
Abstract
Introduction:
Comorbid mild traumatic brain injury (mTBI) and post-traumatic stress disorder (PTSD) are common in military service members. The aim of this study is to investigate brain metabolic interrelationships in service members with and without persistent PTSD symptoms after mTBI by using 18F-fluorodeoxyglucose (FDG) positron emission tomography.
Methods:
Service members (n = 408) diagnosed with mTBI were studied retrospectively. Principal component analysis was applied to identify latent metabolic systems, and the associations between metabolic latent systems and self-report measures of post-concussive and PTSD symptoms were evaluated. Participants were divided into two groups based on DSM-IV-TR (Diagnostic and Statistical Manual of Mental Disorders, fourth edition–Text Revision) criteria for PTSD, and structural equation modeling was performed to test a priori hypotheses on metabolic interrelationships among the brain regions in the cortico-limbic circuitry responsible for top–down control and bottom–up emotional processing. The differences in metabolic interrelationships between age-matched PTSD-absent (n = 204) and PTSD-present (n = 204) groups were evaluated.
Results:
FDG uptake in the temporo-limbic system was positively correlated with post-concussive and hyperarousal symptoms. For the bottom–up emotional processing, the insula and amygdala-hippocampal complex in the PTSD-present group had stronger metabolic interrelationships with the bilateral rostral anterior cingulate, left lingual, right lateral occipital, and left superior temporal cortices, but a weaker relationship with the right precuneus cortex, compared with the PTSD-absent group. For the top–down control, the PTSD-present group had decreased metabolic engagements of the dorsolateral prefrontal cortex on the amygdala.
Discussion:
Our results suggest altered metabolic interrelationships in the cortico-limbic circuitry in mTBI subjects with persistent PTSD symptoms, which may underlie the pathophysiological mechanisms of comorbid mTBI and PTSD.
Impact statement
This is the first 18F-fluorodeoxyglucose positron emission tomography study to investigate brain metabolic interrelationships in service members with persistent post-traumatic stress disorder (PTSD) symptoms after mild traumatic brain injury (mTBI). We identified that the temporo-limbic metabolic system was associated with post-concussive and hyperarousal symptoms. Further, brain metabolic interrelationships in the cortico-limbic circuitry were altered in mTBI subjects with significant PTSD symptoms compared with those without them.
Introduction
Traumatic brain injury (TBI), the majority of which are mild (Lindquist et al., 2017), and post-traumatic stress disorder (PTSD) are often considered to be the signature injuries of the latest military conflicts (Hines et al., 2014; Howlett and Stein, 2016). However, their pathophysiological bases remain unclear (Howlett and Stein, 2016).
Accumulating evidence has identified TBI as an important risk factor for PTSD (Bryant, 2011). In a large study of active-duty Marine service members who served in Iraq or Afghanistan and reported minimal to no symptoms before deployment, deployment-related TBIs nearly doubled the odds of developing PTSD (Yurgil et al., 2014). The prevalence of comorbid TBI and PTSD in military personnel is high. In a study of service members after their return from a year-long deployment to Iraq, 43.9% of those with mild TBI (mTBI) who had loss of consciousness met the criteria for PTSD (Hoge et al., 2008). Further, comorbid mTBI and PTSD increase the risk of neuropsychological deficits and psychiatric disorders, relative to mTBI alone (Combs et al., 2015; Lange et al., 2021; Pietrzak et al., 2009). Understanding how PTSD interacts with mTBI to negatively impact neurocognitive functioning and neurobehavioral abilities is important for prognosis and treatment recommendations.
The PTSD symptoms include intrusive thoughts, avoidance, hyperarousal and hypervigilance, flashbacks, emotional numbing, sleep disturbances, and changes in memory and concentration (Hayes et al., 2012b). The PTSD symptoms can be caused by a failure of cortical regions to dampen the exaggerated symptoms of arousal and distress mediated through the amygdala in response to reminders of the traumatic event (Nutt and Malizia, 2004). Meta-analyses of PTSD have identified key regions in the cortico-limbic circuit, including the amygdala, hippocampus, anterior cingulate cortex (ACC), and prefrontal cortex (PFC) as having consistent changes in neural activity (Etkin and Wager, 2007; Hayes et al., 2012a). The amygdala is a key limbic structure involved in the modulation of affectively influenced memory and emotional reactivity in the cortico-limbic circuit, over which regions of the PFC exert an inhibitory effect (Lobo et al., 2011; Phelps et al., 2004). On the other hand, the hippocampus is involved in the modulation of memory functions (Brohawn et al., 2010) and fear-related learning processes (Corcoran et al., 2005; Quirk and Mueller, 2008). The amygdala-hippocampal complex governs two independent memory systems that act together when complex emotional memories are created or retrieved (Phelps, 2004). The ACC plays an important role in both bottom–up emotional processing and top–down inhibitory control by acting as an amplifier and attenuator of emotional and self-referential information.
The bottom–up emotional processing is driven by salient stimuli. The amygdala has been implicated in emotional salience and encoding emotional relevance, and it plays an important role in the consolidation of information from other brain regions such as the insula and hippocampus during an emotional experience (Fitzgerald et al., 2018). The insula is involved in interoceptive awareness of an emotional state (Craig, 2009; Grupe and Nitschke, 2013), and the hippocampus is involved in contextual learning and emotional memory (Opitz, 2014; Strange and Dolan, 2006). Since interactions among those regions during emotional processing were not clearly known, a latent variable was constructed in this study as an underlying cause of their metabolic activities. We hypothesized that the latent variable has an influence on other diverse brain regions involved in PTSD, including the rostral ACC (rACC) (Musgrove et al., 2015; Weston, 2014), precuneus (Rangaprakash et al., 2019), visual cortices (lingual and lateral occipital cortices) (Chao et al., 2012; Weston, 2014), ventromedial PFC (vmPFC), and superior temporal cortex (Fenster et al., 2018; Weston, 2014).
For the top–down control, the dorsolateral PFC (dlPFC) has long been thought to be critically involved in cognitive control (Aupperle et al., 2012; Brosnan and Wiegand, 2017; Egner and Hirsh, 2005), whereas the vmPFC (Hiser and Koenigs, 2018; Koenigs and Grafman, 2009; Urry et al., 2006) and rACC (Newsome et al., 2013; Szekely et al., 2017) are integral components of the default mode network (DMN) that regulate emotional responses by controlling the functions of the amygdala. The amygdala has greater white matter connectivity with the medial PFC (mPFC) and rACC relative to the dlPFC and dorsal ACC (dACC) (Goetschius et al., 2019). The influence of the dlPFC on the amygdala is likely mediated by the rACC and vmPFC that have strong connections with the amygdala (Beckmann et al., 2009; Ongür and Price, 2000).
Studies have examined comorbid mTBI and PTSD by using 18F-fluorodeoxyglucose (FDG) positron emission tomography (PET), and they found larger cluster size of low uptake voxels (Buchsbaum et al., 2015) and hypometabolic activities in the brain regions such as the cerebellum, vermis, pons, and medial temporal lobe (Peskind et al., 2011), compared with control groups. However, no prior FDG PET study has investigated brain metabolic interrelationships in comorbid mTBI and PTSD, although functional magnetic resonance imaging (fMRI) studies have reported abnormal resting-state functional connectivity within the DMN in subjects with comorbid mTBI and PTSD (Nathan et al., 2017; Santhanam et al., 2019b).
A multi-modal study with healthy participants showed that DMN functional connectivity during rest is closely related to metabolic activity by comparing fluctuations in FDG uptake and blood oxygen level-dependent signal detected in fMRI (Passow et al., 2015). Therefore, we hypothesize that brain metabolic interrelationships are altered in mTBI subjects with significant PTSD symptoms compared with those without them. In this study, we performed principal component analysis (PCA) to identify latent metabolic systems on brain FDG uptake of service members at rest and investigated their association with post-concussive and PTSD symptoms. In addition, we applied structural equation modeling (SEM) to examine metabolic interrelationships between brain regions responsible for the top–down control and bottom–up emotional processing in the cortico-limbic circuit.
Materials and Methods
Participants and clinical assessment
This study was approved by the Institutional Review Board at the Walter Reed National Military Medical Center (WRNMMC) and is compliant with the Health Insurance Portability and Accountability Act guidelines. Military service members were enrolled after providing informed consent at the National Intrepid Center of Excellence (NICoE, Bethesda, MD). All participants were active duty service members. Diagnosis of mTBI was determined during routine comprehensive clinical assessments at the WRNMMC based on VA/DoD clinical practice guidelines (Management of Concussion-Mild Traumatic Brain Injury Working Group, 2016). Study participants who had experienced loss of consciousness that lasted up to 30 min, post-traumatic amnesia for up to 1 day, or alteration of consciousness for up to 24 h were diagnosed with mTBI. Participants with a history of major neurologic or psychiatric conditions such as psychosis, stroke, multiple sclerosis, or spinal cord injury were excluded. A total of 469 participants with chronic (>6 months post-injury) mTBI were evaluated. On the day of PET scan, participants completed the Neurobehavioral Symptom Inventory (NSI) and the PTSD Check List–Civilian Version (PCL-C). The NSI is a self-report measure of 22 non-specific post-concussion symptoms. The NSI total symptom score was divided into four subscores: (1) somatosensory (headache, nausea, vision problems, sensitivity to light, sensitivity to noise, numbness/tingling, change in taste/smell), (2) affective (fatigue, sleep, anxiety, depression, irritability, frustration), (3) cognitive (concentration/distraction, forgetfulness, difficulty making decisions, slowed thinking), and (4) vestibular (dizziness, imbalance, coordination) (Caplan et al., 2010). The PCL-C is a standardized self-report rating scale comprising 17 items that correspond to the key symptoms of PTSD. Since the PCL–Military Version (PCL-M) is focused solely on military-related psychological trauma, this study used the PCL-C so that traumatic events unrelated to military service would not be missed. The PCL-C was divided into three subdomains corresponding to three main syndromes of the disorder: reexperiencing (items 1–5), avoidance (items 6–12), and hyperarousal (items 13–17). To test whether metabolic interrelationships between brain regions are different between mTBI subjects with and without significant PTSD symptoms, study participants were divided into two groups based on DSM-IV-TR (Diagnostic and Statistical Manual of Mental Disorders, fourth edition–Text Revision) criteria for PTSD (American Psychiatric Association, 1994). Participants were classified into the PTSD-present group (n = 204) based on the endorsement of moderate or higher symptoms for one or more Criterion B (reexperiencing) symptoms, three or more Criterion C (avoidance) symptoms, and two or more Criterion D (hyperarousal) symptoms. These participants were age-matched to participants who did not meet DSM-IV-TR criteria for PTSD and assigned to the PTSD-absent group (n = 204). Age matching was carried out by using the R package MatchIt (Ho et al., 2011). The majority of the subjects were male (97%) and had a history of blast exposure (86%). Demographics and clinical characteristics of the study participants are summarized in Table 1.
Descriptive Statistics for Demographics and Clinical Characteristics of Subjects in Two Mild Traumatic Brain Injury Groups
NSI, Neurobehavioral Symptom Inventory; PCL-C, PTSD Check List–Civilian Version; PTSD, post-traumatic stress disorder.
Image acquisition
All mTBI participants were scanned with a PET/CT (computed tomography) scanner (Biograph 64 mCT; Siemens Healthcare, Knoxville, TN). A static PET/CT scan was performed at 50 min after intravenous injection of FDG (9–11 mCi). Participants fasted for 6–8 h before the scan. The 20-min PET images were reconstructed by using the ordered subset expectation maximization algorithm with 6 iterations and 24 subsets (matrix = 400 × 400 × 148, voxel size = 1 × 1 × 1.5 mm3). In addition, participants were scanned with a 3T MRI scanner (Discovery MR 750; GE Healthcare, Milwaukee, WI) by using a 32-channel head coil (MR Instruments, Minneapolis, MN). T1-weighted MR images were acquired with a 3D BRAVO sequence (TR/TE = 6.7/2.5 ms, FA = 12°, matrix = 512 × 512 × 312, voxel size = 0.5 × 0.5 × 0.6 mm3).
Image processing
T1-weighted structural images were parcellated and segmented in the subject's native space by using FreeSurfer 5.3 (Fischl, 2012). The regions of the subcortex and the Deskan-Killiany cortical atlas were segmented, and several PFC regions were combined into larger regions. The vmPFC is composed of two regions defined by FreeSurfer, the medial and lateral orbitofrontal cortices (Boes et al., 2009). The dlPFC was created by combining the superior frontal, rostral middle frontal, and caudal middle frontal cortices, whereas the ventrolateral PFC was created by combining the pars opercularis, pars triangularis, and pars orbitalis (Vijayakumar et al., 2014). A complete list of the regions used in this study can be found in the Supplementary Table S1. Subjects' FDG uptake was normalized to their mean whole brain activity (Buchsbaum et al., 2015; Ito et al., 2016; Mendez et al., 2013; Nakayama et al., 2006; Peskind et al., 2011) and smoothed with a 4 mm FWHM Gaussian kernel. The FDG PET images were registered to T1-weighted images. The mean FDG uptake of 75 brain regions was then calculated.
Principal component analysis
Figure 1 shows the overview of image processing and statistical analyses. Pearson's product-moment partial correlation matrix of FDG uptake was calculated between each pair of the 75 brain regions after controlling for gender and age. The PCA was applied to reduce a large number of the brain regions by finding the eigenvectors (principal components) and the eigenvalues, the amount of variance accounted for by each of these components. The PCA decomposes the correlation matrix and minimizes the sum of the squared perpendicular distances to the component axis. The components that account for at least 85% of the total variance were retained. A varimax rotation was applied to maximize the sum of the variances of the squared loadings, resulting in high loadings for a smaller number of regions and low loadings for the rest. Using orthogonal rotation such as varimax, the independence of the factors is preserved, whereas with oblique rotation, factors are allowed to correlate. Once factors are correlated, the interpretation of the factorial space becomes less obvious. Rotating the factors obliquely might give a better fit to data. However, orthogonal rotation has been recommended because of its simplicity and replicability (Rennie, 1997). To justify the use of orthogonal rotation in this study, we conducted oblique rotation and checked whether factor correlations exceed the threshold of 0.32 (Tabachnick and Fidell, 2007) indicating 10% or more overlap in variance among factors, enough variance to warrant oblique rotation. The regions with a loading greater than 0.6 were reported since a factor is regarded as reliable if it has four or more loadings of at least 0.6 regardless of sample size (Field, 2005; Guadagnoli and Velicer, 1988). The component (factor) scores of each retained component were calculated for individual subjects, which were then used for the correlation analyses with the total symptom severity scores and subscores of NSI and PCL-C.

Flowchart of the PCA and SEM performed in this study. In addition to the brain regions known to be involved in PTSD, those with high loadings from PCA after varimax rotation were included in the initial bottom–up model, and the resulting initial model is illustrated in Figure 2. PCA, principal component analysis; PTSD, post-traumatic stress disorder; SEM, structural equation modeling. Color images are available online.

Path diagrams of the initial bottom–up
Structural equation modeling
The SEM applies a hybrid technique that encompasses aspects of multiple regressions and confirmatory factor analysis (Yeh et al., 2010). The factors in the confirmatory factor analysis are latent variables representing shared information of indicator (observed) variables. Latent variables were scaled by fixing a factor loading to 1 for one of the indicator variables (Schweizer et al., 2019). Endogenous variables act as a dependent variable in at least one of the SEM equations, whereas exogenous variables are always independent variables in the SEM equations. In SEM, the relationships between variables represent the hypotheses regarding their influences, and they cannot be statistically tested for directionality.
In this study, SEM tests interrelationships among manifest (observed) and latent (unobserved) variables, represented by the metabolic activities of brain regions within the cortico-limbic circuits that are responsible for top–down control and bottom–up emotional processing. The SEM was applied separately to the two groups, that is, PTSD-present and PTSD-absent groups. Alternatively, one could apply a model to the entire population by including a grouping variable, but it would complicate building a model with the grouping variable. Moreover, it does not provide any information on individual paths for each group that we are interested in to compare differences in metabolic relationships between the two groups. Partial correlation coefficient matrices were calculated for each group and were used for SEM by using the lavaan package (Rosseel, 2012) in R software (R Core Team, 2020). We evaluated model fit with the standardized root mean square residual (SRMR), comparative fit index (CFI), and goodness of fit index (GFI). The general guidelines in SEM literature are that SRMR <0.05 indicates an excellent fit whereas <0.08 is acceptable; CFI and GFI >0.90 are acceptable, and >0.95 are excellent fit (Gunzler and Morris, 2015).
Bottom–up model
Figure 2 shows the initial bottom–up model that was built based on literature and PCA results. An exogenous latent variable, amygdala-hippocampus-insula (AHI), was constructed as an underlying cause of the metabolic activities of the amygdala, hippocampus, and insula during emotional processing. We tested a priori hypothesis that the latent AHI (exogenous variable) has an influence on other diverse brain regions (endogenous variables), including the rACC, vmPFC, precuneus, lingual, lateral occipital, and superior temporal cortices. Additional regions with high loadings (>0.8) from the PCA were included as endogenous variables in the initial bottom–up model. Endogenous variables not contributing to the model were removed from the initial bottom–up model by using two different methods (Fig. 1), resulting in two final candidate models: (1) by removing them iteratively evaluating three goodness-of-fit indices and (2) by using modification indices assessed for all paths in the initial model. For the first technique, at each iteration, the endogenous variables were deleted one at a time, and the three goodness-of-fit measures were computed. If two or more measures improved for both hemispheres by deleting the endogenous variable, the variable was eliminated from the model. Otherwise, it was retained. This was repeated sequentially for each endogenous variable and iteratively until no endogenous variables were removed in a given iteration. For the second technique, endogenous variables with modification indices less than 10.83 (p < 0.001) were removed. The modification index is the χ2 value with 1 degree of freedom by which model fit would improve if a particular path is added. The one yielding better goodness-of-fit indices between the two final candidate models determined through the two different approaches was chosen for further analysis.
Top–down model
We constructed two a priori models (Fig. 2) by postulating that the dlPFC influences the amygdala via rACC and vmPFC with and without the direct path from the dlPFC to the amygdala, and we evaluated model fits by using the goodness-of-fit indices to select the final top–down model. Since mediator variables were included in the top–down model, indirect and total effects of the dlPFC on the amygdala were assessed (Gunzler et al., 2013). The indirect effect is the effect of the exogenous variable on the endogenous variable that goes through a mediator and is calculated by multiplying all path coefficients of the path that constitutes the effect. The total effect is a sum of all direct and indirect effects.
Correlation and group comparison analyses
All the statistical analyses were performed by using R software. Partial correlation coefficients between symptom severity and component scores were calculated along with Bonferroni-corrected p-values after controlling for gender and age. Pearson's chi-squared tests were performed to compare two groups in gender and history of blast exposure. Two-sample t-tests or Mann–Whitney's U tests were performed to compare age, years of education, and time since injury, depending on results of the Shapiro–Wilk test for normality.
Two-sample t-tests with Bonferroni correction were applied to compare path coefficients and factor loadings between two groups, in which standard deviations were obtained from the standard error of a mean by multiplying by the square root of the sample size, and the pooled degrees of freedom was estimated by the Welch-Satterthwaite equation for unequal variance. Statistical significance was set at p < 0.05.
Results
Demographics and clinical characteristics
Differences between groups in demographics and clinical characteristics are summarized in Table 1. There was no significant difference in age, gender, time since injury, years of education, and history of blast exposure between the two groups.
PCA and correlation with symptoms
Six components that accounted for 87% of the total variance were retained after applying PCA to regional FDG uptake of all the subjects. None of the factor correlations exceeded 0.32 after oblique rotation (see the Supplementary Table S2 for details), indicating that the factors remained nearly orthogonal, which justifies the use of orthogonal rotation in our study. The first component that accounts for the largest variance (47.2%) comprised brain regions, including the amygdala, insula, hippocampus, superior temporal and entorhinal cortices, and fusiform. The second component comprised brain regions, including precentral, postcentral, and superior parietal cortices, and dlPFC. The third component comprised brain regions, including the thalamus, putamen, caudate, and pallidum. The regions and their loadings for the six components are shown in Table 2. These six components can be interpreted as brain circuitry systems, that is, the temporo-limbic, fronto-parietal, striato-thalamic, ventral PFC, occipital, and brainstem-cerebellar systems.
Names of Brain Regions and Their Loadings for the Six Principal Components Accounted for 87% of the Total Variance
The six principal components represent latent metabolic systems, that is, the TL, FP, ST, VP, OC, and BC systems, respectively.
BC, brainstem-cerebellar; FP, fronto-parietal; dACC, dorsal anterior cingulate cortex; dlPFC, dorsolateral prefrontal cortex; L, left; OC, occipital; PFC, prefrontal cortex; R, right; ST, striato-thalamic; TL, temporo-limbic; vlPFC, ventrolateral prefrontal cortex; vmPFC, ventromedial prefrontal cortex; VP, ventral prefrontal cortex.
The component score of the temporo-limbic system was positively correlated with both NSI and PCL-C total scores, as well as NSI somatosensory, affective, cognitive, and vestibular clusters, and PCL-C hyperarousal symptoms (Table 3), indicating that high FDG uptake in the temporo-limbic system is related to increased symptoms. No other systems showed significant correlations with any of the symptoms.
Partial Correlation Coefficients and Associated p-Values Between Each Pair of the Symptom Severity and Component Scores Controlling for Age and Gender for the Six Principal Components
The coefficients shown in bold represent significant correlations after Bonferroni correction.
Missing data: NSI total (15), somatosensory (9), affective (6), cognitive (4), and vestibular (7).
Bottom–up model
Figure 3 shows partial correlation coefficient matrices used in the SEM analyses. The regions with high loadings from PCA, including the precentral, postcentral, and superior parietal cortices, were included in the initial bottom–up model (Fig. 2). However, all of the regions were eliminated along with the vmPFC because of their insignificant contribution to the model according to the iterative method (see Supplementary Table S3 for details). Thus, the rACC, lingual, lateral occipital, superior temporal, and precuneus cortices were retained in the final bottom–up model. This final model (left: SRMR = 0.039, CFI = 0.95, GFI = 0.96; right: SRMR = 0.040, CFI = 0.92, GFI = 0.94) was found to outperform the one obtained by model selection by using modification index (left: SRMR = 0.092, CFI = 0.83, GFI = 0.88; right: SRMR = 0.071, CFI = 0.84, GFI = 0.87). Figure 4 shows the path coefficients and loadings of the final bottom–up model for each group. All the indicators (amygdala, insula, and hippocampus) were found to be in the first component (the temporo-limbic system) with high loadings. The bottom–up model met the criteria of all the goodness-of-fit indices for both the PTSD-absent group (left: SRMR = 0.046, CFI = 0.91, GFI = 0.95; right: SRMR = 0.046, CFI = 0.93, GFI = 0.96) and the PTSD-present group (left: SRMR = 0.049, CFI = 0.94, GFI = 0.94; right: SRMR = 0.045, CFI = 0.90, GFI = 0.90). The AHI was more strongly associated with the bilateral rACC (left: t = 5.4, corrected p < 0.001; right: t = 6.6, corrected p < 0.001), left lingual (t = 3.3, corrected p = 0.007), left superior temporal (t = 4.3, corrected p < 0.001), and right lateral occipital cortices (t = 4.4, corrected p < 0.001), but more weakly with the right precuneus (t = −5.0, corrected p < 0.001) in the PTSD-present group compared with the PTSD-absent group, suggesting that metabolic interrelationships in the cortico-limbic circuit were altered in subjects with significant PTSD symptoms. In addition, there were significant increases in the loadings of the amygdala (left: t = 6.8, corrected p < 0.001; right: t = 6.3, corrected p < 0.001) and the insula (left: t = 4.0, corrected p < 0.001; right: t = 4.7, corrected p < 0.001) on the AHI in the PTSD-present group compared with the PTSD-absent group, which suggest that the amygdala and insula have greater responses on the rACC, lingual, lateral occipital, precuneus, and superior temporal cortices in the PTSD-present group than the PTSD-absent group.
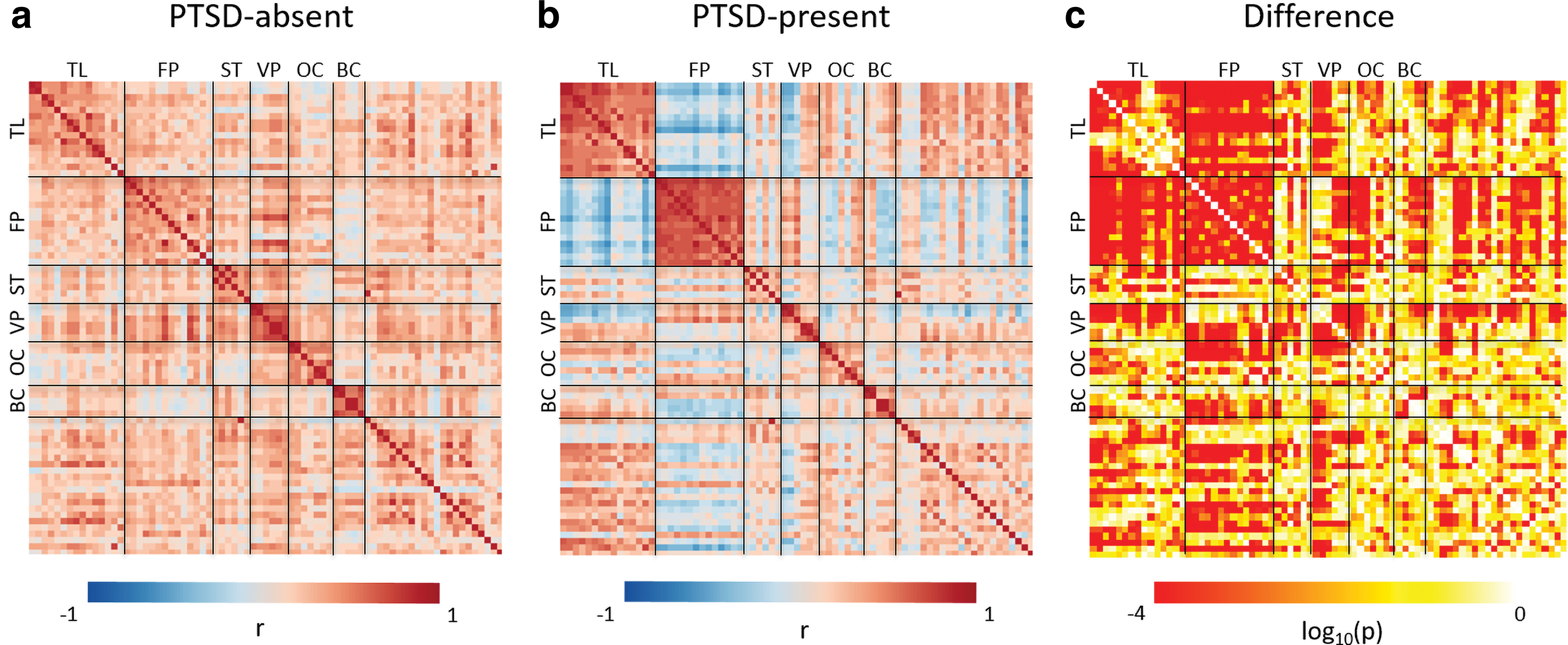
Partial correlation and p-value matrices. The partial correlation coefficients were computed between every pair of the 75 brain regions for PTSD-absent
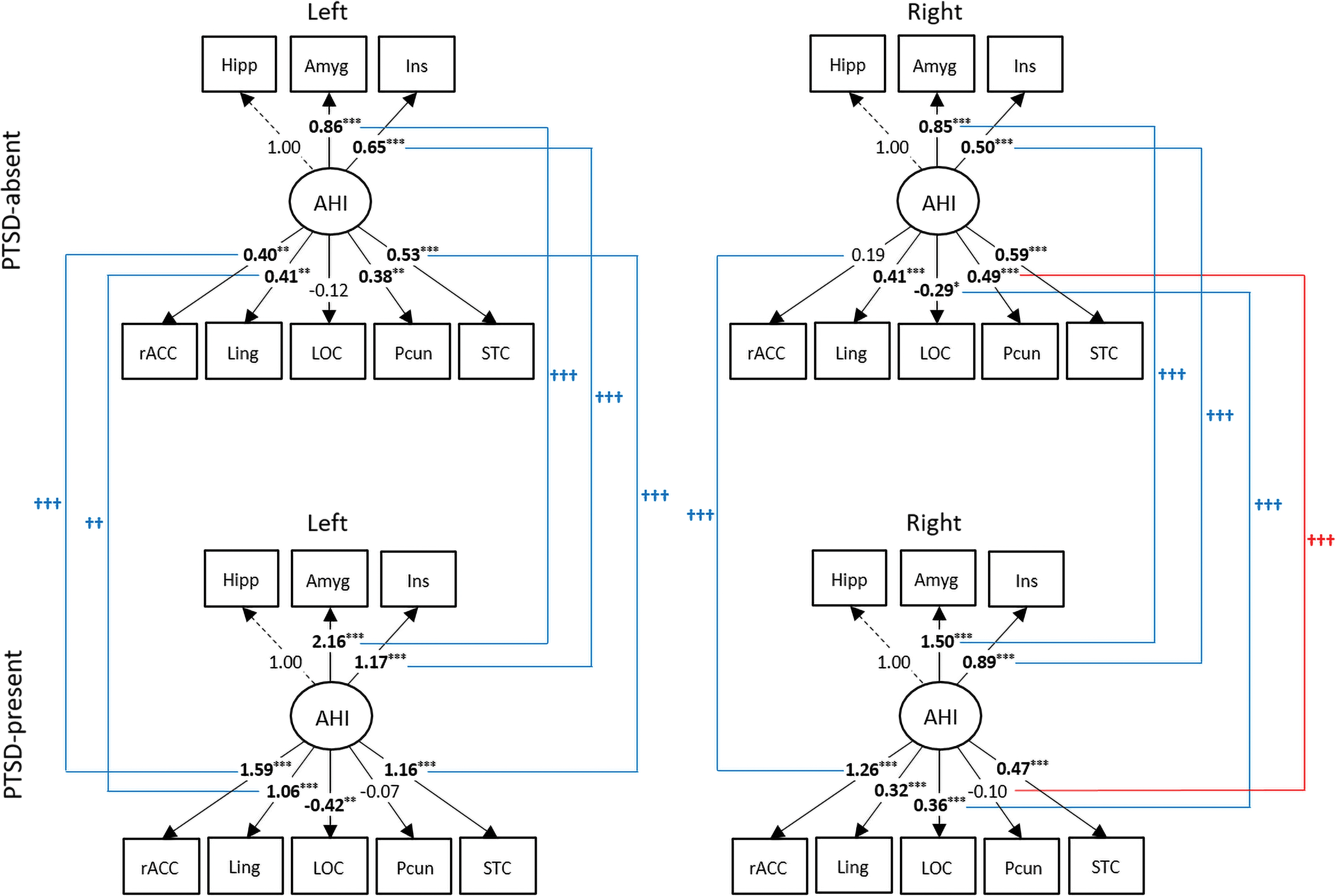
Path diagrams representing the bottom–up model for two mTBI groups. An ellipse-shaped variable represents a latent variable, and square-shaped variables represent manifest variables. Significant path coefficients and loadings are indicated by * (*p < 0.05, **p < 0.01, ***p < 0.001). Two-sample t-tests were performed to compare path coefficients and loadings between the two groups. The significant group differences (Bonferroni corrected p < 0.05) on estimated parameters are illustrated by † (†† p < 0.01, ††† p < 0.001) with different colors depending on whether parameter values increased (blue) or decreased (red). mTBI, mild traumatic brain injury. Color images are available online.
Top–down model
Goodness-of-fit indices for the top–down model without the direct path from the dlPFC to the amygdala were acceptable for the PTSD-absent group (left: SRMR = 0.052, CFI = 0.94, GFI = 0.98; right: SRMR = 0.032, CFI = 0.98, GFI = 0.99) but not for the PTSD-present group (left: SRMR = 0.191, CFI = 0.51, GFI = 0.76; right: SRMR = 0.190, CFI = 0.41, GFI = 0.72). The top–down model with the direct path from the dlPFC to the amygdala was found to fit better than the one without it; therefore, we retained the direct path in the final model (Fig. 5). With the final top–down model, the PTSD-absent group showed better goodness-of-fit (left: SRMR = 0.025, CFI = 0.99, GFI = 1.00; right: SRMR = 0.001, CFI = 1.00, GFI = 1.00) compared with the PTSD-present group (left: SRMR = 0.131, CFI = 0.88, GFI = 0.93; right: SRMR = 0.075, CFI = 0.97, GFI = 0.98). Compared with the PTSD-absent group, the dlPFC in the PTSD-present group was less positively associated with the vmPFC (left: t = −12.1, corrected p < 0.001; right: t = −10.8, corrected p < 0.001) and more negatively associated with the amygdala in the right hemisphere (t = −4.4, corrected p < 0.001). Contrary to the PTSD-absent group, the PTSD-present group had a negative association between the dlPFC and rACC (left: t = −6.3, corrected p < 0.001; right: t = −5.0, corrected p < 0.001). Unlike the vmPFC which had no significant difference in its association with the amygdala, the rACC was more strongly associated with it (left: t = 4.5, corrected p < 0.001; right: t = 5.9, corrected p < 0.001) in the PTSD-present group, compared with the PTSD-absent group.
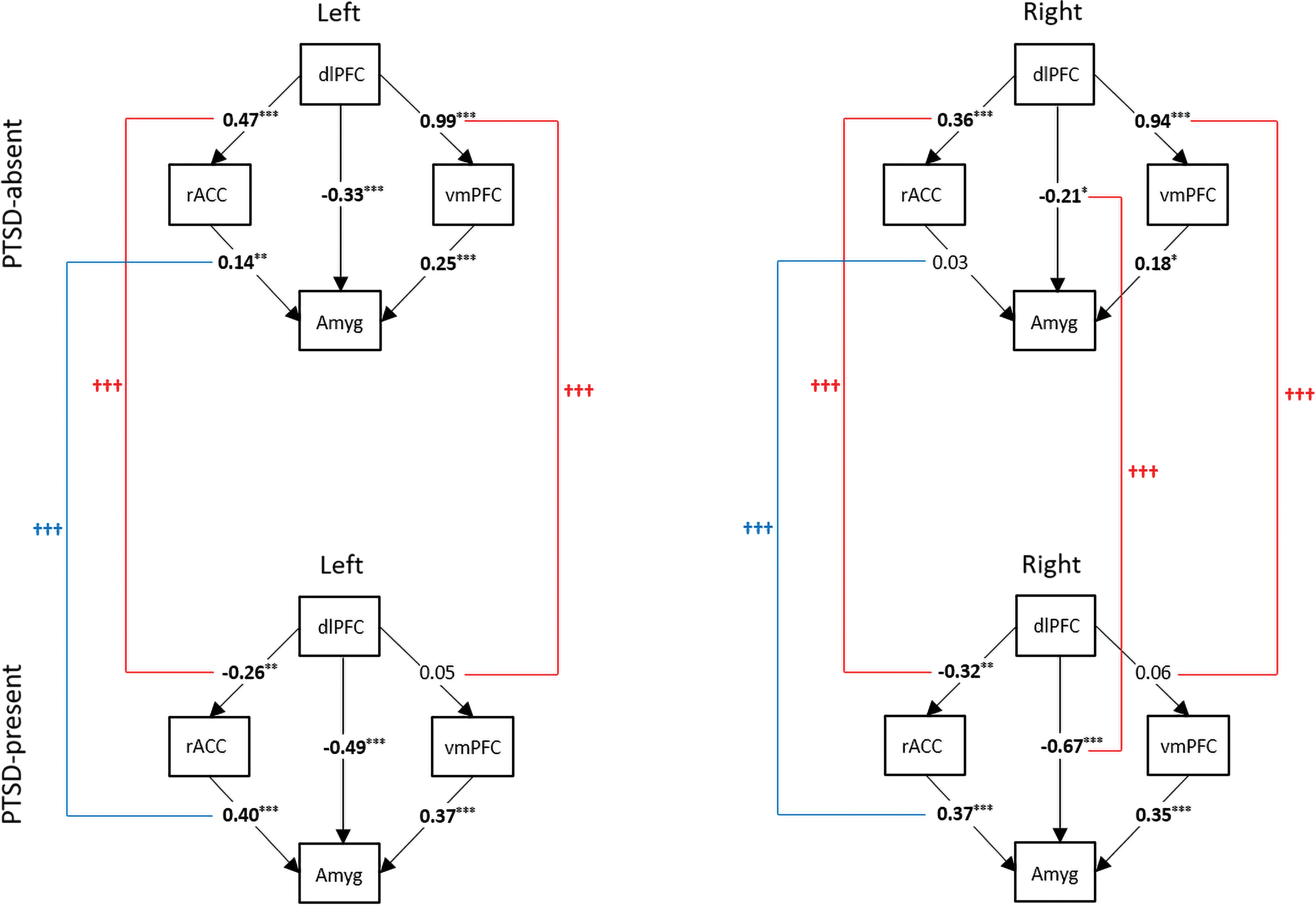
Path diagrams representing the top–down model for two mTBI groups. Square-shaped variables represent manifest variables. Significant path coefficients are indicated by * (*p < 0.05, **p < 0.01, ***p < 0.001). Two-sample t-tests were performed to compare path coefficients between the two groups. The significant group differences (Bonferroni corrected p < 0.05) on estimated parameters are illustrated by † (††† p < 0.001) with different colors depending on whether parameter values increased (blue) or decreased (red). Color images are available online.
The indirect effect of the dlPFC on the amygdala via the rACC was positive (left: B = 0.07, SE = 0.02, p = 0.007; right: B = 0.01, SE = 0.02, p = 0.489) in the PTSD-absent group, but negative (left: B = −0.10, SE = 0.03, p = 0.002; right: B = −0.12, SE = 0.04, p = 0.001) in the PTSD-present group, which significantly reduced the indirect effect (left: t = −4.1, corrected p < 0.001; right: t = −3.2, corrected p = 0.013). On the other hand, the indirect effect of the dlPFC on the amygdala via the vmPFC was positive (left: B = 0.25, SE = 0.07, p < 0.001; right: B = 0.17, SE = 0.07, p = 0.018) in the PTSD-absent group, but not significant in the PTSD-present group, significantly reducing the indirect effect in the left hemisphere (t = −3.3, corrected p = 0.009). Overall, the total effect of the dlPFC on the amygdala was not significant in the PTSD-absent group, but was negative in the PTSD-present group (left: B = −0.58, SE = 0.06, p < 0.001; right: B = −0.77, SE = 0.06, p < 0.001). The total effect of the dlPFC on the amygdala was significantly decreased in both hemispheres (left: t = −6.8, corrected p < 0.001; right: t = −8.5, corrected p < 0.001) in the PTSD-present group compared with the PTSD-absent group.
Discussion
We found significant associations between the self-reported symptoms and the regional FDG uptake in the temporo-limbic system. In addition, the results of SEM demonstrated altered metabolic interrelationships within the neural circuits involving the bottom–up emotional processing and top–down control in the PTSD-present group. For the bottom–up emotional processing, the AHI in the PTSD-present group had stronger interrelationships with the bilateral rACC, left lingual, left superior temporal, and right lateral occipital cortices, but a weaker interrelationship with the right precuneus cortex, compared with the PTSD-absent group. For the top–down control, the PTSD-present group had decreased metabolic engagements of the dlPFC on the amygdala. These findings suggest impaired bottom–up emotional processing and top–down control in the cortico-limbic circuit in comorbid mTBI and PTSD.
Correlation between FDG uptake and self-reported symptoms
Using PCA, we identified latent metabolic systems of the brain regions based on intrinsic features of the regional FDG uptake, that is, the temporo-limbic, fronto-parietal, striato-thalamic, ventral PFC, occipital, and brainstem-cerebellar systems. The FDG uptake in the temporo-limbic system was found to be positively correlated with the severity of post-concussive and hyperarousal symptoms. Of note, three of the five hyperarousal symptoms on the PCL-C (i.e., sleep disturbance, irritability, difficulty concentrating) highly overlapped with post-concussive symptoms (Table 3). Given no other PTSD clusters and all NSI clusters were correlated with the temporo-limbic system, the FDG uptake of the temporo-limbic system may be associated with increased symptom reporting generally, rather than symptoms specific to PTSD or mTBI. Post-concussion symptoms are nonspecific (Iverson et al., 2013; McCrae, 2008) and regularly occur in healthy individuals (Iverson, 2005). Further, post-concussive symptoms have consistently been shown to be most strongly predicted by PTSD symptoms (French et al., 2014; Hoge et al., 2008; Lange et al., 2020a,b; Pietrzak et al., 2009; Schneiderman et al., 2008). Indeed, in our sample, PTSD and neurobehavioral symptoms were highly collinear (PCL-C total and NSI total, r = 0.79). Unfortunately, this prevented us from including neurobehavioral symptoms in the models to better parse out whether these relationships were truly specific to PTSD or instead indicative of psychological distress more generally.
Bottom–up emotional processing
The amygdala, insula, hippocampus, and superior temporal cortex in the bottom–up model were among the regions with high loadings in the first component (the temporo-limbic system) of the PCA results. On the contrary, those with high loadings in the second component (the fronto-parietal system), which were initially included in the bottom–up model, did not significantly contribute to the model. Instead, we found that the regions of the occipital system were more critical in the bottom–up model, which suggests that the temporo-limbic system is more closely associated with the occipital system than the fronto-parietal system. These findings suggest that altered metabolic interrelationships in the occipito-temporo-limbic circuit play an important role in the emotional processing of military mTBI patients with significant PTSD symptoms at rest.
Although dACC is responsible for executive functions by influencing multiple cognitive processes, rACC is involved in emotion experience and processing (Bush et al., 2000). In PTSD, the amygdala has been widely reported to be hyperactive, likely resulting from dysfunction of the top–down inhibitory control; the insula has also been found to be hyperactive, with greater positive connectivity with the amygdala (Sripada et al., 2012), but findings regarding hippocampal activity are mixed (Hughes and Shin, 2011). This study found that the positive association between the AHI and the rACC significantly increased in the PTSD-present group compared with the PTSD-absent group. On the contrary, a PET study found hypoactive rACC in PTSD patients with comorbid cocaine and alcohol abuse during rest (Semple et al., 2000). However, excessive rACC activity has been associated with symptom severity in depressive disorder (Matthews et al., 2009). Further, rACC has been a target of deep brain stimulation, and inhibiting rACC activity has been shown to be effective in treating patients with treatment-resistant depressive disorder (Mayberg et al., 2005).
The patients with PTSD often feel overwhelmed by multiple and simultaneous sensory stimuli (Stewart and White, 2008). Hyperactivity of the amygdala likely drives enhancement of visual perceptual processing as well as enlargement of visual cortex representation of trauma-related stimuli, which can lead to hypervigilance, visual intrusions, and visual flashbacks/hallucinations (Rangaprakash et al., 2019). This study found stronger interrelationships between the AHI and visual cortices, including the left lingual and the right lateral occipital cortices. Consistent with our findings, a neuroimaging study found increased activation in the visual cortex (V1/V2) at rest in patients with PTSD compared with controls (Zhu et al., 2014). The increased activation of the visual cortices may not be dampened due to disrupted sensory filtering or problems with suppressing irrelevant environmental sensory stimuli. Interestingly, studies with patients with PTSD have reported cortical thinning in the left lingual gyrus (Wrocklage et al., 2017) and right lateral occipital cortex (Qi et al., 2013; Wrocklage et al., 2017) and reduced gray matter volume in the lingual gyrus (Nardo et al., 2013). The volume differences may be related to hyper-activation of the visual cortical regions.
The superior temporal cortex was loaded strongly with the temporo-limbic system along with the amygdala, hippocampus, and insula, suggesting a strong association with those limbic structures. Moreover, our results of SEM found an increased positive association between the AHI and the superior temporal cortex in the left hemisphere in the PTSD-present group. Contrary to the findings of reduced volume in the visual cortices, a study in pediatric PTSD found that volumes of both left and right superior temporal gyri were larger in PTSD subjects compared with control subjects (De Bellis et al., 2002). The increased volume in the superior temporal gyrus may result from morphological compensatory plasticity in acquired strong visual stimulation (Noppeney et al., 2005).
The precuneus is involved in autobiographical memory and is related to self-referential processing (Cavanna and Trimble, 2006; Kelley et al., 2002). Thus, alterations in the precuneus activity may potentially be related to altered memory and self-referential processes such as memory deficits, intrusions, or flashbacks. Neuronal activities of the precuneus have been shown to be related to trauma memory generalization (Hayes et al., 2011) and flashbacks (Whalley et al., 2013). In this study, we found that unlike the other regions in the bottom–up model, the PTSD-present group had a significantly reduced interrelationship between the AHI and the precuneus in the right hemisphere compared with the PTSD-absent group. A resting-state fMRI study found that decreased activity in the precuneus correlates with more severe reexperiencing symptoms (Yan et al., 2013). Another study demonstrated that eye movement desensitization and reprocessing therapy can increase metabolic activities of the precuneus, which was correlated with a reduction of PTSD symptoms (Rousseau et al., 2019).
Top–down control
The dlPFC has long been thought to be critically involved in cognitive control (Aupperle et al., 2012; Connolly et al., 2017; Davidson et al., 2000; Urry et al., 2006). Compared with the PTSD-absent group, our findings of a less engaged metabolic interrelationship between the dlPFC and amygdala in the PTSD-present group suggest impaired top–down regulation in comorbid mTBI and PTSD. The PTSD may impact the frontal regions associated with cognitive and executive functioning, which may result in disruption of cognitively regulating emotional responses to aversive events. Studies have suggested that the dlPFC could be pivotal in the conscious regulation of emotion to reduce fear responses (Phelps et al., 2004; Phillips et al., 2008) and the recovery from PTSD (Rauch et al., 2006). Resting-state fMRI studies have reported that patients with PTSD had reduced functional connectivity between the dlPFC and amygdala compared with healthy controls (Chen et al., 2018). The reduced connectivity between the two regions has also been reported in patients with depressive and anxiety disorders (Liu et al., 2015; Ochsner et al., 2002; Satterthwaite et al., 2016). Further, this study found that the dlPFC had a significant direct effect on the amygdala in both hemispheres. In addition to the indirect effects via the vmPFC and rACC, the dlPFC might influence the amygdala by modulating other brain regions such as posterior areas of perceptual processing (i.e., occipital/parietal cortex) (Ochsner and Gross, 2005; Ochsner et al., 2002) since the dlPFC is unlikely to directly influence the amygdala due to their weak connections (Goetschius et al., 2019; McDonald, 1998).
The rACC located between the neocortex and the limbic system is also a key brain region for the regulation of emotion. Resting-state fMRI studies have found that patients with PTSD show more positive or less anticorrelated functional connectivity between the amygdala and rACC (Birn et al., 2014; Brown et al., 2014; Sripada et al., 2012). Consistent with the previous studies, this study found increased engagement of the rACC with the amygdala in the PTSD-present group whereas the dlPFC had significantly reduced influence on the amygdala.
In addition, the vmPFC has been known to regulate emotional responses by controlling the functions of the amygdala (Connolly et al., 2017; Davidson et al., 2000; Koenigs and Grafman, 2009; Urry et al., 2006). The dlPFC inhibition of the amygdala during cognitive regulation appears to be mediated through the vmPFC (Ghashghaei et al., 2007; Lobo et al., 2011; Phillips et al., 2008) that has dense connections with the amygdala (Goetschius et al., 2019; McDonald, 1998). Examination of functional connectivity at rest has shown decreased coupling between the amygdala and vmPFC in PTSD (Sripada et al., 2012). A study of service members with comorbid mTBI and PTSD using diffusion tensor imaging (DTI) tractography (Santhanam et al., 2019a) has demonstrated damaged uncinate fasciculus, a major white matter fasciculus inter-connecting the amygdala and mPFC (Von Der Heide et al., 2013). Another DTI study has shown a negative correlation between the integrity (fractional anisotropy) of the uncinate fasciculus and PTSD symptoms in service members (Costanzo et al., 2016), suggesting that alterations in effective connectivity between the amygdala and mPFC may have a structural basis of comorbid mTBI and PTSD. Even though the current study with mTBI patients found that the metabolic interrelationship between the amygdala and the vmPFC does not significantly differ between the PTSD-present and the PTSD-absent groups, our finding of a decreased association between the dlPFC and the amygdala via the vmPFC in the PTSD-present group suggests decreased inhibition of the amygdala by the dlPFC via the dlPFC-vmPFC-amygdala pathway and may lead to impaired emotion control.
Limitations and future work
The findings presented in this analysis should be considered within the context of the limitations of the study design. First, the assessment of the PTSD was based on self-reported symptom severity (PCL-C) in the current study, rather than diagnosed by a clinician. Future studies should employ clinician-administered interviews, such as the Clinician-Administered PTSD scale (CAPS), to better characterize PTSD symptoms. Another limitation of the study was the lack of service members without mTBI, precluding the elucidation of metabolic interrelationships primarily attributed to mTBI.
Nevertheless, the findings suggest that, in individuals with mTBI, comorbid significant PTSD symptoms were associated with altered FDG uptake. Our findings lay the groundwork for future studies using additional neuroimaging techniques to investigate associations between the networks within this analysis and PTSD symptoms. For instance, future work evaluating the transition from non-stress (PFC regulation) to stress conditions (amygdala control) by further elucidating stress signaling pathways using task-based functional neuroimaging may advance this line of research in comorbid mTBI and PTSD.
Conclusions
We found significant associations between FDG uptake in the temporo-limbic system and post-concussive and hyperarousal symptoms in mTBI subjects. The PTSD-present group showed altered interrelationships in FDG uptake between brain regions in the cortico-limbic circuitry of the bottom–up emotional processing and top–down control. The altered patterns of the metabolic interrelationships may be associated with severity of PTSD symptoms in mTBI subjects. Our findings have potential applications for a better understanding of the underlying brain pathophysiological alterations in comorbid mTBI and PTSD, which could lead to important developments for future interventions.
Footnotes
Acknowledgments
The authors would like to thank Dr. Theresa Woo of the WRNMMC/NICoE for her valuable comments, and the 15-year Longitudinal TBI Study team of the Defense and Veterans Brain Injury Center for recruiting, screening, scheduling, and arranging travel for a portion of the participants in this study.
Authors' Contributions
C.S. contributed to study design, data processing and analyses, and interpretation of results and wrote the article. P.-H.Y. contributed to study design, critical review, and interpretation of results. J.O. contributed to critical review. C.S.R. and S.M.L. contributed to interpretation of results. G.R. supervised the study. G.H.B. supervised data acquisition and analyses. All authors contributed to revision of the article and approved the submitted version.
Author Disclosure Statement
No competing financial interests exist. The views expressed in this article are those of the authors and do not reflect the official policy of the Department of Army, Navy, Air Force, Department of Defense, or U.S. Government. The identification of specific products, scientific instrumentation, or organizations is considered an integral part of the scientific endeavor and does not constitute endorsement or implied endorsement on the part of the author, DoD, or any component agency.
Funding Information
This research was supported by CNRM grant #300606 and CDMRP grant PT074437.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.