
Abstract
Background:
Tractography based on diffusion-weighted magnetic resonance imaging (DWI) models the structural connectivity of the human brain. Deep brain stimulation (DBS) targeting the subthalamic nucleus is an effective treatment for advanced Parkinson's disease, but may induce adverse effects. This study investigated the relationship between structural connectivity patterns of DBS electrodes and stimulation-induced side effects.
Materials and Methods:
Twenty-one patients with Parkinson's disease treated with bilateral subthalamic DBS were examined. Overall, 168 electrode contacts were categorized as inducing or noninducing depending on their capability for inducing side effects such as motor effects, paresthesia, dysarthria, oculomotor effects, hyperkinesia, and other complications as assessed during the initial programming session. Furthermore, the connectivity of each contact with target regions was evaluated by probabilistic tractography based on DWI. Finally, stimulation sites and structural connectivity patterns of inducing and noninducing contacts were compared.
Results:
Inducing contacts differed across the various side effects and from those mitigating Parkinson's symptoms. Although contacts showed a largely overlapping spatial distribution within the subthalamic region, they could be distinguished by their connectivity patterns. In particular, inducing contacts were more likely connected with supplementary motor areas (hyperkinesia, dysarthria), frontal cortex (oculomotor), fibers of the internal capsule (paresthesia), and the basal ganglia-thalamo-cortical circuitry (dysarthria).
Discussion:
Side effects induced by DBS seem to be associated with distinct connectivity patterns. Cerebellar connections are hardly associated with side effects, although they seem relevant for mitigating motor symptoms in Parkinson's disease. A symptom-specific, connectivity-based approach for target planning in DBS may enhance treatment outcomes and reduce adverse effects.
Impact statement
Tractography based on diffusion-weighted magnetic resonance imaging has become a prominent technique for investigating the connectivity of human brain networks in vivo. However, the relationship between structural connections and brain function is still hardly known. The present study examined the relationship between adverse behavioral effects induced by deep brain stimulation (DBS) and tractography patterns in individual brains. The results suggest that DBS-based side effects depend on the structural connections of electrode contacts rather than their location. Network-based target planning in DBS may improve treatment by avoiding side effects. Moreover, the adopted approach may serve as a paragon for investigating structure/function relationships.
Introduction
Subthalamic nucleus (STN) deep brain stimulation (DBS) substantially mitigates symptoms of Parkinson's disease (Deuschl et al., 2006; Wodarg et al., 2012). Together with the internal globus pallidus (GPi), the STN represents the major target structure in DBS for this condition. However, STN-DBS is associated with several relevant complications. Besides acute surgery-related complications such as wound infections, hemorrhage, and seizures (Boviatsis et al., 2010), and direct stimulation-associated motor (e.g., muscle contractions, dyskinesias) and nonmotor symptoms (e.g., paresthesia, acute neuropsychiatric decompensation, autonomous side effects) may occur (Baizabal-Carvallo and Jankovic, 2016; Zheng et al., 2010). Long-term side effects, including motor (e.g., spasticity, dysarthria, gait disorder, dyskinesia, hypotonia), psychiatric (e.g., depression, suicidality, cognitive dysfunction, agitation, mania, emotional changes), and technical complications (e.g., lead breakage, lead migration, infection), may also occur (Koeglsperger et al., 2019; Witt et al., 2008). Most studies assume adverse effects being primarily a function of electrode location (e.g., STN division) or stimulation properties (e.g., amplitude).
Tractography based on diffusion-weighted magnetic resonance imaging (DWI) has become a prominent technique for investigating the connectivity of human brain networks in vivo. In recent years, several studies adopted tractography approaches to examine the connectivity patterns of DBS electrodes that mitigate motor symptoms (Accolla et al., 2016; Akram et al., 2017; Horn et al., 2017; Strotzer et al., 2019; Vanegas-Arroyave et al., 2016). However, only marginal reports are available regarding the relationship between DBS-induced side effects and the connectivity patterns of the electrodes (Akram et al., 2017; Mahlknecht et al., 2017; Prent et al., 2019). Therefore, this study aimed to investigate the location and structural connectivity of DBS contacts associated with side effects in STN-DBS for Parkinson's disease. This information may improve target planning in DBS surgery by avoiding targets associated with adverse effects and help to understand the structure/function relationship of other neurological conditions. A patient-specific (rather than normative) approach (Wang et al., 2020) using probabilistic tractography (rather than deterministic) (Schlaier et al., 2017) was adopted as it tends to be most sensitive for stimulation-dependent connectivity patterns.
Materials and Methods
Patients
Twenty-one Parkinson's patients who received bilateral STN-DBS between 2013 and 2017 were included in this monocenter retrospective study. Idiopathic Parkinson's disease was diagnosed based on the U.K. brain bank criteria (Hughes et al., 1992), and eligibility for DBS surgery was evaluated using a standardized preoperative assessment (Hilker et al., 2009). Exclusion criteria for surgery were age (>75 years), dementia, severe neuropsychiatric impairments, lack of compliance, coagulation disorders, immunosuppression, severe cerebral atrophy, and cerebral macroangiopathy.
Patient-awake surgeries were performed in all cases. Target points for DBS electrodes were selected preoperatively in iPlanNet Stereotaxy (v3.0; Brainlab, Munich, Germany) using T1- and T2-weighted magnetic resonance imaging (MRI) coregistered to stereotactic computed tomography (CT) scans. Intraoperative verification included electrophysiological microelectrode recordings and testing by experienced movement disorder specialists.
The study was approved by the University Medical Center Ethics Committee and conducted according to the Declaration of Helsinki. All data were acquired during the standard routine in DBS surgery. Part of the data regarding Parkinson's symptom alleviation and dysarthria were reported in a recent study (Strotzer et al., 2019). The present work applied similar methods for data preparation and processing.
Image acquisition
MRI was acquired at a 1.5T Avanto from nine patients and a 3T Skyra scanner (both Siemens, Erlangen, Germany) from 12 patients before surgery. General anesthesia was applied to reduce movement artifacts. Images were recorded using a 12-channel head coil (Avanto) or a 20-channel head-neck coil (Skyra). The imaging protocol included structural sagittal T1-weighted, and axial and sagittal T2-weighted acquisitions for trajectory planning and DWI for probabilistic tractography.
T1-weighted images were acquired by a 3D magnetization-prepared rapid-acquisition gradient-echo sequence (1.5T: repetition time, TR: 1680 ms; echo time, TE: 2.97 ms, flip angle, FA: 15°, field of view, FOV: 240 × 240 mm2, voxel size: 0.90 × 0.90 × 1.10 mm3, 160 slices; 3T: TR: 1910 ms, TE: 3.67 ms, FA: 9°, FOV: 250 × 250 mm2, voxel size: 0.98 × 0.98 × 1.00 mm3, 160 slices). Sagittal T2-weighted images were acquired by a turbo spin-echo sequence with a slice gap of 0.2 mm (1.5T: TR: 3030 ms, TE: 102 ms, FA: 180°, FOV: 230 × 230 mm2, voxel size: 0.90 × 0.72 × 2.00 mm3, 15 slices; 3T: TR: 5320 ms, TE: 87 ms, FA: 150°, FOV: 230 × 230 mm2, voxel size: 0.64 × 0.51 × 2.00 mm3, 23 slices). The 40 axial T2-weighted slices were positioned parallel to the intercommissural plane. On the 1.5T scanner, they were acquired by a spin-echo sequence (TR: 4070 ms, TE: 80 ms, FA: 90°, FOV: 250 × 250 mm2, voxel size: 0.98 × 0.98 × 2.00 mm3) and on the 3T scanner by a turbo spin-echo sequence (TR: 5170 ms, TE: 67 ms, FA: 150°, FOV: 210 × 210 mm2, voxel size: 0.82 × 0.82 × 2.00 mm3). DWI was acquired with a single-shot spin-echo sequence with echo-planar readout along 64 axial slices (1.5T: TR: 5700 ms, TE: 98 ms, FA: 90°, FOV: 230 × 230 mm2, voxel size: 1.80 × 1.80 × 3.00 mm3; 3T: TR: 10,800 ms, TE: 82 ms, FA: 90°, FOV: 256 × 256 mm2, voxel size: 2.00 × 2.00 × 2.00 mm3). Diffusion was probed across 64 (20 patients) or 30 (one patient) gradient directions with a b-value of 1000 s/mm2. In addition, a single pair of b-zero (without diffusion-weighting) volumes with opposing phase-encoded readout were acquired to estimate the susceptibility field. A gadobutrol contrast agent was administered before the T1-weighted acquisition. Interference of the agent with other acquisitions was avoided by acquiring T1-weighted images at the end.
Before surgery, CT was performed with a mounted stereotactic frame (Radionics CRW; Burlington, US) for surgery planning on a Somatom Definition Flash scanner (Siemens). Postoperative imaging included CT with 1 mm slice thickness to verify surgery results.
Fiber tracking
Coordinates of each contact of the quadripolar DBS electrodes were manually extracted from the postoperative CT scans using iPlanNet Stereotaxy (v3.0; Brainlab). The probabilistic tracking algorithm was seeded by a volume of 3 × 3 × 1 voxels of the axial T2-weighted images (about 15 mm3) around the center of every contact. Overall, 168 connectivity profiles (4 contacts per hemisphere, 21 patients) were computed.
DWI preprocessing and fiber tracking were performed with the Diffusion Toolbox FDT (v5.0.9; Centre for Functional Magnetic Resonance Imaging of the Brain, Oxford, United Kingdom) (Behrens et al., 2007) following procedures as described before (Strotzer et al., 2019). In brief, DWI was corrected for susceptibility artifacts with TOPUP (using acquisitions with opposing polarities of the phase-encode blips) and eddy current distortions using EDDY and coregistered to the T1-weighted and T2-weighted images. Voxel-wise distributions of fiber orientations were estimated based on the Markov Chain Monte Carlo sampling with BEDPOSTX (2 fibers per voxel, weight: 1, burn-in period: 1000 iterations, jumps: 1250, sampled every 25 steps). Probabilistic tracking was seeded from the contact locations and performed with PROBTRACKX (5000 streamlines per voxel, maximum step number: 2000, step length: 0.5 mm, subsidiary fiber volume threshold: 0.01, curvature threshold: 80°, no anisotropy threshold). The resulting track frequency maps were transformed into log-scaled track probability maps (Smith et al., 2018) and thresholded at 0.5.
Subsequently, track probability maps were superimposed on patients' individual axial T2-weighted slices, and every electrode contact was separately evaluated by its connections with predefined subcortical and cortical target areas. The presence and absence of connections were binary encoded and noted in a standardized protocol. Structures were visually identified in individual brains guided by stereotactic/neuroanatomical atlases (Morel, 2007; Nieuwenhuys et al., 2008; Schaltenbrand et al., 2005). Inferior frontal gyrus (IFG) refers to Brodmann areas 45/46.
Clinical testing
Testing was performed separately for every electrode contact by experienced movement disorder specialists in an initial programming session 1 month after surgery. The baseline was reestablished between testing. The stimulation mode was set to monopolar. Frequency (130 Hz) and pulse width (60 microseconds) were fixed. Stimulation amplitude was gradually increased (up to a maximum of 7.0 V). Following the Unified Parkinson's Disease Rating Scale, effects on contralateral bradykinesia, rigidity, and tremor were noted in a standardized protocol. Furthermore, the occurrence of direct stimulation-induced side effects and the associated threshold amplitudes were assessed. This included dysarthric, motor (e.g., phasic movements, facial and limb muscle contractions, paresis, conjugate eye deviation), and oculomotor (unilateral eye deviation) effects, paresthesia, and hyperkinesia (tonic movements). Autonomous side effects and other unspecified complications (e.g., dizziness, tiredness, restlessness, head and neck pressure) were categorized as other side effects. Only acute stimulation-associated symptoms were investigated. Assessments were performed in a medication OFF state.
Statistical analysis
Electrode contacts were classified into inducing contacts (stimulation-elicited side effects) and noninducing contacts (no effect elicited). Connectivity with target areas was encoded in a binary manner. Demographic data, clinical data, and electrode contact coordinates (relative to AC/PC) were evaluated by descriptive statistics and t-tests. Normal distribution was assessed by Shapiro–Wilk tests. The relationships between side effects (co-occurrence) were evaluated with 2 × 2 contingency coefficients (phi). These were tested for independence by two-tailed chi-squared tests. Fisher's exact tests were used when >20% of cells had frequencies <5.
Connectivity patterns for each side effect were analyzed in two steps. First, connectivity patterns (frequency distributions of connections between inducing or noninducing contacts and target structures) were compared for each pair of side effects with a 2 × k chi-squared test for homogeneity. We reasoned that side effects with the same connectivity pattern should not deviate in their frequency distribution. A deviation from homogeneity suggests that at least some connections differ. To reveal which connections differed, subsequent analyses evaluated for each target structure the relationship between side effect induction and connectedness with 2 × 2 contingency coefficients (phi). These contingencies were tested by chi-squared tests (or Fisher's exact tests). The type I error was set at p ≤ 0.05 (uncorrected). To rule out possible confounds by the two different acquisition protocols (1.5T vs. 3T), all analyses were repeated by agglutination tests (Cochran, 1954) (assuming separate samples per acquisition protocol). These agglutination tests provided very similar results as the chi-squared tests for the combined sample and, hence, are not reported in detail. Nevertheless, those cases showing minor differences are reported.
Results
General
Five of the twenty-one patients were female. The mean age was 60.0 years with a standard deviation (SD) of 6.9. The mean disease duration was 11.4 years (SD: 5.6). Ten patients were rigid-akinetic, nine showed an equivalent disease type, and two were tremor-dominant. The daily L-dopa equivalent dose before surgery averaged at 1039.0 mg (SD: 399.0). The Unified Parkinson's Disease Rating Scale part III (maximum: 108) was determined in L-dopa ON (mean: 14.9, SD: 7.7) and OFF (mean: 40.1, SD: 9.4) state before surgery. For 16 patients, the daily dosage of dopaminergic medication 12 months after surgery was available and was reduced significantly (p < 0.001) by 57.3% from a mean dose of 1002.2 mg/day (SD: 427.8) to 428.1 mg/day (SD: 210.5).
Frequency and co-occurrence of side effects
Motor complications were the most frequently observed immediate adverse effects (32.1% of all electrode contacts), closely followed by paresthesia (30.4% of contacts). See Table 1 for details. Despite these relatively large occurrence rates, there was no significant positive contingency between side effects as expressed by phi coefficients (Table 2). For instance, among the 54 contacts inducing motor effects, only 18 (33.3%) also induced paresthesia. Among the 118 contacts not inducing motor effects, 33 (29.0%) caused paresthesia resulting in an almost zero phi coefficient (phi = 0.045, not significant). We only observed significant negative contingencies, for instance, oculomotor effects were less likely induced by contacts leading to motor effects (phi = −0.163, p = 0.035). Table 2 also lists contingencies between side effect induction and effectiveness in reducing Parkinson's symptoms (adopted from (Strotzer et al., 2019)). Except for the relationship between bradykinesia reduction and dysarthria induction, there was no significant contingency.
Investigated Side Effects
This table lists the number (and percentage) of electrode contacts inducing side effects. Also, the threshold stimulation amplitudes at which the side effects occurred are listed.
SD, standard deviation.
Co-Occurrence of Side Effects and Symptom Reduction
The table lists pairwise contingency coefficients (phi) for the co-occurrence of side effects by the inducing electrode contacts. Positive phi: both side effects are induced/noninduced by the same contacts. Negative phi: side effects are induced/noninduced by different contacts. The bottom of the list shows pairwise contingency coefficients for the co-occurrence of side effects with symptom reduction [adopted from (Strotzer et al., 2019)]. Results of chi-squared or exact tests (p-values) are provided in parenthesis. n = 168 (except when otherwise stated). *≤0.05.
Contact locations
Although the side effects seemed relatively unrelated, coordinates of inducing electrode contacts (relative to the patient-specific midpoint between anterior and posterior commissure) overlapped. Only minor (nonsignificant) differences in the mean contact location were observed across side effects (except when otherwise reported below). Contact coordinates are listed in Table 3 and illustrated in Figure 1A.
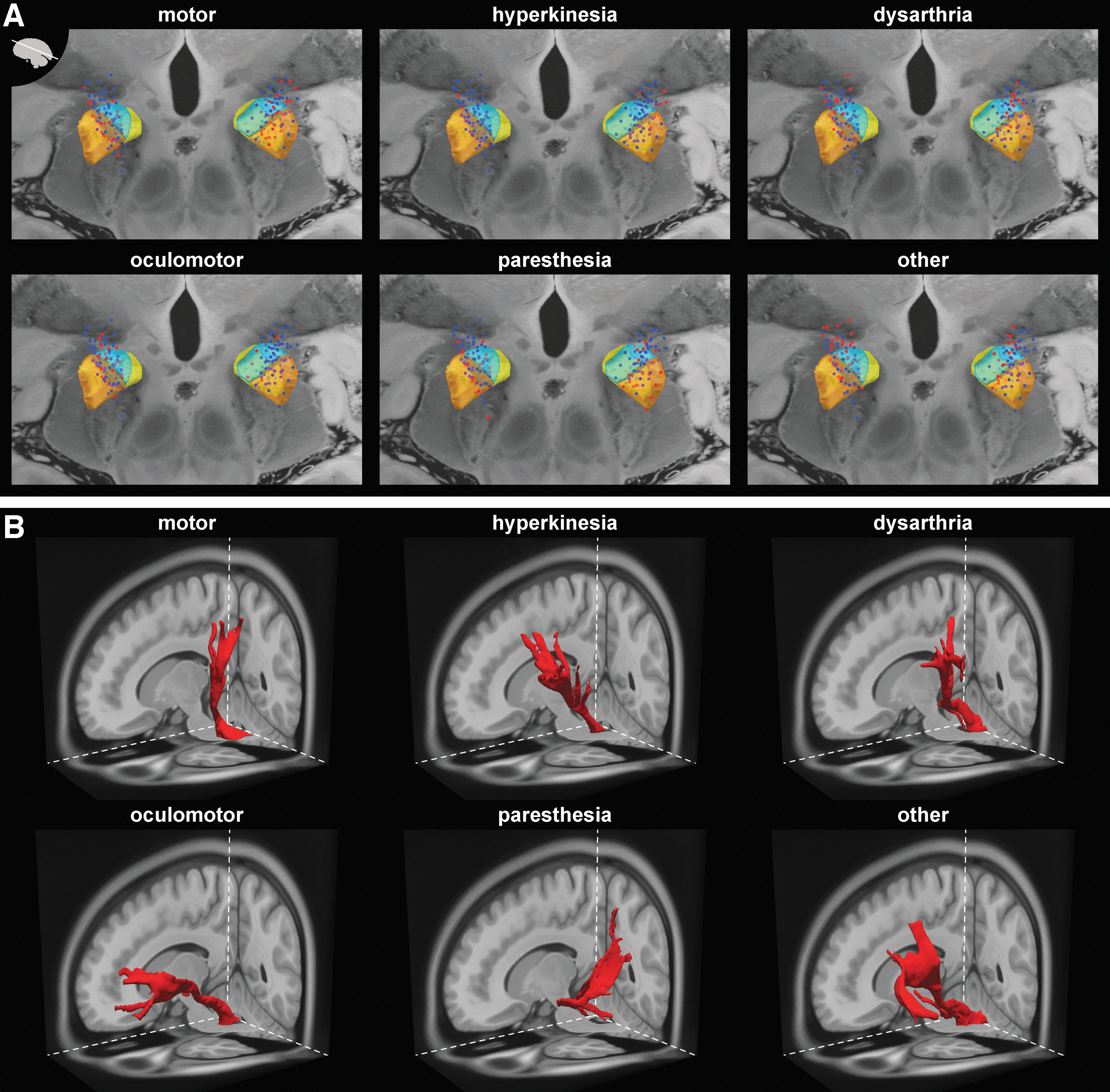
Stimulation sites and representative fiber tracks.
Mid-AC/PC Contact Location
The table lists the coordinates (mean in mm ± SD) of inducing electrode contacts relative to the patient-specific midpoint between anterior and posterior commissure (mid-AC/PC). Axes: x (left/right), y (posterior/anterior), and z (inferior/superior).
Contact connections
As for the spatial overlap in stimulation sites (despite functional differences), we performed a connectivity-based analysis. First, pairs of side effects were compared by their connectivity patterns across all target structures by chi-squared tests for homogeneity. This analysis showed that the connectivity pattern for hyperkinesia and oculomotor effects differed significantly from all other side effects. In addition, the connectivity pattern for motor effects differed from that for dysarthria. Moreover, the connectivity patterns of all side effects differed from the connectivity patterns for symptom reduction (bradykinesia, rigor, tremor). See Table 4 for details. The results of this omnibus comparison suggest that each side effect is associated with one or several distinct connections.
Deviations from Homogeneity of Connectivity Patterns
The table lists the results of chi-squared tests for homogeneity of connectivity patterns (across all target structures) between pairs of side effects. Chi-squared statistics (df = 127 for all comparisons) are shown together with the p-values (in parenthesis). The bottom of the list shows pairwise comparisons between side effects and symptom reduction (adopted from (Strotzer et al., 2019)). A chi-squared value of zero indicates that the frequency distributions (number of connections of inducing or noninducing contacts with target structures) have the same patterns (homogeneity); chi-squared values greater than zero indicate a deviation from homogeneity. Note that the chi-squared statistic is a non-normalized measure of homogeneity (or deviation from it) and a direct comparison of its value is only meaningful for comparisons with the same sample size (n) and the same degrees of freedom (df). n = 168 (except when otherwise stated). *** ≤0.001.
To reveal which of the connections with target structures are relevant, we subsequently examined the contingency between side effect induction and contact connectedness with target structures. This contingency is expressed by phi coefficients (Fig. 2), through which a positive phi means that electrode contacts inducing a side effect are more likely connected with the target structure than noninducing contacts (or vice versa for negative phi). Target structures are ipsilateral to the stimulation site unless otherwise stated. Representative examples of fiber tracks seeded with a single inducing contact are illustrated in Figure 1B.

Connectivity patterns. Contingencies (phi coefficients) between contact induction of side effects and connectedness with target structures for all structures showing a significant association for motor
Motor side effects
Electrode contacts inducing motor side effects did not show significant positive contingencies with target areas. Interestingly, muscle contraction-inducing contacts were not more frequently connected with fibers of the pyramidal tract (internal capsule as reference), primary motor area (M1), or supplementary motor area (SMA) than contacts without motor effects (Fig. 2A). Significant negative contingencies (inducing electrode contacts less frequently connected with the target area) were found for various structures. These included ipsilateral dentate nucleus (DN), ansa lenticularis (AL), retrolenticular/retrolentiform part of the internal capsule (RLIC), lateral part of the internal capsule's anterior limb (ALIC), GPi, internal medullary lamina of the thalamus (IML), and IFG. To control for differences in the acquisition protocols, additional agglutination tests were performed. These tests confirmed the results of the chi-squared test for the whole data set except for the AL target, for which only a marginally significant (p = 0.054) relationship was observed. The mean coordinates of contacts inducing motor effects were significantly more posterior [t(132.1) = 3.30, p = 0.001] and slightly more lateral (not significant) than noninducing contacts (Fig. 1A).
Hyperkinesia
A positive contingency for hyperkinesia existed with the SMA (Fig. 2B). Negative contingencies were found with the ipsilateral superior cerebellar peduncle (SCP), lenticular (f-H2) and thalamic fasciculus (f-H1), AL, and superior parietal lobe (SPL). Inducing contacts were significantly more lateral [t(166) = −3.02, p = 0.003] and superior [t(166) = 3.51, p = 0.001] than noninducing contacts (Fig. 1A).
Dysarthria
Electrode contacts associated with dysarthria were significantly more often connected with the SMA, f-H2, and the decussating dentatorubrothalamic tract (DRTT), but not with M1 (Fig. 2C). Dysarthria inducing contacts were significantly more lateral [t(166) = −4.82, p < 0.001] than noninducing contacts (Fig. 1A).
Oculomotor
Positive contingencies existed between inducing electrode contacts, IFG, and lateral ALIC (Fig. 2D). Negative contingencies were found with M1, SMA, IML, and RLIC. Inducing contacts were significantly more medial [t(166) = 4.85, p < 0.001] and inferior [t(166) = 4.28, p < 0.001] than noninducing contacts (Fig. 1A).
Paresthesia
Inducing contacts had a positive contingency with the RLIC, IML, and the SPL, including the postcentral gyrus (Fig. 2E). Negative associations existed with central pons (cPons) and IFG. Interestingly, inducing contacts were not significantly more often connected to the medial lemniscus than noninducing contacts (82.4% of inducing, 77.8% of noninducing contacts; p = 0.502). Electrode contacts inducing paresthesia were significantly more posterior than noninducing contacts [t(166) = 3.21, p = 0.002] (Fig. 1A).
Other side effects
Due to their manifold presentation and difficult detection, we refrained from analyzing autonomous effects (e.g., sweating) separately and combined them into the group of other side effects. Further symptoms occasionally observed in patients were vertigo, dizziness, dullness, tiredness, restlessness, head/neck pressure, heavy eyes, and limb heaviness. Acute neuropsychiatric symptoms such as anxiety and fear were also included here. Psychiatric side effects (e.g., depression) or other long-term consequences not observable in the acute phase are not included.
Concerning this group, inducing contacts showed significant positive contingencies with contralateral superior (SCP) and middle cerebellar peduncles (MCP), IFG, and superior frontal gyrus (SFG) (Fig. 2F). However, agglutination tests (controlling for acquisition protocol) showed only a marginally significant relationship with SCP (p = 0.065) and MCP (p = 0.065). Inducing contacts were located significantly more medial [t(166) = 3.51, p = 0.001] and anterior [t(166) = −2.53, p = 0.012] than noninducing contacts (Fig. 1A).
Discussion
Our results showed that stimulation of subthalamic electrode contacts resulted in several adverse effects, including motor, oculomotor, paresthesia, dysarthria, hyperkinesia, and other complications. Almost no significant positive contingencies were observed between adverse effects, suggesting contact-specific behavioral effects of stimulation. Despite this specificity, the spatial contact locations largely overlapped. Instead, we found symptom-specific structural connectivity patterns of inducing electrode contacts compared with contacts not associated with these symptoms.
Contact locations
The STN is organized into three functional zones: motor (posterolateral), associative (central), and limbic (anteromedial) (Lambert et al., 2012). These subdivisions are connected with different cortical and subcortical structures (Plantinga et al., 2018). The optimal subthalamic stimulation site for alleviating the main motor symptoms of Parkinson's disease differs from stimulation sites inducing side effects (Akram et al., 2017). Consistently, we observed no contingency between electrode contacts alleviating motor symptoms and those inducing side effects (Table 2), except for contacts inducing dysarthria (see also (Strotzer et al., 2019)).
Most authors proposed lateral STN stimulation as the most effective (best clinical outcome, low stimulation parameters) in alleviating motor disorders (Wodarg et al., 2012). Our results showed that posterior contacts were most likely to induce motor side effects and paresthesia consistent with the notion of the posterolateral STN containing the motor zone (Lambert et al., 2012). Oculomotor effects were caused by medial and inferior contacts, likely adjacent to the oculomotor nerve. Dysarthria-inducing contacts were more lateral, likely overlapping with the motor part of the STN or reflecting the spread of electric current into the internal capsule. Hyperkinesia-inducing contacts were superolateral, likely overlapping with connections to the motor cortex via the internal capsule.
Contact connections
Although side effects tended to depend on the electrode location as expressed by average coordinates, there was a substantial dispersion among inducing contacts. The spatial distribution of inducing contacts overlapped across side effects, and even mean locations were not distinguishable for motor effects and paresthesia. This observation contrasts with the high specificity we observed by our contingency analyses. Group-averaged contact locations as performed regularly (Akram et al., 2017; Horn et al., 2017) are approximate and do not consider interindividual differences in organization and connectivity of the subthalamic area. Our connectivity-based analysis further separated inducing and noninducing contacts based on side effects not distinguishable by contact location.
Motor side effects
This group included phasic movements, facial or limb spasms, paresis, and conjugate eye deviation. These effects are sometimes denoted as capsular side effects because they are likely mediated by corticobulbar and corticospinal fibers (Gorgulho et al., 2009). Other authors (Matias et al., 2015) also included dysarthria and oculomotor effects, which we analyzed separately. We did not find significant positive contingencies between the induction site and connected target areas. Interestingly, no contingencies were observed with targets for SMA and M1, suggesting that stimulation sites related to motor effects are diffusely connected with several cortical motor areas. Moreover, we did not find specific connections with parts of the internal capsule. Together with the observation that contacts are primarily found in the posterior STN region, this suggests that motor side effects are mediated by a complex interaction of structures and not just through individual connections such as the pyramidal tract.
Even though mechanisms differ, and the co-occurrence analysis suggests that it involves different contacts (Table 2), some contacts may elicit both motor and oculomotor side effects (unilateral eye deviation) simultaneously. Reasons may be current spreading beyond both medial and lateral STN borders at higher stimulation amplitudes. This may be exacerbated by differences in local brain tissue (Alonso et al., 2021). Also, stimulation in inferior and anterior STN regions, where the pyramidal tract and the course of the oculomotor nerve are closest in the brainstem, may result in both side effects.
Hyperkinesia
STN-DBS can cause dyskinesia and hyperkinesia (Baizabal-Carvallo and Jankovic, 2016; Zheng et al., 2010), and lesions of the STN may cause symptoms such as hemichorea and hemiballism (Hawley and Weiner, 2012). Our results showed contacts inducing dyskinesia (excessive normal or abnormal movements) were located in the superolateral STN area. These contacts primarily showed connectivity with the SMA (but not M1), a cortical area with diffuse connections (e.g., M1, basal ganglia, corticospinal tract) (Nachev et al., 2008). It has excitatory and inhibitory effects on motor action (Fried et al., 1991). We found significant negative contingencies with the pallido-thalamic fibers (f-H1, f-H2, AL). This complies with pallidal stimulation mitigating dystonia (Kupsch et al., 2006). Some studies (Akram et al., 2017) suggested that connectivity of DBS electrodes with SMA mitigates bradykinesia in Parkinson's patients. Using a different methodology, we could not confirm this notion (Strotzer et al., 2019). Possibly moderate stimulation of fibers connected to the SMA is beneficial for bradykinesia, but overstimulation seems to result in hyperkinesia. Within this notion, DBS-induced dyskinesia reflects fewer side effects but overstimulation of the target area.
Dysarthria
STN-DBS can induce dysarthria, possibly mediated by cerebello-thalamic fibers (Akram et al., 2017; Fenoy and Schiess, 2017; Fytagoridis et al., 2013). Our results showed that contacts inducing dysarthria were associated with the decussating DRTT, a pathway originating in the cerebellum, passing the nucleus ruber, traversing along medial parts of the STN, and terminating in the thalamus (ventral intermediate nucleus) (Kwon et al., 2011). However, contact sites inducing dysarthria in our study were widely distributed and also involved lateral contacts. Moreover, we observed positive contingencies between dysarthria and f-H2, a tract connecting thalamic nuclei with basal ganglia (particularly GPi) and cortical areas (Neudorfer and Maarouf, 2018). This agrees with a recent meta-analysis (Alomar et al., 2017) showing both thalamotomy and thalamic DBS induced dysarthria in about 20% of patients. Furthermore, contacts inducing dysarthria showed a positive connection contingency with the SMA (but not M1). Previous research showed that cortical stimulation of premotor areas was associated with dysarthria/anarthria (Tate et al., 2014). A recent study showed that DBS contacts located close to the pyramidal tract are more likely to induce dysarthria than more distant contacts (Prent et al., 2019). Consistent with this observation, our results suggest that dysarthria induction primarily involves pyramidal tract sections connected with the SMA.
Our analysis did not distinguish between different types of dysarthria. Hence, it is possible that the various networks related to dysarthria, as observed here, reflect various behavioral effects. For instance, cerebellar circuits (DRTT) may be linked to ataxic dysarthria, whereas cortical circuits (SMA) may be linked to spastic dysarthria (Tsuboi et al., 2015). In addition, our clinical testing approach did not contain adequate assessment regarding verbal fluency; an issue that should also be further investigated in future studies.
Oculomotor side effects
These complications are sometimes classified as capsular (conjugate eye deviation) or noncapsular (unilateral eye deviation) (Matias et al., 2015). Shields and colleagues (2007) suggested that contacts inducing conjugate eye deviation stimulate frontal eye field axons with trajectories in the medial part of the ALIC. We found a significantly positive contingency with the lateral ALIC and less with its medial aspect. The frontal eye field is usually located in the middle frontal gyrus and its vicinity (Vernet et al., 2014). Consistently, we found a positive contingency with the IFG. The inferior/ventral frontal cortex has been associated with inhibitory oculomotor control (Hodgson et al., 2007). Oculomotor effects in our study were primarily characterized by unilateral eye deviations. Hence, these effects may be mediated by the oculomotor nerve. Contacts inducing these side effects were located in the medial and inferior parts of the STN-region adjacent to the ipsilateral oculomotor nucleus and nerve (Park et al., 2017). Unfortunately, our tractography approach did not allow us to identify this nerve.
Paresthesia
Consistent with previous studies (Fytagoridis et al., 2013), contacts of the posterior STN were associated with paresthesia. Our connectivity-based results suggest this occurs from current spreading beyond the posterior edge of the STN, affecting spinothalamic fibers and parts of the RLIC, where optic radiation fibers travel next to the sensory fibers of the internal capsule's posterior limb. Furthermore, paresthesia may be caused by thalamo-cortical projections to the parietal lobe (Gibson et al., 2016). Accordingly, we found inducing contacts were connected with the IML and the SPL (including the postcentral gyrus). Our data only showed a weak contingency for connections with the medial lemniscus, suggesting it plays a minor role in evoking paresthesia.
Other side effects
Not only behavioral effects but also the spatial location (anterior and medial) and the connectivity pattern of inducing contacts were diverse in this group. Positive contingencies were found with the frontal cortex and contralateral cerebellar peduncles (findings for DN approach significance), suggesting at least some of these side effects were mediated by diffuse connections to the frontal and cerebellar structures (beyond the cerebello-thalamo-cortical loop). Previous research showed that unilateral stroke in the MCP might produce vertigo and imbalance (Kim and Kim, 2019). Moreover, fatigue seems to involve cerebellar structures (vermis crus I) (Lazzarotto et al., 2020) and the lateral frontal cortex (Goñi et al., 2018).
Negative contingencies
We also observed negative contingencies (negative phi) for most effects, meaning contacts where stimulation induced specific behavior were significantly less often connected with target structures. This was predominantly found for motor, hyperkinesia, and oculomotor side effects. These electrode contacts may be considered “safe spaces” for avoiding side effects.
Methodological considerations
Advantages
Fiber tracking and track-to-target evaluation were performed in the patients' individual image space (rather than group-averaged space), which accounted for interindividual anatomical differences (Wang et al., 2020). We investigated a relatively homogenous patient collective and a vast number of target areas. Probabilistic tractography is more sensitive and specific than deterministic approaches (Schlaier et al., 2017).
Limitations
Our results are based on single-center data. We only investigated structural connectivity by tractography based on diffusion MRI. Combining structural and functional connectivity may be beneficial (Horn et al., 2017). Our seed regions were not adjusted by stimulation parameters (Gunalan et al., 2018) but kept of constant size. Our data were collected by two different clinical scanners (1.5T, 3T) available at the time of data collection. Fractional anisotropy may increase with field strength (Chung et al., 2013). However, in our study, differences in connectivity between inducing and noninducing contacts showed a similar pattern regardless of the acquisition protocol or field strength. Only immediate stimulation effects during the first programming session were investigated. Delayed effects such as the influence on freezing were not captured by our standardized test battery (Barbe et al., 2020; Fasano et al., 2011; Georgiades et al., 2019). Our classification of side effects was motivated by clinical demands, and other authors may adopt a different classification. Some side effects were assessed by self-report of the patient only and could not be objectively measured.
Conclusions
Adverse effects of STN-DBS are associated with specific electrode contacts and usually involve other contacts than those effective in mitigating Parkinson's symptoms. Although these electrode contacts partially overlap in volumetric space, they may be separated by their structural connectivity patterns. Contacts inducing side effects are connected with the SMA, DRTT, IML, f-H1, f-H2, SPL, frontal gyrus, cerebellar structures, and internal capsule fibers. Although relevant for mitigating motor symptoms in Parkinson's disease, cerebellar circuits seem to be less involved in the occurrence of side effects. Target planning in DBS may substantially benefit from connectivity patterns of electrode contacts in the individual brain. Connectivity-based targeting may not only maximize treatment effectiveness (Accolla et al., 2016; Akram et al., 2017; Horn et al., 2017; Strotzer et al., 2019; Vanegas-Arroyave et al., 2016), but it may also minimize DBS-induced side effects.
Data Access and Responsibility
Q.D.S., J.R.S., and A.L.B. take responsibility for the integrity of the data and the accuracy of the data analysis. All data used for this study are preserved at the Neurosurgical Institute of the University Hospital Regensburg. Data are accessible on-demand as far as allowed by the guidelines of the Ethics Committee of the University Hospital Regensburg. Requests should be addressed to the corresponding author.
Footnotes
Acknowledgments
We thank F. Zeman (Center for Clinical Trials, University Hospital Regensburg, Germany) for help with the statistical analysis and M. Lange for his valuable advice.
Authors' Contributions
Q.D.S.—Research project: conception, organization, and execution; statistical analysis: design and execution; article preparation: writing of the first draft, figure preparation, and final approval. Z.K.—Research project: execution and data acquisition; statistical analysis: review and critique; article preparation: review and critique, and final approval. J.M.A., R.F., and C.F.—Research project: conception, organization, execution, and data acquisition; article preparation: review and critique, and final approval. N.O.S.—Statistical analysis: review and critique; article preparation: review and critique, and final approval. E.T.—Research project: execution and data acquisition; article preparation: review and critique, and final approval. M.W.G.—Statistical analysis: review and critique; article preparation: review and critique, and final approval. J.R.S.—Research project: conception, organization, execution, and data acquisition; statistical analysis: review and critique; article preparation: review and critique, and final approval. A.L.B.—Research project: conception, organization, and execution; statistical analysis: design and execution; article preparation: review and critique, figure preparation, and final approval.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Q.D.S. has received a study scholarship from the German Academic Scholarship Foundation (Studienstiftung des deutschen Volkes). J.R.S. has received personal compensation for teaching, sponsored scientific talks, and consultation from Medtronic and Abbott (St. Jude Medical) outside of the submitted work. Z.K. has received personal compensation for lecturing from Desitin GmbH, AbbVie, and UCB Pharma outside of the submitted work. M.W.G. and A.L.B. were partially supported by the Deutsche Forschungsgemeinschaft (DFG, Project GR988-25/1). All other authors have no funding information to report. Sponsors were not involved in the study design, collection, analysis, and interpretation of data, the writing of the report, and the decision to submit the article for publication.