of phase dependence across and beyond the modulated network nodes.
The present study replicates the core findings of phase-dependent modulation of TEPs observed in our previous study (Fehér et al., 2017). We observe that the TEPs depended on the phase of the introduced 6 Hz activity during in-phase as well as during antiphase tACS. The timeline for emergence of phase-dependent TEP modulation in posterior areas in our in-phase tACS condition (Fig. 3) is also in agreement with the timeline that we have previously observed in our DLPFC-only tACS condition (Fehér et al., 2017). When comparing DLPFC-only with in-phase (i.e., simultaneous DLPFC and PPC) tACS, the essential difference is that we left the state of excitability at the PPC uninfluenced when tACS was solely applied to the DLPFC. If the additional manipulation of oscillatory activity in the PPC in the present study had an impact on the phase-dependent transmission to the occipital region, this would manifest in the magnitude of phase-dependent TEP modulation, but not in the timing of the spread of phase-dependent TEPs. In the DLPFC-only tACS condition of our previous study, significant phase-dependent differences in TEPs had emerged over a posterior cluster of channels at 100–140 and 160–200 msec after TMS (Fehér et al., 2017). Consistent with this, we observed significant phase-dependent differences in TEPs between TMS delivery phases during in-phase tACS in this channel cluster within the time-window 100–200 msec.
Here, we provided evidence that the difference in transmission of phase-dependent TEPs is reflected in the inputs from the PPC to connected areas. Although the contralateral PPC is well connected to the PPC via transcallosal connections (Barbas and Pandya, 1984; Hofer et al., 2008), we also expected to observe differences in the spread of tACS-phase-dependent TEP modulation not only within the left PPC, but also in the left extrastriate areas that have been shown to be strongly connected to and modulated by top–down influences from the PPC (Lewis and Van Essen, 2000; Mars et al., 2011; Maunsell and van Essen, 1983; Ruff et al., 2006; Tong, 2003; Ungerleider and Desimone, 1986). Although we did observe weaker effects in the left PPC in source space, the lack of findings in source space in the proximity of the left PPC such as the ipsilateral extrastriatal areas may be due to the tACS-induced noise in this area masking such effects.
Sustained elevated power in the targeted frequency band is assumed to reflect the efficacy of tACS in driving ongoing intrinsic oscillatory activity. Several studies have evidenced that these after-effects are influenced by the momentary brain state during stimulation, dictated, for instance, by the task performed by the subject while being stimulated (Feurra et al., 2013; Kar and Krekelberg, 2014), or whether the subjects had their eyes open or closed (Alagapan et al., 2016; Neuling et al., 2013). Recently, two studies have also addressed the online state-dependent effect of tACS by using concurrent tACS and magnetoencephalography, showing how keeping the eyes open or closed affected the efficacy of the entrainment during stimulation (Fuscà et al., 2018; Ruhnau et al., 2016). Although we did not address momentary intrinsic brain states in the present study, the concurrent tACS-TMS-EEG method could be applied in future studies to further explore how intrinsic brain states influence the effect of tACS on excitability and signaling efficacy during stimulation.
Due to functional limitations of the stimulators that we used, application of tACS over two scalp locations added additional noise to our EEG data, as compared with using a single scalp tACS electrode location as done in our previous study (Fehér et al., 2017). To further increase the efficacy of our artifact removal procedure, we removed the highly contaminated frontal channels from our data. Due to the thorough cleaning measures, a substantial fraction of neural signals might have been removed in the early time-periods after TMS, with this being the potential reason why we see little results within the first 100 msec after TMS as compared with the DLPFC-only tACS condition of our previous study.
The tACS artifact removal process leads to a reduction in power up to about 15 Hz (Supplementary Fig. S5), but is otherwise able to recover the TEPs, as can be seen when comparing data from TMS without and with concurrent tACS, after artifact removal. As an online sham, we also analyzed the data in the period 1000–1500 msec after the TMS, and we found no phase-dependent differences between TEPs. Lastly, we have in a previous publication shown that phase-dependent differences between TEPs arose with a delay when tACS was applied not to the region under the TMS but to the parietal cortex, as compared with applying tACS directly to the DLPFC under the TMS (Fehér et al., 2017). Together, these results support that the artifact removal pipeline could recover the TEPs during tACS and is unlikely to create false-positive differences among TMS-delivery conditions, verifying the validity of our results.
Some concern of using dual-site tACS to address the role of inter-nodal phase relations in network communication and cognitive operations has recently received increased attention. One concern in particular is that the density of current in the area of interest differs between the tACS conditions, and this might explain the results observed during such studies (Saturnino et al., 2017). Our modeling of current density distribution does not seem to indicate a lower current density at our targeted DLPFC and PPC nodes of interest during antiphase tACS as compared with during in-phase tACS (Supplementary Fig. S1). A prediction from a lesser influence of tACS on excitability in the targeted region in one of the tACS conditions would have been the observation of attenuated phase-dependent differences in TEPs in this condition. However, our data did not indicate the latter nor did it point to an alteration in the spread of phase-dependent TEPs between the in-phase and antiphase conditions, beyond the delayed propagation to posterior regions (Figs. 3 and 4), supporting that a comparable influence over excitability was exhibited during antiphase and in-phase tACS. For the purpose of improving the reliability of comparison between the effect of in-phase and antiphase tACS however, especially for reliably addressing behavioral effects, Saturnino et al. (2017) recently suggested an electrode montage for implementing in-phase and antiphase tACS between the DLPFC and the PPC that may avoid differences in current density distribution and current flow between in-phase and antiphase tACS. It would be of interest to replicate the present study with these montage recommendations, and the recommended montage would be of particular use for replicating previously observed behavioral effects of tACS-modulated inter-nodal phase relations.
The concurrent tACS-TMS-EEG method holds great promise in adding yet another measure of underlying neurophysiological mediators of behavioral effects of tACS-modulated oscillatory activity; in this case the effects on excitability and signal transmission during tACS. Although measuring motor-evoked potentials is limited to address the neurophysiological response to tACS within the primary motor cortex (Guerra et al., 2016; Nakazono et al., 2016; Raco et al., 2016), the concurrent tACS-TMS-EEG method can be applied to address regionally and network-specific effects of tACS on excitability and neuronal signal transmission across cortico-cortical networks. In future applications, the method could be employed to address how changes in excitability and signal transmission during tACS mediate the behavioral outcome measure during and after tACS. tACS as well as transcranial electric stimulation in general has been shown to suffer from both large inter- and intra-individual variation in behavioral effects (Horvath et al., 2014, 2015a,b; Hsu et al., 2016; Li et al., 2015; Veniero et al., 2017). More online measures of the neurophysiological effects of tACS are needed to unravel factors that contribute to those variations in behavioral performance modulation, and to correlate the specific individual neurophysiological response to the behavioral outcome measure of interest.
Conclusions
Taken together, we have shown that the tACS-induced oscillatory phase relation across the frontoparietal network affects the propagation of maintained phase-dependent TMS-induced activity across and, importantly, beyond the frontoparietal network. Our results add new insights into the widespread effect of tACS beyond the stimulated area and add evidence for the causal role of inter-nodal endogenous oscillatory dynamics in modulating inter-nodal information flow.
Footnotes
Acknowledgment
The authors thank T. Koenig for advice on the EEG setup.
Authors' Contributions
Y.M conceptualized the experiments. K.D.F., M.N., and Y.M. collected data, and K.D.F. and Y.M. analyzed data. K.D.F. and Y.M. discussed and interpreted the results and wrote the article.
Author Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This work was supported by the Japan Science and Technology Agency PRESTO program (10238; Y.M.).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.