
Abstract
Introduction:
The thalamus, a heterogeneous brain structure, is involved in the generation of sleep-related thalamo-cortical oscillations. Higher order nuclei might possess a distinct function compared with first-order nuclei in brain communication. Here it is investigated whether this distinction can also be found during the process of falling asleep and deepening of slow-wave sleep.
Methods:
A nonlinear version of Granger causality was used to describe changes in directed network activity between the somatosensory cortex and rostral reticular thalamic nucleus (rRTN) and caudal reticular thalamic nucleus (cRTN), the higher order posterior (PO)- and anterior-thalamic nuclei (ATN), and the first-order ventral posteromedial thalamic nucleus (VPM) as assessed in local field potential recordings acquired during passive wakefulness (PW), light slow-wave sleep (LSWS), and deep slow-wave sleep (DSWS) in freely behaving rats. Surrogate statistics was used to assess significance.
Results:
Decreases in cortico-thalamo-cortical couplings were found. In contrast, multiple increases in intrathalamic couplings were observed. In particular, the rRTN increased its inhibition on the ATN from PW to LSWS, and this was further strengthened from LSWS to DSWS. The cRTN increased its coupling to VPM and PO from PW to LSWS, but the coupling from cRTN to VPM weakened at the transition from LSWS to DSWS, while its coupling to PO strengthened. Furthermore, intra-RTN coupling from PW to LSWS was differently changed compared with the change from LSWS to DSWS.
Discussion:
It can be inferred that higher order (ATN and PO) and first-order nuclei (VPM) are differentially inhibited during DSWS, which might be relevant for a proper functioning of sleep-related processes.
Impact statement
The functionally heterogeneous thalamus is affected by the different sleep/wake states. Changes in directed functional coupling between the thalamus and cortex and between functional different thalamic nuclei during the process of falling asleep and deepening to slow-wave sleep were investigated. It was revealed that the rostral and caudal subparts of the reticular thalamic nucleus, constituting the major source of intrathalamic inhibition, decouple from each other and show different coupling profiles with other thalamic nuclei. Specifically, higher order nuclei were found to be more inhibited than first-order nuclei during deep slow-wave sleep. These differences might be relevant for a proper coordination of sleep-related processes such as housekeeping, forgetting of irrelevant information, and consolidation of episodic memory.
Introduction
The thalamo-cortical system is a strongly interconnected brain system involved in a multitude of physiological processes. Different nuclei of the thalamus are involved in the relay or processing of sensory information, the direction of attention, or are involved in cognitive functions such as decision-making or memory processing (Jones, 2012). In general, thalamic nuclei are subdivided into two major classes. First-order thalamic nuclei receive their driving input from the periphery and relay information to the cortex. In contrast, higher order thalamic nuclei receive their driving input from the cortex and are proposed to be involved in corticocortical communication (Sherman, 2016). Neurons projecting to or from either first-order or higher order thalamic nuclei possess different firing properties. For example, higher order nuclei are more prone to burst firing compared with first-order nuclei (Ramcharan et al., 2005). The reticular thalamic nucleus (RTN), which constitutes the major source of intrathalamic inhibition in the rodent brain, directs parvalbumin-positive gamma-aminobutyric acid (GABAergic) neurons to first-order nuclei, and calbindin-positive GABAergic neurons to higher order thalamic nuclei. In turn, both cell types were reported to differ in their firing properties (Martinez-Garcia et al., 2020). As a consequence, it can be hypothesized that higher order and first-order nuclei support the abovementioned physiological and cognitive functions in different ways.
The cortico-thalamo-cortical system is strongly affected by different sleep/wake states. Thalamic degeneration, with selective or prevalent involvement of the anterior or dorsomedial nuclei, virtually abolishes the ability to generate electroencephalographic (EEG) sleep patterns (Lugaresi, 1992). Moreover, it is known that the RTN hyperpolarizes thalamic relay neurons during sleep to reduce the transfer of incoming sensory stimuli during slow-wave sleep (Coenen and Vendrik, 1972). Network analysis in wild-type (WT) mice with an adaptive nonlinear Granger causality (GC) method revealed a bidirectional reduction of coupling strength between the ventral posteromedial thalamic nucleus (VPM), a first-order thalamic nucleus, and the somatosensory cortex during slow-wave sleep compared with wakefulness (Zobeiri et al., 2019). However, it is less clear if this gating of information is gradually increasing during the process of falling asleep and deepening of slow-wave sleep, if first- and higher order thalamic nuclei are equally affected by inhibition of the RTN and what type of changes are occurring within the thalamus between the functionally distinct thalamic nuclei.
The cortico-thalamo-cortical system is also involved in the pathophysiological condition of absence epilepsy, characterized by short lasting (5–20 sec) spontaneous hypersynchronous activity expressed as spike and wave discharges (SWDs) in the EEG of genetic epileptic rats (Inoue et al., 1993). In an attempt to investigate network mechanisms that are responsible for the generation of SWDs, multisite local field potential (LFP) recordings were simultaneously recorded in layers 4, 5, and 6 of the somatosensory cortex (S1), the VPM (first-order nucleus), the posterior thalamic nucleus (PO; higher order nucleus), the caudal RTN (cRTN), which sends GABAergic projections to PO and VPM and received collaterals of cortico-thalamic neurons, the rostral RTN (rRTN), which projects to the limbic thalamus, and the anterior thalamus nucleus (ATN; another higher order thalamic nucleus), which receives GABAergic projections of the rRTN (Crabtree, 2018; Gonzalo-Ruiz and Lieberman, 1995; Pinault and Deschênes, 1998). This was done in a Wistar derived genetic generalized absence epilepsy model, rats of the Wistar Albino Glaxo rats from Rijswijk (WAG/Rij) strain. Rats of this strain show normal amounts of slow wave sleep and a normal circadian distribution of the various sleep/wake stages (Smyk and van Luijtelaar, 2020). Moreover, SWDs typical for this strain are not preceded or followed by deep slow-wave sleep (DSWS) (Smyk et al., 2019) and SWD-related changes in network communication are restricted to roughly 2 sec before SWD on- and offset (Lüttjohann et al., 2014; Lüttjohann and van Luijtelaar, 2012). Therefore, this data set is suited to provide insight into the abovementioned questions concerning cortico-thalamo-cortical and intrathalamic network changes occurring during the process of falling asleep and deepening of slow-wave sleep. In the current study, these were investigated with a directed nonlinear version of GC (Sysoev and Sysoeva, 2015), which was used to determine the strength and direction of coupling between brain structures during periods of passive wakefulness (PW), light slow-wave sleep (LSWS), and DSWS.
Methods
Animals, experimental procedure
LFPs were recorded in layers 4, 5, and 6 of the somatosensory cortex, ATN, rRTN, cRTN, PO, and VPM in 16 freely moving adult WAG/Rij rats. In brief, the LFP signals, amplified, filtered (1 [high pass]–100 Hz [low pass], 50 Hz [notch-filter]), and digitalized at 2048 Hz, were acquired for 4 h during the dark phase of the 12–12 light-dark-cycle. Movements of rats were registered by the Passive-Infrared-Registration system (PIR, RK2000DPC LuNAR PR Ceiling Mount, Rokonet). For detailed information concerning surgical procedures and histological verification of electrode positions, the reader is referred to Lüttjohann et al. (2013) as well as Supplementary Data. The Ethics Committee on Animal Experimentation of Radboud University, Nijmegen, approved all the experimental procedures.
Coupling analysis
Ten representative 10-sec lasting episodes of PW, LSWS, and DSWS of each animal were selected by a trained electrophysiologist based on the following criteria: PW was characterized by a motionless animal as indicated by a flat PIR with only minor movements and a low-amplitude, high-frequency (∼8–20 Hz) cortical EEG. DSWS was characterized by a flat PIR and a high-amplitude, slow wave (1–5 Hz) cortical EEG, LSWS by a motionless animal (flat PIR), and an EEG of intermediate amplitude and intermediate frequency (3–6 Hz; Fig. 1). Each period had a minimal distance of 10 sec to epileptic SWD, to ensure analysis of signals that are free of network changes, related to either the generation or termination of SWDs.
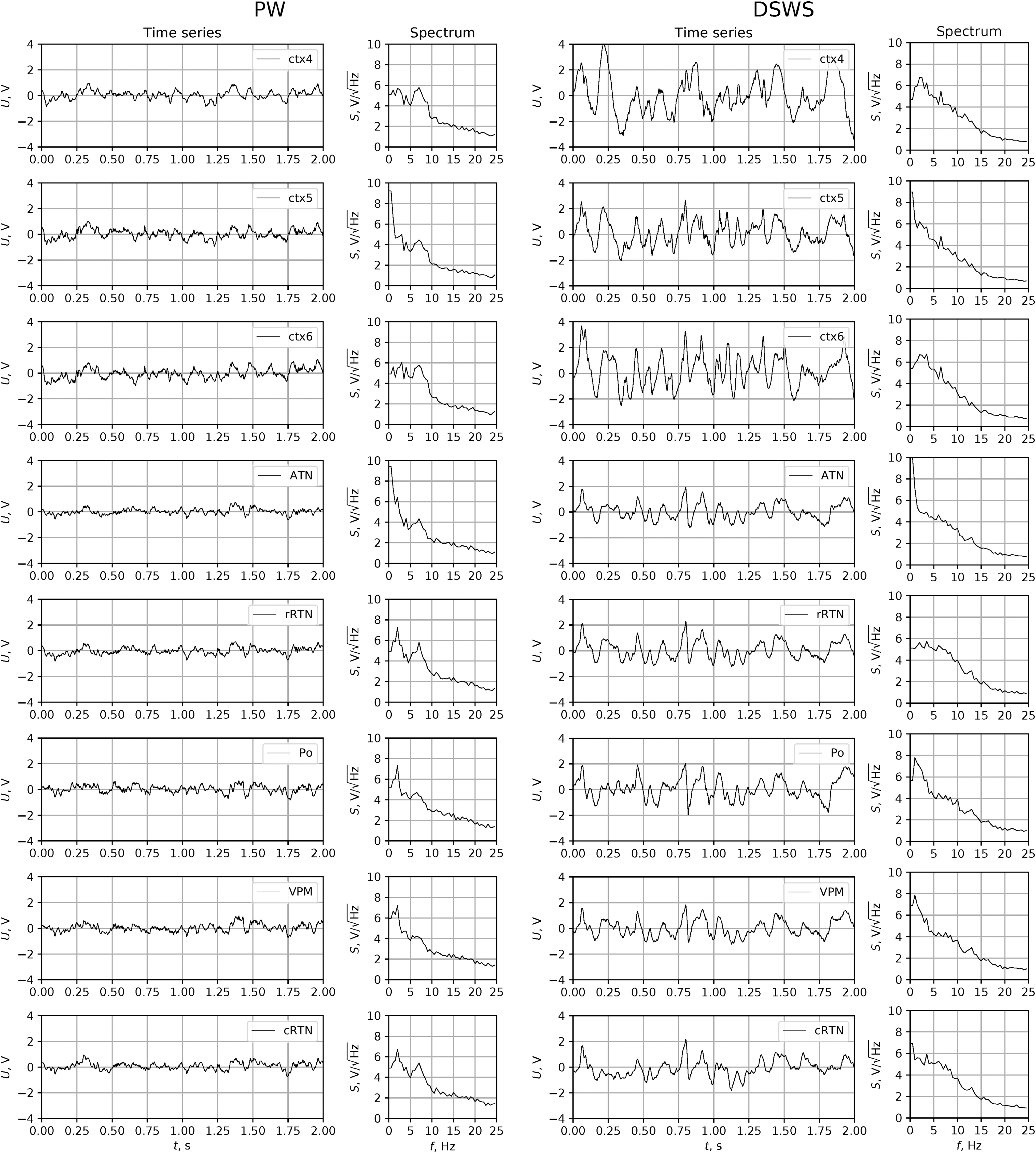
Exemplary LFP traces recorded in layers 4, 5, and 6 of the somatosensory cortex and different thalamic nuclei during PW (left panel) or DSWS (right panel) and corresponding power spectra. ATN, anterior thalamus nucleus; cRTN, caudal reticular thalamic nucleus; ctx4, layer 4 of somatosensory cortex, ctx5, layer 5 of somatosensory cortex, ctx6, layer 6 of somatosensory cortex; DSWS, deep slow-wave sleep; LFP, local field potential; PO, posterior thalamic nucleus; PW, passive wakefulness; rRTN, rostral reticular thalamic nucleus; VPM, ventral posteromedial thalamic nucleus.
Coupling between brain structures was analyzed for the three vigilance states using the nonlinear adapted GC method (Kornilov et al., 2016; Sysoev and Sysoeva, 2015). Differences between PW and LSWS as well as between LSWS and DSWS were determined and subject to statistical analysis. The GC method is a popular approach to coupling analysis in neuroscience and has been improved significantly throughout the years (Chen et al., 2004; Kornilov et al., 2016; Marinazzo et al., 2006; Marinazzo et al., 2008). Here, the specific model developed by Sysoeva and Sysoev (2012) is used. This model is constructed according to a nonlinear autoregressive model Eq. (1).
where f is a polynomial of general type of order P from Ds
variables—components of the state vector
Polynomial f has
Two models, a univariate (self) Eq. (1), and bivariate (joint) Eq. (2) are necessary to compute the prediction improvement (PI), a commonly used characteristic of GC.
where Da
is an additional dimension—dimension of vector
Parameters l,
When Eqs. (1) and (2) were fitted to data using a least-squares routine (Virtanen et al., 2020), the PI was calculated following Eq. (3). For this, the mean least-square errors of Eqs. (1) and (2) denoted further as
Since some method parameters (Ds and P) were chosen separately and differently for the three considered states, the absolute PI values by themselves cannot be straightforwardly compared between different states. Instead, an approach based on the number of significant coupling detections in the ensemble of episodes was used (see the Statistics section; Grishchenko et al., 2020).
Statistics
Surrogate tests for series permutation surrogates were used to identify differences in coupling between the three states (Sysoev and Sysoeva, 2015). LFP data of 10 real episodes of the same state (PW, LSWS, and DSWS) from a particular channel-pair of each animal were shuffled randomly to obtain 100 different combinations. From these combinations, only 10 corresponded to real data and 90 were surrogates. The GC method was applied both to the real and surrogate data. The highest surrogate value calculated for surrogates
The total number of significant PIs was divided by the total number of considered episodes n, for all included animals, providing the average over animals' percentage of significant couplings (averaged coupling strength). To compare the number of significant coupling detections for different states, a correction for multiple comparison was used
where
Equation (5) was used to determine the significance of coupling in the selected state rather than the difference between two states; it is similarly obtained as in Eq. (4).
This approach was justified and described in detail by Grishchenko et al. (2020).
The traditional value of
Results
Significant couplings between the cortex (ctx) and thalamus and vice versa and within the thalamus were present during all three brain states, and coupling strength as well as the dominant directions for most of them changed between states (Figs. 2 and 3; Table 2). We report all couplings during PW and all changes in coupling during the process of falling asleep and deepening of sleep, which were significant (p < 0.011) as assessed by surrogate statistics (see the Methods section).
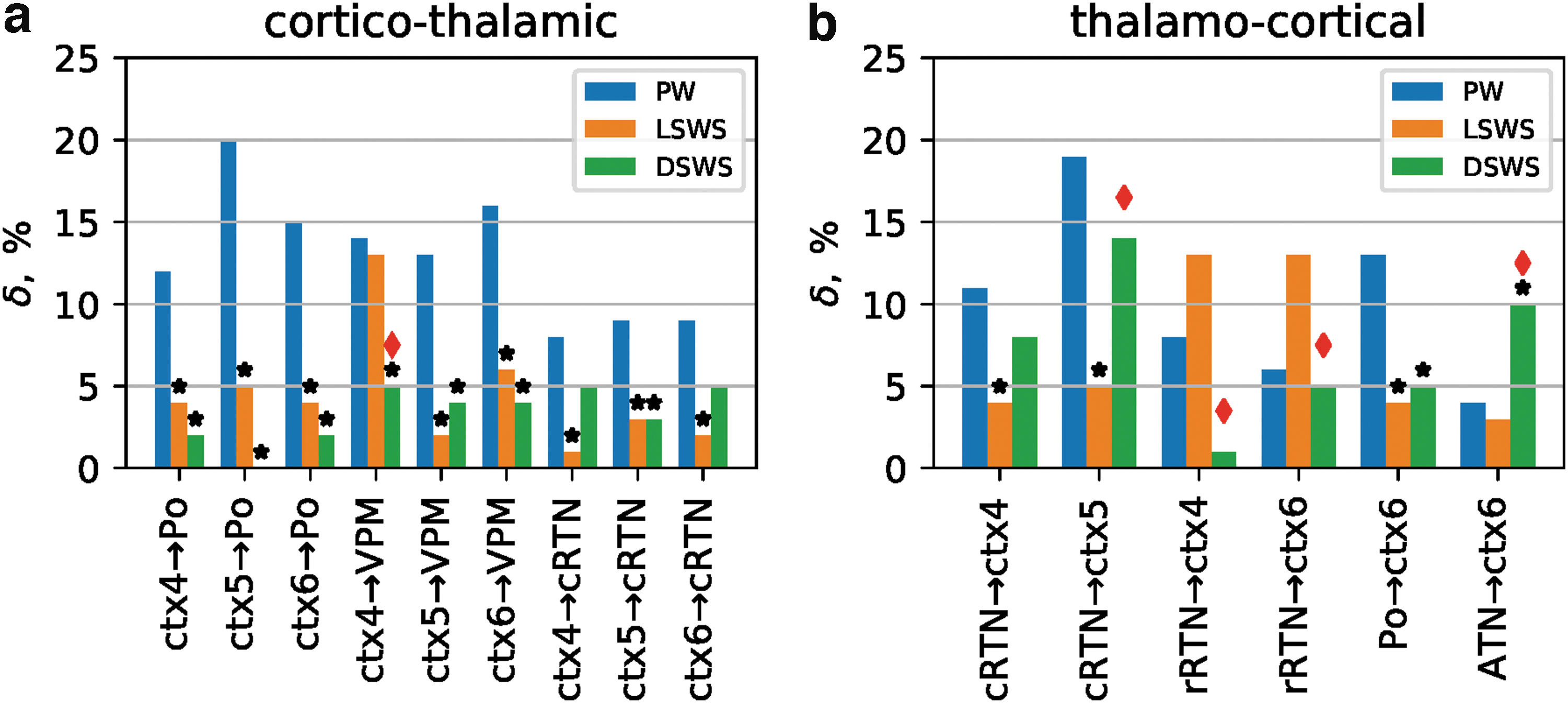
Cortico-thalamic
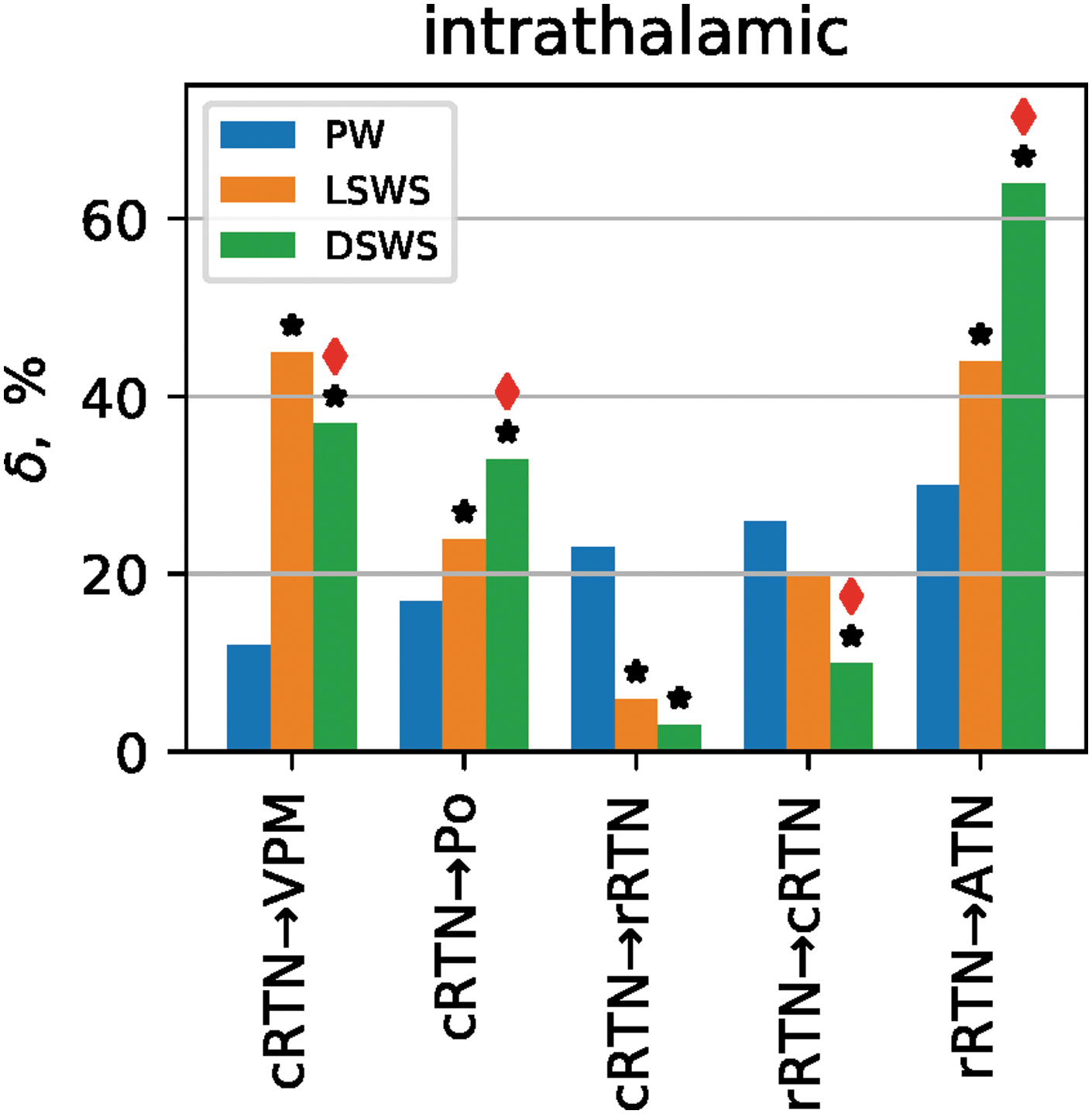
Intrathalamic coupling during PW, LSWS, and DSWS. Bar plot of coupling strength for all intrathalamic pairs with a direct anatomical connection, in which at least one transition from PW to LSWS or from PW to DSWS or from LSWS to DSWS was found to be significant, is shown. Bars demonstrate the coupling strength in percent (
Intrathalamic Coupling During Passive Wakefulness, Light Slow-Wave Sleep, and Deep Slow-Wave Sleep
All marked results are significant at a p < 0.01 level as verified using surrogate statistics.
Indicates a significant coupling during PW.
Indicates a significant change in coupling strength compared with PW.
Indicates a significant change in coupling strength compared with LSWS.
Cortico-thalamo-cortical couplings
Couplings from ctx to thalamus during PW were characterized by significant couplings (
Cortico-Thalamo-Cortical Coupling During Passive Wakefulness, Light Slow-Wave Sleep, and Deep Slow-Wave Sleep
All marked results are significant at a p < 0.01 level as verified using surrogate statistics.
Indicates a significant coupling during PW.
Indicates a significant change in coupling strength compared with PW.
Indicates a significant change in coupling strength compared with LSWS.
ATN, anterior thalamus nucleus; cRTN, caudal reticular thalamic nucleus; ctx, cortex; DSWS, deep slow-wave sleep; LSWS, light slow-wave sleep; PO, posterior thalamic nucleus; PW, passive wakefulness; rRTN, rostral reticular thalamic nucleus; VPM, ventral posteromedial thalamic nucleus.
Significant changes in cortico-thalamo-cortical coupling as a consequence of falling asleep or deepening of sleep are depicted in Figure 2.
As can be seen for most of the abovementioned cortico-thalamic couplings (i.e., from ctx 5 and ctx6→VPM [both p < 0.001]; ctx4, ctx5, and ctx6→PO [all p < 0.002]; and ctx4, ctx5, and ctx6→cRTN [all p < 0.01]), a significantly reduced coupling both during LSWS and DSWS was found compared with PW. Of note, the decrease in coupling for all these pairs already occurred at the transition from PW to LSWS, while no further significant additional reductions were found during deepening of sleep (i.e., from LSWS to DSWS; Fig. 2a; Table 1). Significant decreases in coupling from PW to LSWS in the reverse direction were found between PO→ctx6 (p < 0.001), cRTN→ctx4 (p < 0.001), and cRTN→ctx5 (p < 0.001). Again, no further decrease was found between LSWS and DSWS (Fig. 2b; Table 1). Of note, the cRTN only possesses a bisynaptic connection to the somatosensory cortex (i.e., it projects to PO and VPM, which in turn project to S1).
Compared with a relatively high number (11 out of 30) of decreases in couplings between cortex and thalamus observed from PW to LSWS, only a few coupling (3 out of 30 and only unidirectional) diminishments were observed between LSWS and DSWS. One from ctx4→VPM, and two from rRTN to ctx4 and ctx5. Of note, the rRTN does not possess a direct connection to the cortex (i.e., rRTN projects to ATN, which in turn projects to frontal cortices but not to S1).
Lastly, in contrast to the overall decrease in cortico-thalamo-cortical couplings in the process of falling asleep and deepening of sleep, only a single increase in coupling was seen between PW and DSWS for the ATN→ctx6 (p < 0.01) pair. However, it needs to be mentioned again that there is no direct anatomical connection between these two structures (i.e., ATN projects to frontal cortices but not to S1). In general, it can be stated that the changes are more pronounced in the direction from cortex to thalamus than from thalamus to cortex.
Intrathalamic couplings
Many significant couplings between the various thalamic nuclei were found during PW (Fig. 3; Table 2). The VPM, ATN, and PO do not possess direct anatomical connection with each other, but all are indirectly coupled via the RTN (i.e., ATN is bidirectionally connected to the rRTN, PO and VPM are connected to the cRTN, and both RTN subparts are coupled to each other).
Most pronounced and significant (+30%, p < 0.001) were the direct unidirectional coupling rRTN→ATN, and the direct bidirectional coupling between the two parts of the RTN (rRTN→cRTN, +26%, p < 0.01); cRTN→rRTN (+23%, p < 0.01). Most other thalamic nuclei were also bidirectional and significantly coupled with each other, although less strong (Fig. 3; Table 2).
The RTN parts modulate not only each other, but also the three other thalamic nuclei. The modulation of the cRTN→VPM was expected, but also the limbic ATN is strongly modulated by both the rRTN (+30%, p < 0.001) and cRTN (+20%, p < 0.001). The higher order PO nucleus was also bidirectionally modulated by both the cRTN and rRTN (rRTN→PO; +8%, p < 0.01) and PO→rRTN (+11%, p < 0.01); cRTN→PO (17%, p < 0.001); and PO→cRTN (12%, p < 0.01), all significant. This implies a function of the RTN, which goes beyond the modulation of only sensory information in the VPM.
Sleep-induced changes in intrathalamic couplings, which also do possess a direct anatomical connection, are presented in Figure 3.
It can be seen that, both during LSWS and DSWS, the inhibitory coupling of both subparts of the RTN to their primary projection nuclei was significantly increased compared with PW. At the same time, coupling between the two RTN subparts was significantly decreased compared with PW (Fig. 3).
Interestingly, the coupling changes between the two poles of the RTN were rather different compared with the changes between RTN and most other thalamic nuclei. They showed during the shift from PW to LSWS, a unidirectional decrease (−16%, p < 0.001) in coupling strength from cRTN→rRTN, and a unidirectional decrease (−10%, p < 0.01) in coupling in the reversed direction at the transition from LSWS to DSWS (Fig. 3). The couplings within the RTN sketch a picture of reciprocal couplings during PW, a unidirectional decrease from caudal to rostral during LSWS, while a unidirectional decrease from rostral to caudal during DSWS is noticed, indicating that the direction and amount of changes of within RTN effective coupling can be different and depend on the sleep state.
While coupling from cRTN→PO and rRTN→ATN demonstrated a gradual increase in coupling (i.e., the cRTN and rRTN increased its directional coupling on their higher order thalamic nuclei PO (+7%, p < 0.01) and ATN (+14%, p < 0.01) at the transition from PW to LSWS and further strengthened their coupling from LSWS to DSWS (both p < 0.01), coupling from cRTN to first-order nucleus VPM showed an increase from PW to LSWS (+32%, p < 0.001), but thereafter no further increase during the state change from LSWS to DSWS in the direction cRTN→VPM, instead, an effect in the opposite direction, a significant decrease (−8%, p < 0.01), was detected (Fig. 3).
Lastly, next to the changes observed for nuclei with a direct anatomical connection, significant changes were also noted for several indirectly connected nuclei (Table 2). The general picture is again that intrathalamic couplings increased during sleep compared with PW. Most pronounced and consistent changes were observed for the indirect connection between rRTN→PO (+35% and +8 from PW to LSWS and LSWS to DSWS, respectively (both p < 0.01), and the direct connection rRTN→ATN (+14%, +20%, both p < 0.01).
Discussion
An adapted version of GC was used to describe changes in directed coupling between the somatosensory-cortex and various thalamic nuclei and between thalamic nuclei during the process of falling asleep and deepening to slow-wave sleep. The overall effects revealed that (1) the directed connectivity between the cortex and thalamus and to a minor extent vice versa was reduced during sleep, while intrathalamic couplings were largely increased; (2) changes in cortico-thalamic coupling are not gradual as sleep proceeds from PW to DSWS but mostly occur at the transition from PW to LSWS; (3) rostral and caudal subparts of the RTN decoupled from each other and showed different coupling profiles with the other thalamic nuclei; (4) while the rRTN increased its inhibition to higher order ATN and PO and first-order (VPM) from PW to LSWS and also strengthened (increased) this inhibition to the higher order nuclei from LSWS to DSWS, the cRTN then weakened its inhibition to the first-order VPM from LSWS to DSWS, demonstrating a differential inhibition of first- versus higher order nuclei during DSWS.
Cortico-thalamo-cortical couplings
The stronger couplings from the cortex to thalamus compared with the thalamus to cortex, as seen during PW, are in line with studies, describing many more projections from the cortex to thalamus compared with the reversed direction (Deschênes et al., 1998). Decreased coupling during sleep between the cortex and the central-medial-nucleus was also found in humans in combined EEG/functional magnetic resonance imaging (fMRI) studies (Picchioni et al., 2014; Spoormaker et al., 2010), demonstrating that the thalamo-cortical connectivity was sharply reduced at the transition from wakefulness to light sleep.
Our study also revealed that cortico-thalamic coupling decreases were most pronounced for the PW to LSWS transition. The reduced coupling was then maintained during DSWS but did not decrease any further when sleep deepened, demonstrating that the reduction in cortico-thalamic communication as a function of falling asleep and deepening of sleep is not a gradual process. It might be speculated that a minimally required communication is already reached during LSWS and/or that a certain amount of cortico-thalamic communication remains to be maintained for processes proposed to take place during DSWS, for example, for housekeeping or restorative processes (Landolt and Dijk, 2019), or the consolidation of memory (Born and Wilhelm, 2012; Dalrymple-Alford et al., 2015; Diekelmann and Born, 2010). Based on the overall reduction in cortico-thalamo-cortical coupling, it is proposed that a functional consequence of slow-wave sleep (SWS) is a diminishment of cortico-thalamic communication.
Electrocorticography studies, simultaneously targeting the thalamus and cortex, are extremely rare. In our hands, the general decrease in coupling between the cortex and thalamus when sleep occurred or deepened was present in both directions, but more pronounced from cortex to thalamus. This coupling from cortex to thalamus is much less investigated than in the reversed direction. In the latter line, there are different observations, including the early neurophysiological observations in sleeping cats, showing that the ratio between outgoing and incoming action potentials of the lateral geniculate is reduced during sleep and anesthesia compared with wakefulness (Coenen and Vendrik, 1972), and that during sleep thalamo-cortical (TC) cells fire in a bursting mode due to increased GABAergic inhibition induced by the RTN. Due to that, a basic property of hyperpolarized thalamic cells devoid from depolarizing afferents, gating of information, is enhanced. Larson-Prior et al. (2009) proposed that the transitions from wakefulness to successively deeper stages of slow-wave sleep progressively disengage the self from the environment; it is rather likely that the reduction in thalamic sensory transfer contributes to this. Other evidence for reduced sensory information transfer from thalamus to cortex during sleep comes from auditory and visual-evoked potential studies, in which characteristics of auditory evoked potentials and visual evoked potentials during sleep were slower and with some modified components during slow-wave sleep compared with wakefulness in humans and in rats (Campbell, 2000; Meeren et al., 2001), and from combined fMRI-EEG studies (Portas et al., 2000). Direct evidence for diminished bidirectional couplings between the somatosensory cortex and lateral basal complex of the thalamus during natural occurring DSWS compared with wakefulness was recently reported in free moving WT mice (Zobeiri et al., 2019). We found that only some thalamic nuclei were selectively less effected during sleep by the somatosensory cortex: PO, VPM, and cRTN were no longer driven, the more limbic rRTN and ATN kept their drive from the cortex. As both these structures belong to the limbic thalamus, which is involved in emotional and memory processes, it can be speculated that a certain degree of cortico-thalamic communication is needed to be maintained during sleep for proper functioning of the limbic system.
Intrathalamic couplings: the distinction between higher order and first-order thalamic nuclei and rRTN and cRTN
The projections within the RTN and from RTN to VPM, PO, and ATN are all GABAergic (Gonzalo-Ruiz and Lieberman, 1995; Martinez-Garcia et al., 2020), the projections from thalamus to the RTN are glutamatergic, as well as the TC and cortico-thalamic (CT) cells to and from the cortex, respectively. It is also commonly accepted that at the transition from PW to DSWS, the classical sensory relay cells and also other thalamo-cortical projecting neurons become more hyperpolarized due to an increase in GABAergic influences from the RTN (Sherman and Guillery, 2006). The RTN is in rats the only source of inhibitory influences on TC cells, with an exception for the lateral geniculate. This implies that if there is more GABAergic inhibition during slow-wave sleep than during PW, than the increases in coupling strength from the RTN to the other thalamic nuclei, as indicated in Figure 3, demonstrated an increase in inhibitory effects. The RTN increases its inhibitory effect from PW to LSWS to PO, and ATN as well as VPM. It is striking that the RTN only further increases its inhibitory coupling to the higher order nuclei ATN and PO when sleep deepens; while this is not the case for the cRTN- to the first-order thalamic nucleus VPM. This puts the VPM in a different position as the two higher order nuclei. The VPM is a classical relay station for environmental somatosensory information passing to the cortex. The inhibitory RTN is often regarded as a gatekeeper to the cortex, and in this way, the cortex receives less sensory inputs during light sleep, most likely to protect the brain from awakening. The lack of strengthening of inhibitory influence from the cRTN to VPM in the transition from LSWS to DSWS might ensure the possibility for an urgent wake-up call in case of the presence of important environmental stimuli. In fact this has been already proposed by Sherman (2001).
ATN and PO, on the contrary, are higher order thalamic nuclei, which receive their “driving input” from the cortex and are thought to be involved in cortico-cortical communication. PO neurons are driven by layer 5 input from a cortical area, and these signals are transferred via the PO to other cortical areas (Sherman and Guillery, 2006). The RTN showed a large increase in coupling to the PO from PW to LSWS and an additional smaller enhancement during DSWS, while the cortex reduced its excitatory drive during LSWS. The transfer of information function seems inhibited during sleep and together with the increased GABAergic inhibition, it is suggested that the PO becomes more isolated from the cortex, partly preventing cortico-cortical information transfer via thalamus during sleep. In line with this, decreases in intracortical coupling (cortical effective connectivity) during sleep were reported in EEG of sleeping humans (Massimini et al., 2005). Given its isolated position during sleep, including the reduced inputs from the periphery, it can be speculated that the PO and also the decoupled cortical areas can fulfill their internal maintenance function during SWS, as proposed in the homeostatic theory of sleep. The same picture holds for the ATN, another higher order nucleus involved in emotional and memory processes (Dalrymple-Alford et al., 2015; Markowitsch, 1988).
As mentioned, the rRTN in rats is reciprocally coupled to the ATN (Aker et al., 2006; Lozsádi, 1994; but not in cats and other species). Moreover, the RTN receives significant projections from the hippocampal formation (Cavdar et al., 2008) and from the amygdala (Zikopoulos and Barbas, 2012), and these interconnected brain regions are considered to be part of the limbic emotion system. The ATN receives also major inputs from the parahippocampal area. Considering the role of the ATN also as part of a memory retrieval system (Dalrymple-Alford et al., 2015), it can be speculated that the increased functional unidirectional coupling between the rRTN and ATN during LSWS and even more during DSWS, which was one of the most pronounced changes observed, supports the occurrence of specific intra-ATN processes and hippocampal-ATN transfer, leading to the reflection or facilitation of either reactivation or consolidation of various types of memories. This process includes emotional memory consolidation, which was proposed to be one of the functions of slow-wave sleep and takes place in the diencephalon (Markowitsch, 1988). This view might fit into a consolidation hypothesis, as was proposed in Diekelmann and Born (2010). Information, stored in the parahippocampal or hippocampal area, might be reprocessed during light and even more during DSWS in the ATN. Whether the increased couplings between rRTN and ATN during slow-wave sleep is indeed allowing or reflecting emotional memory-related processes awaits a functional study in which this hypothesis can be either falsified or verified.
The rRTN and cRTN differ in another aspect: while during PW their reciprocal inhibition is balanced as expressed by similar directed couplings, this picture changes when sleep occurs. The cRTN diminishes its influence on the rRTN at the transition to LSWS, while during DSWS, the rRTN diminishes its influence on the cRTN. The increase in reciprocal independence of the two poles when sleep deepens allows each part of the RTN to modulate or inhibit other thalamic nuclei in their own way, permissive for the functional heterogeneousness of the thalamic nuclei. The discrepancy in functional connectivity within the RTN (between rRTN and cRTN) as sleep deepens is in line with the literature regarding the heterogeneity between the different subparts of the RTN. Nowadays functional heterogeneities are described for the RTN (Cavdar et al., 2002; Crabtree, 2018). The decrease in coupling between the cRTN and rRTN during sleep might suggest that each part can more easily fulfill a different function. The RTN is a unique structure, and its function during wakefulness has been demonstrated in selective attention, and in deviant detection (Yu et al., 2009). During sleep, the RTN produces sleep spindles (Steriade et al., 1987) and in WAG/Rij rats, particularly, the rostral pole of the RTN contributes to the maintenance of this strain of rats typical SWDs (Lüttjohann and van Luijtelaar, 2015; Meeren et al., 2009). It might be argued that the presence of these SWDs hampers the conclusions and the interpretation regarding whether sleep-related changes in functional network connectivity, as found here, are identical as in nonepileptic controls. Therefore, it needs to be investigated whether similar changes can be found in controls as well.
In all, various TC and CT projection neurons as well as the intrathalamic projections are differentially controlling and controlled during wakefulness, LSWS, and DSWS. This allows each of the thalamic neurons to fulfill its own and unique function. The sleep-state-dependent changes in intra-RTN couplings might be leading in these processes.
Methodological issues and study limitations
Directed connectivity was investigated between several structures that are part of the cortico-thalamo-cortical network. Sometimes connectivities were revealed not immediately fitting a well-established anatomical pathway. These connections might be spurious; however, this is unlikely considering that couplings may occur via other structures of pathways than those that are directly connected, including areas not recorded in this study. It can be mentioned, however, that all primary conclusions drawn from the study, including a nongradual decrease in cortico-thalamic coupling and a differential inhibition of first- versus higher order nuclei by the RTN during DSWS, are based on anatomically direct structures.
The study was done in a genetic epileptic rat model and the presence of this neurological condition may present a significant confound to any inferences made in this study. The literature on sleep in patients with absence epilepsy (Dinopoulos et al., 2018) as well in WAG/Rij rats agrees regarding that, in particular, REM sleep is reduced, and that arousal mechanisms are affected, again both in humans and in the presently used rat model (van Luijtelaar and Bikbaev, 2007). The circadian distribution of sleep in WAG/Rij seems normal and at the time of day in which DSWS dominates, SWDs are very rare (Smyk and van Luijtelaar, 2020). Percentages of non-REM sleep do not deviate from normal controls. Moreover, DSWS and SWDs are neurophysiological spoken, not compatible. Although it cannot be excluded that some aspects of DSWS might be different from those of controls, there are no indications that DSWS is qualitatively or quantitatively different or has another function in WAG/Rij rats as in nonepileptic rats.
The study toward the cortico-thalamo-cortical couplings was restricted to the somatosensory cortex and it remains to be investigated, if the reported changes are also representative for different cortical areas.
Footnotes
Authors' Contributions
A.L. and G.v.L. conceived the study. A.L. acquired the data. I.V.S. analyzed the data. All authors interpreted the data and wrote the article.
Author Disclosure Statement
None of the authors has any conflict of interest to disclose.
Funding Information
The Russian Science Foundation, project No. 19-12-00201, supported this work.
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.