
Abstract
Background:
Stress-related disruption of emotion regulation appears to involve the prefrontal cortex (PFC) and amygdala. However, the interactions between brain regions that mediate stress-induced changes in emotion regulation remain unclear. The present study builds upon prior work that assessed stress-induced changes in the neurobehavioral response to threat by investigating effective connectivity between these brain regions.
Methods:
Participants completed the Montreal Imaging Stress Task followed by a Pavlovian fear conditioning procedure during functional magnetic resonance imaging. Stress ratings and psychophysiological responses were used to assess stress reactivity. Effective connectivity during fear conditioning was identified using multivariate autoregressive modeling. Effective connectivity values were calculated during threat presentations that were either predictable (preceded by a warning cue) or unpredictable (no warning cue).
Results:
A neural hub within the dorsomedial PFC (dmPFC) showed greater effective connectivity to other PFC regions, inferior parietal lobule, insula, and amygdala during predictable than unpredictable threat. The dmPFC also showed greater connectivity to different dorsolateral PFC and amygdala regions during unpredictable than predictable threat. Stress ratings varied with connectivity differences from the dmPFC to the amygdala. Connectivity from dmPFC to amygdala was greater in general during unpredictable than predictable threat, however, this connectivity increased during predictable compared with unpredictable threat as stress reactivity increased.
Conclusions:
Our findings suggest that acute stress disrupts connectivity underlying top-down emotion regulation of the threat response. Furthermore, increased connectivity between the dmPFC and amygdala may play a critical role in stress-induced changes in the emotional response to threat.
Impact statement
The present study builds upon prior work that assessed stress-induced changes in the human neurobehavioral response to threat by demonstrating that increased top-down connectivity from the dorsomedial prefrontal cortex to the amygdala varies with individual differences in stress reactivity. These findings provide novel evidence in humans of stress-induced disruption of a specific top-down corticolimbic circuit during active emotion regulation processes, which may play a causal role in the long-term effects of chronic or excessive stress exposure.
Introduction
Exposure to stress has disruptive effects on cognitive and emotional function that may culminate in the development of an anxiety disorder (Antov et al., 2013; Raio and Phelps, 2015). For example, emotion dysfunction can arise following high levels of stress exposure, in part, from the disruption of emotion learning, memory, and regulation processes (Maier and Watkins, 1998). Thus, excessive stress appears to promote the development of stress-related disorders via disruptions of normal emotion processes. Recent studies have begun to identify the neural basis for these stress-induced shifts in emotion regulation, by examining corresponding changes in regional brain activity (Goodman et al., 2018b; Merz et al., 2013). Specifically, these studies suggest that activity within the prefrontal cortex (PFC) and amygdala play an important role in the stress-induced disruption of emotional function. However, stress-induced changes in the function of these brain regions as a network (i.e., functional connectivity) during subsequent emotional learning and memory functioning remain poorly understood. Accordingly, a more detailed understanding of the functional connectivity of this network would offer valuable insight into the neural processes that underlie stress-induced changes in the regulation of the emotional response to threat.
Pavlovian fear conditioning is a procedure that is often used to study emotional processes, including emotional learning, memory, and regulation. In a typical Pavlovian fear conditioning procedure, a warning cue, also called a conditioned stimulus (CS+), is paired with a threat, also called an unconditioned stimulus (UCS). After repeated pairings of these stimuli, the warning cue begins to produce a conditioned response (CR). The CR is often taken as evidence that the association between the warning cue and threat has been learned. However, learning-related changes in the response to threat, also called the unconditioned response, are frequently observed. Conditioned diminution of the response to threat is demonstrated by a diminished threat response when the threat follows a warning cue (i.e., predictable threat) compared with when a threat follows a safety cue or is presented alone (i.e., unpredictable threat) (Domjan, 2005; Harnett et al., 2015; Knight et al., 2010; Rust, 1976; Wood et al., 2012). This conditioned diminution of the threat response provides a quantitative index of the ability to regulate the emotional response to threat (Goodman et al., 2018a). Prior work suggests that the PFC and amygdala underlie stress-induced changes in the conditioned diminution of the emotional response to threat (Goodman et al., 2018b). However, stress-induced changes in the functional connectivity of the brain regions that support threat-related emotion regulation processes remain to be examined. Given that emotion regulation is supported by a network of brain regions, assessment of the dynamic interactions within this network is critical for a better understanding of stress-induced disruption of emotion regulation processes.
Effective connectivity is an approach to functional connectivity that describes the directional influence of one brain region on another and has previously been used to assess task-related functional connectivity underlying cognitive and emotional processes, such as emotion regulation (Wheelock et al., 2014). In particular, effective connectivity techniques based on the multivariate autoregressive (MVAR) model [e.g., Granger causality; Granger (1969)] can determine the directional causal influence of one brain region over another (Deshpande and Jia, 2020; Wang et al., 2017b). In contrast, standard functional connectivity analyses [e.g., Pearson correlations between time-courses; Whitfield-Gabrieli and Nieto-Castanon (2012)] assess nondirectional correlations among regions within brain networks. Thus, effective connectivity analyses extend the assessment of the brain networks that underlie cognitive and emotional processes by describing the directional influence of each brain region on another.
Assessment of the directional influence between brain regions provides greater insight into the information processing supported by neural networks and their impact on cognitive and emotional processes. For example, prior effective connectivity studies have demonstrated that trait anxiety (Wheelock et al., 2014) and early life stress (Grant et al., 2015) are linked to alterations in the effective connectivity of the PFC and amygdala during Pavlovian conditioned diminution of the emotional response to threat. Specifically, early life stress was associated with altered connectivity during unpredictable threats, which was characterized by weaker connectivity from the ventromedial PFC (vmPFC) to the bilateral amygdala (Grant et al., 2015). Also, the directional connectivity between the PFC and the amygdala seems to be disrupted in stress-related disorders such as posttraumatic stress disorder (Rangaprakash et al., 2018). This prior work suggests that trait anxiety and early life stress lead to persistent disruptions in the PFC to amygdala connectivity, impacting the ability to adaptively regulate the emotional response to threat (Grant et al., 2015; Wheelock et al., 2014). Taken together with our prior findings that PFC and amygdala activity is altered by interindividual variability in stress reactivity (Goodman et al., 2018b), we hypothesized that reactivity to stress would correspond to the changes in PFC to amygdala effective connectivity that support conditioned diminution of the emotional response to threat.
The current study investigated effective brain connectivity during Pavlovian fear conditioning following exposure to psychosocial stress. More specifically, effective connectivity among functionally defined neural networks was assessed during Pavlovian conditioned diminution of the emotional response threat. The relationship between stress reactivity and effective connectivity for predictable and unpredictable threats was compared to better understand stress-related disruption of the neural networks that underlie emotion regulation. Accordingly, this assessment will extend prior findings of stress-induced changes in regions (i.e., PFC and amygdala) that mediate the emotional response to threat (Goodman et al., 2018b) by further investigating the stress-altered effective connectivity of this neural network. Further investigation of the effective connectivity between these brain regions will provide new knowledge regarding the stress-related disruption of neural networks that support emotion regulation.
Methods
Participants
Data from 101 healthy right-handed participants (58 males, 43 females; age range = 18–23) who participated in a previous neuroimaging investigation of stress-induced differences in fear conditioning (Goodman et al., 2018b) were analyzed for the present study. All participants provided written informed consent as approved by the Institutional Review Board of the University of Alabama at Birmingham.
Task design
Participants completed functional magnetic resonance imaging (fMRI) during the Montreal Imaging Stress Task [MIST; Dedovic et al. (2005); Goodman et al. (2016); Orem et al. (2019); Wheelock et al. (2016)], which was followed by Pavlovian fear conditioning (Harnett et al., 2015, 2019). Data from the current study were derived from a previously published study that investigated the neural substrates underlying the effects of acute stress on Pavlovian fear conditioning (Goodman et al., 2018b). The current report presents a follow-up analysis to determine the impact of psychosocial stress on effective connectivity during Pavlovian conditioning.
Montreal Imaging Stress Task
Participants completed two separate blood oxygen-level-dependent (BOLD) fMRI scans during a modified version of the MIST, which has been [described in prior work; Goodman et al. (2016); Orem et al. (2019); Wheelock et al. (2016)]. Briefly, participants completed math problems during Control and Stress conditions of the MIST. Before the Control condition, investigators lowered participants' stress level by telling them “It is OK if you do not answer all of the math problems correctly.” During the Control condition, investigators lowered participants' stress level by providing positive auditory feedback and providing sufficient time (i.e., 5 sec) to respond to each math problem. In contrast, stress levels were elevated before the Stress task by emphasizing that questions must be answered correctly. Participants were told that other participants had answered more than 80% of the items correctly and if he/she did not answer at least 80% correct, his/her data would not be used. Furthermore, negative auditory feedback was provided during the Stress task to elevate stress levels. Failure on the task was ensured by modifying the amount of time the participant was given to respond to the problems. Consistent with prior research, the Control condition of the MIST was followed by the Stress condition of the MIST for all subjects (Allendorfer et al., 2014, 2019; Goodman et al., 2016, 2019, 2020; Orem et al., 2019; Wheelock et al., 2016, 2018). Participants began the fear conditioning task ∼25 min after completing the MIST. As indicated by prior research, a 25-min delay between stress induction and Pavlovian fear conditioning is sufficient for stress to affect the subsequent conditioning task (Hermans et al., 2014; Raio and Phelps, 2015).
Pavlovian fear conditioning
Two auditory stimuli (i.e., 700 and 1300 Hz tones; counterbalanced) served as conditioned stimuli (i.e., CS+ and CS−) during the fear conditioning procedure used in the current study. The CS+ always coterminated (i.e., 100% pairing rate) with the UCS (100-dB white noise, 0.5-sec duration), while the CS− was always presented without the UCS (i.e., 0% pairing rate). In addition, the UCS was presented alone on some trials (UCS alone). During two conditioning scans, a total of 72 trials (18-sec intertrial interval [ITI]) were presented. Each scan lasted ∼15 min and included 36 trials per scan (12 CS+, 12 CS−, 12 UCS-alone trials). The order of conditioning trials was pseudorandomly determined (without replacement), such that no more than two trials of any stimulus type (i.e., CS+, CS−, or UCS alone) were presented consecutively. Research questions for this study were focused on the response to threat (i.e., the UCS). Therefore, statistical analyses compared responses with the CS+UCS (i.e., UCS that followed the CS+) versus UCS alone, similar to prior work (Harnett et al., 2015; Knight et al., 2009, 2010, 2011; Wood et al., 2013, 2015). UCS expectancy was assessed during conditioning as a continuous measure of expectation of UCS presentation. Participants were asked to use a rating bar on a 0 (certain the UCS would not be presented) to 100 (certain the UCS would be presented) scale to rate their expectation of UCS presentation on a moment-to-moment basis. These methods and data have been described and presented in previously published work upon which the current connectivity analyses are based (Goodman et al., 2018b).
Psychophysiological and behavioral analysis
Montreal Imaging Stress Task
Following the MIST, participants completed a questionnaire that assessed self-reported stress during the Control and Stress conditions of the MIST (Goodman et al., 2016; Wheelock et al., 2016). Participants rated the applicability of eight statements (e.g., “I felt overwhelmed”) on a 1–5 scale for both Control and Stress conditions. Accordingly, total scores could range from 8 (low stress) to 40 (high stress) for each condition. Stress ratings for the Control and Stress conditions of the MIST were transformed before further analysis. Specifically, each participant's stress ratings were squared (Stress2, Control2) and the differences between Stress and Control conditions (Stress2 − Control2) were used to index individual differences in stress reactivity (Goodman et al., 2018b).
Functional MRI acquisition
Neuroimaging data were collected using a 3T Siemens Allegra scanner with a gradient-echo echo planar pulse sequence (TR = 2000 msec, TE = 30 msec, FOV = 240 mm, matrix = 64 × 64, slice thickness = 4 mm). High-resolution magnetization-prepared rapid gradient echo (MPRAGE) T1-weighted images were collected as an anatomical reference (TR = 2300 msec, TE = 3.9 msec, FOV = 256 mm, matrix = 256 × 256, slice thickness = 1 mm, with a 0.5 mm gap). BOLD fMRI data were collected during the MIST and conditioning tasks, however, results from the MIST have been previously reported (Goodman et al., 2016; Wheelock et al., 2016, 2018) and have not been included in the current report. Furthermore, our prior study reported stress-induced changes in the neural function underlying conditioned diminution of the emotional response to threat (Goodman et al., 2018b). Thus, the current analyses focus on stress-induced changes in effective connectivity during the subsequent Pavlovian fear conditioning task.
Analyses
Information regarding details of BOLD fMRI processing and analyses has been reported previously (Goodman et al., 2018b). Briefly, images were processed using the Analysis of Functional NeuroImages (AFNI) software package (Cox, 1996), using standard preprocessing and first-level modeling techniques (Goodman et al., 2018b). The gamma variate hemodynamic response function was used to model BOLD fMRI data. Data were analyzed at the individual subject level using multiple linear regression. The regression included reference waveforms for the CS+UCS, UCS alone, CS+, and CS−, in addition to nuisance regressors for joystick movement and the six standard head motion parameters. The resultant activation maps were normalized to a standard template (Talairach and Tournoux, 1988) for group-level analyses. Correction for multiple comparisons was completed by applying a family-wise error p < 0.05 cluster-level threshold in all analyses. These analyses, which were completed in the prior study, identified 21 areas of activation in which the fMRI signal to predictable versus unpredictable threat varied with stress reactivity (Goodman et al., 2018b). Foci were identified in both the left and right hemispheres, including coordinates within the anterior cingulate cortex (ACC), dorsal ACC (dACC), dorsolateral PFC (dlPFC), dorsomedial PFC (dmPFC), ventrolateral PFC (vlPFC), vmPFC, amygdala, insula, middle cingulate, posterior cingulate cortex (PCC), and inferior parietal lobule (IPL). Accordingly, these 21 functionally defined foci were chosen for the effective connectivity analyses in the present study (Table 1).
Regions of Interest
Hemisphere, region, and Talairach coordinates of center of spherical ROIs derived from prior work (Goodman et al., 2018a). Numbers within parentheses reflect distinct ROI within a particular brain region [e.g., dorsolateral PFC(2) is the second of three ROIs within the right dorsolateral PFC]. PFC, prefrontal cortex; ROI, regions of interest.
Effective connectivity model
The present study used the MVAR model to assess stress-induced changes in effective brain connectivity during Pavlovian fear conditioning. The MVAR model is based on the principle that causality (i.e., influence of one region A on another region B) can be used to estimate effective connectivity if the present and future values of the time series from region B are predicted by past values of the time series from region A (Granger, 1969). This approach used an MVAR model that has been described previously (Deshpande et al., 2009, 2010a,b; Wheelock et al., 2014, 2018) (Fig. 1). This model has been validated using both simulations (Deshpande et al., 2010c; Wen et al., 2013) and experimental data (Wang et al., 2017a,b).

Effective connectivity analysis pipeline. Based on prior analyses (Goodman et al., 2018b), 21 functionally defined ROI coordinates were selected for the effective connectivity analysis. Nodes (i.e., 8-mm-radius spheres) were constructed around each coordinate using AFNI's 3dcalc. The mean fMRI time series from each node during Pavlovian fear conditioning was extracted for all participants. The time series was deconvolved using a cubature Kalman filter to obtain the latent neuronal response. The neuronal response to and from each node was included in the dynamic MVAR model to obtain the causal connectivity metrics between every pair of nodes. Task-specific connectivity weights were obtained for four different stimulus events (CS+, CS−, CS+UCS, and UCS alone). Single-sample t-tests were performed within trial type to obtain significant connections during CS+UCS and UCS-alone trials. Two-sample t-tests were used to identify connections that were significantly different between CS+UCS compared with UCS alone. Finally, differential path weights (UCS alone − CS+UCS) were compared with self-reported stress reactivity (Stress2 − Control2). AFNI, Analysis of Functional NeuroImages; CS, conditioned stimulus; fMRI, functional magnetic resonance imaging; MVAR, multivariate autoregressive; ROI, region of interest; UCS, unconditioned stimulus. Color images are available online.
Effective connectivity analysis
Using 3dcalc (AFNI), individual masks were generated using the coordinates of the peak voxel from each of the 21 areas of activation (Table 1). Peak voxels served as the center location for the construction of spherical region of interests (ROIs) (radius = 8 mm). The mean time series from the 21 ROIs were extracted for each participant. The mean time series for each ROI was temporally normalized and a cubature Kalman filter was used to determine the latent neuronal state variables by hemodynamic deconvolution of the fMRI time series (Havlicek et al., 2011) (Fig. 1). This model has been shown to be robust against overfitting (Sreenivasan et al., 2015). In addition, since the underlying state equation is defined in continuous time, it can be parameterized at a temporal resolution much finer than the TR (we used TR/10 in this study), potentially allowing the model to estimate the underlying neuronal latent variables at a much finer temporal resolution than the measured BOLD data. Hemodynamic differences in the time series from each ROI were removed via deconvolution because vascular differences across brain regions have the potential to spuriously influence effective connectivity estimates (Rangaprakash et al., 2018; Wheelock et al., 2018). A boxcar function representing input events (CS+, CS−, CS+UCS, UCS alone, and ITI) served as an external input along with the normalized fMRI time series from each ROI (Table 1) for the deconvolution model. The hidden neuronal variables identified by the deconvolution procedure were included in a dynamic MVAR model to calculate the effective connectivity between all ROIs for each participant (refer to prior publications for details of the dynamic MVAR model applied to task data) (Hampstead et al., 2016; Hutcheson et al., 2015; Lacey et al, 2014; Sathian et al., 2013). Task-specific connectivity was calculated by populating each participant's causality values into five different samples based on the timing of each event (i.e., CS+, CS−, CS+UCS, UCS alone, and ITI). Although CS+ and CS− path weights were calculated, the current analysis focused on comparisons between CS+UCS and UCS-alone effective connectivity as a function of stress reactivity. Thus, only CS+UCS and UCS-alone conditions were considered for further statistical analyses. Effective connectivity differences between CS+UCS and UCS-alone conditions were evaluated by first characterizing connectivity during each trial type, and then determining which paths differed between CS+UCS and UCS alone. Connectivity during each trial type was characterized using two separate one-sample t-tests to identify significant pathways during CS+UCS and UCS-alone trials (test value = 0). For descriptive purposes, a relatively less conservative significance threshold (α = 0.05) was used for the one-sample tests. This threshold was implemented on the basis that the one-sample t-tests were calculated to capture and depict the effective connectivity identified in each network. A two-sample t-test was used to determine which paths showed significantly different connectivity during CS+UCS versus UCS-alone trials. To maximize the reliability of connectivity differences that were tested based on the study hypothesis, a more conservative significance threshold (α = 0.01) was used for the two-sample test. Figure 1 provides a depiction of the effective connectivity methods and statistical analyses for this project.
Gephi software was used to summarize the results of the present statistical analyses (Bastian et al., 2009). Graph-theoretic analysis was used to investigate the out-degree of effective connectivity networks. Out-degree was defined as the number of significant out-going paths directed from one ROI to all other ROIs. Furthermore, the relationship between individual differences in stress reactivity (i.e., self-reported stress) and brain connectivity was assessed. Specifically, correlation analyses were used to compare self-reported stress with ROI connectivity path weights on CS+UCS and UCS-alone trials to determine the relationship between the acute stress response and differential effective brain connectivity.
Results
Psychophysiological and behavioral results
Montreal Imaging Stress Task
Questionnaires were used to assess self-reported stress to the MIST. The prior published analysis of these stress ratings demonstrated that psychosocial stress was higher during the Stress MIST than the Control MIST (Goodman et al., 2018b). Furthermore, stress ratings varied with the peripheral emotional response (i.e., heart rate) during Stress compared with Control conditions. Specifically, heart rate increased as stress ratings increased (see Goodman et al., 2018b for further details).
Effective connectivity results
CS+UCS and UCS-alone effective connectivity
Several common neural hubs were identified across CS+UCS and UCS-alone trials by the effective connectivity analysis (Fig. 2). Neural hubs were identified, based on out-degree, within the left insula and bilateral vmPFC during the presentation of the UCS on both CS+UCS and UCS-alone trials. On CS+UCS trials, the left insula, right dACC, right dlPFC, right PCC, and bilateral vmPFC had the highest number of outward projections (out-degree = 20; Fig. 2a) followed by the right middle cingulate (out-degree = 17) and left vlPFC (out-degree = 11). During UCS-alone trials, the highest number of outward projections (out-degree = 20; Fig. 2b) included the left insula, left vlPFC, and bilateral vmPFC, followed by the right dlPFC (out-degree = 9) and right middle cingulate (out-degree = 5). The false-discovery rate (FDR)-corrected effective connectivity of these networks is also depicted in the Supplementary Figure S1.
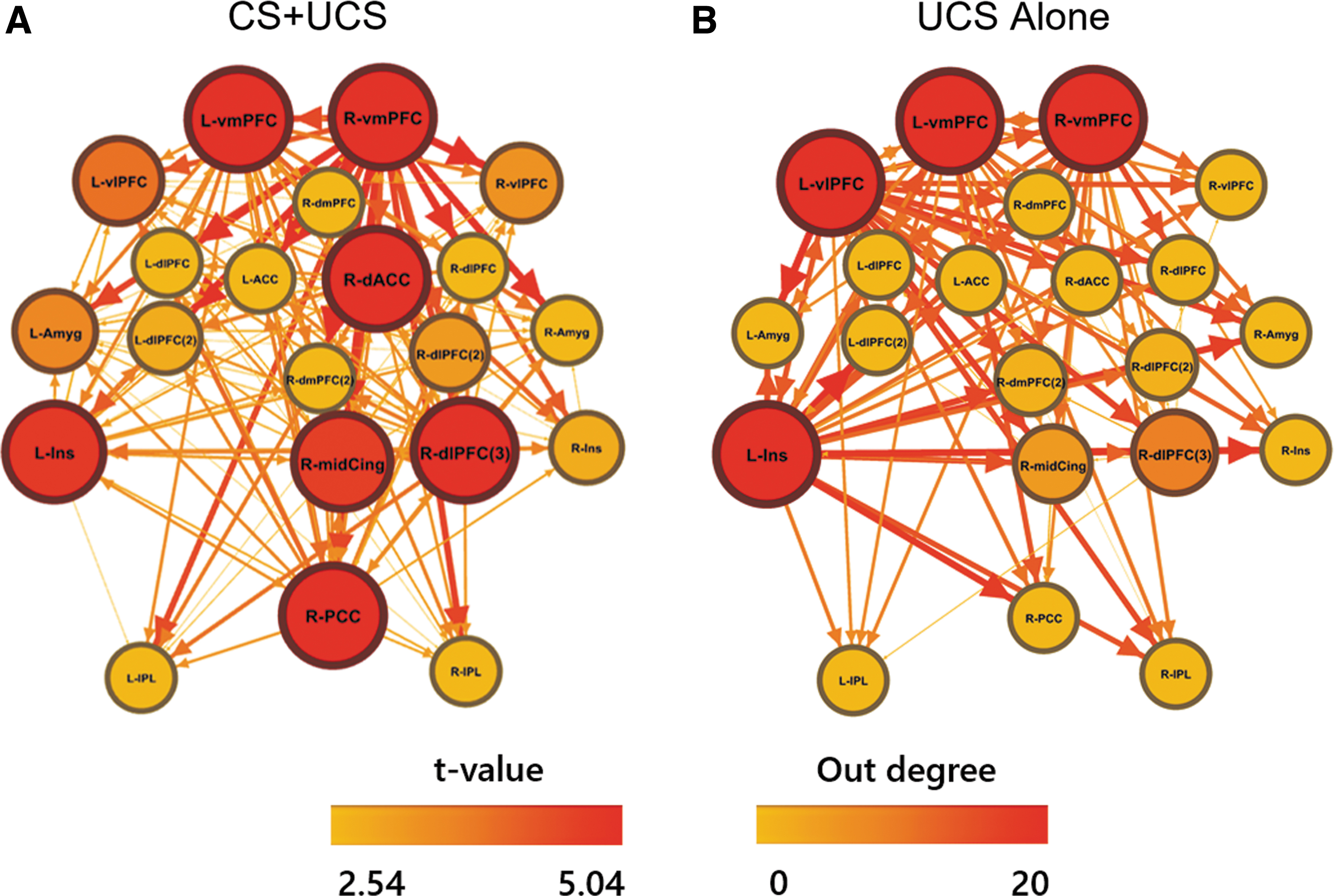
Depiction of effective connectivity path weights during predictable (CS+UCS;
CS+UCS versus UCS-alone effective connectivity
The right dmPFC and dACC showed greater effective connectivity path weights to several other ROIs during CS+UCS compared with UCS-alone trials (Fig. 3a). Furthermore, the right dmPFC also demonstrated the greatest outward projections (out-degree = 12) during CS+UCS compared with UCS-alone trials. Alternatively, the right dmPFC only showed greater effective connectivity path weights to the right dlPFC and amygdala during UCS-alone versus CS+UCS trials (Fig. 3b). Thus, the right dmPFC was identified as the primary hub differentiating effective connectivity during CS+UCS trials compared with UCS-alone trials.
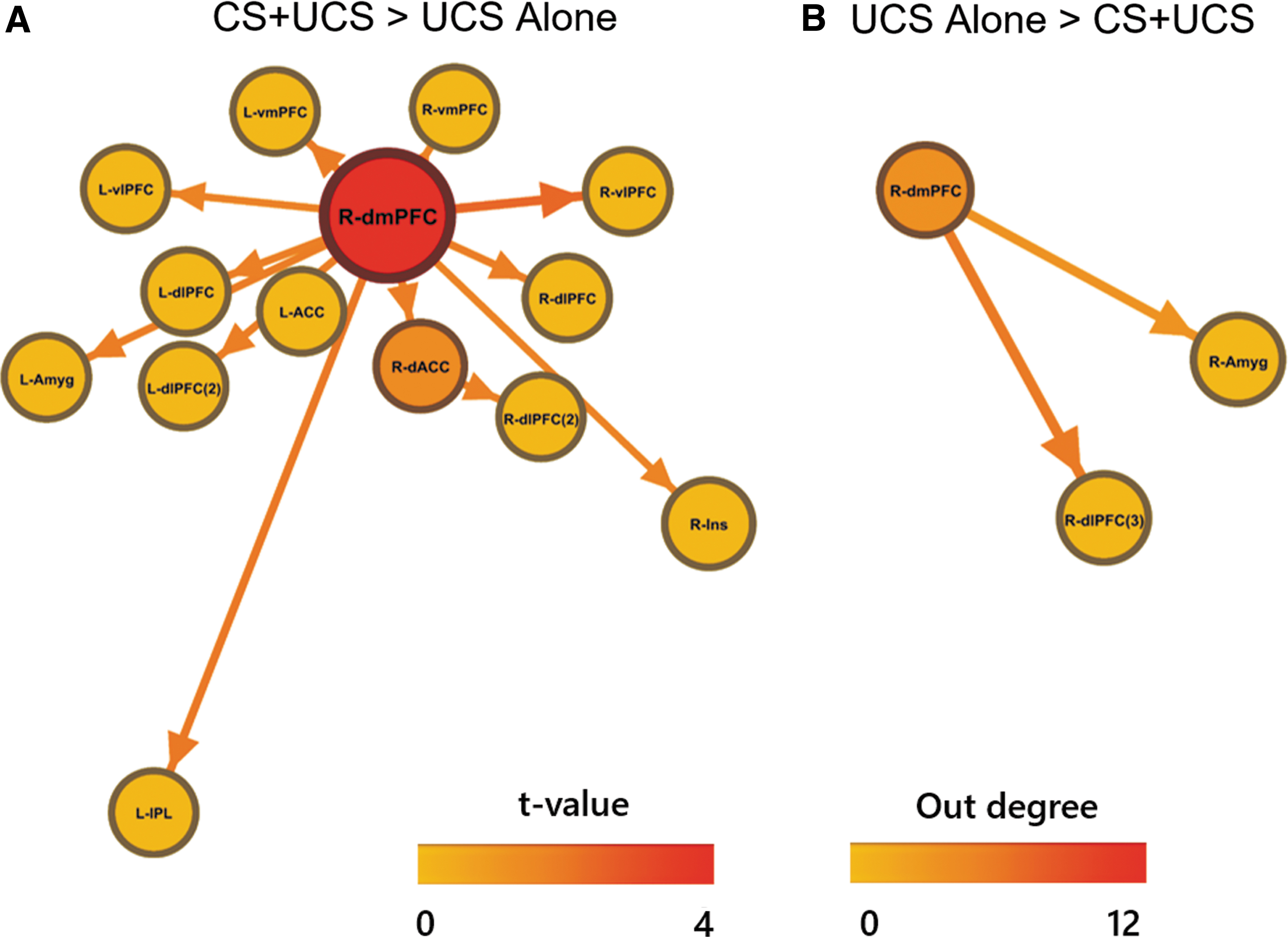
Comparison of effective connectivity path weights during predictable (CS+UCS) versus unpredictable (UCS alone) threat. Greater outward effective connectivity paths from the dmPFC and dACC were observed during predictable threat
Effective connectivity and stress reactivity
Person-to-person differences in brain connectivity and self-reported stress were also investigated. Differential path weights between UCS-alone versus CS+UCS connectivity (obtained from each participant) were correlated with self-reported stress. Differential effective connectivity path weights from the right dmPFC and left IPL showed significant correlations with self-reported stress (Fig. 4 and Table 2). Specifically, connections from the dmPFC node to right dlPFC, vlPFC, amygdala, and IPL nodes were negatively correlated (p < 0.05 uncorrected) with self-reported stress. Likewise, connections from the left IPL node to the right dACC and left dlPFC nodes were negatively correlated (p < 0.05 uncorrected) with self-reported stress. These relationships were not significant when corrected for multiple comparisons (α = 0.05, FDR-corrected).
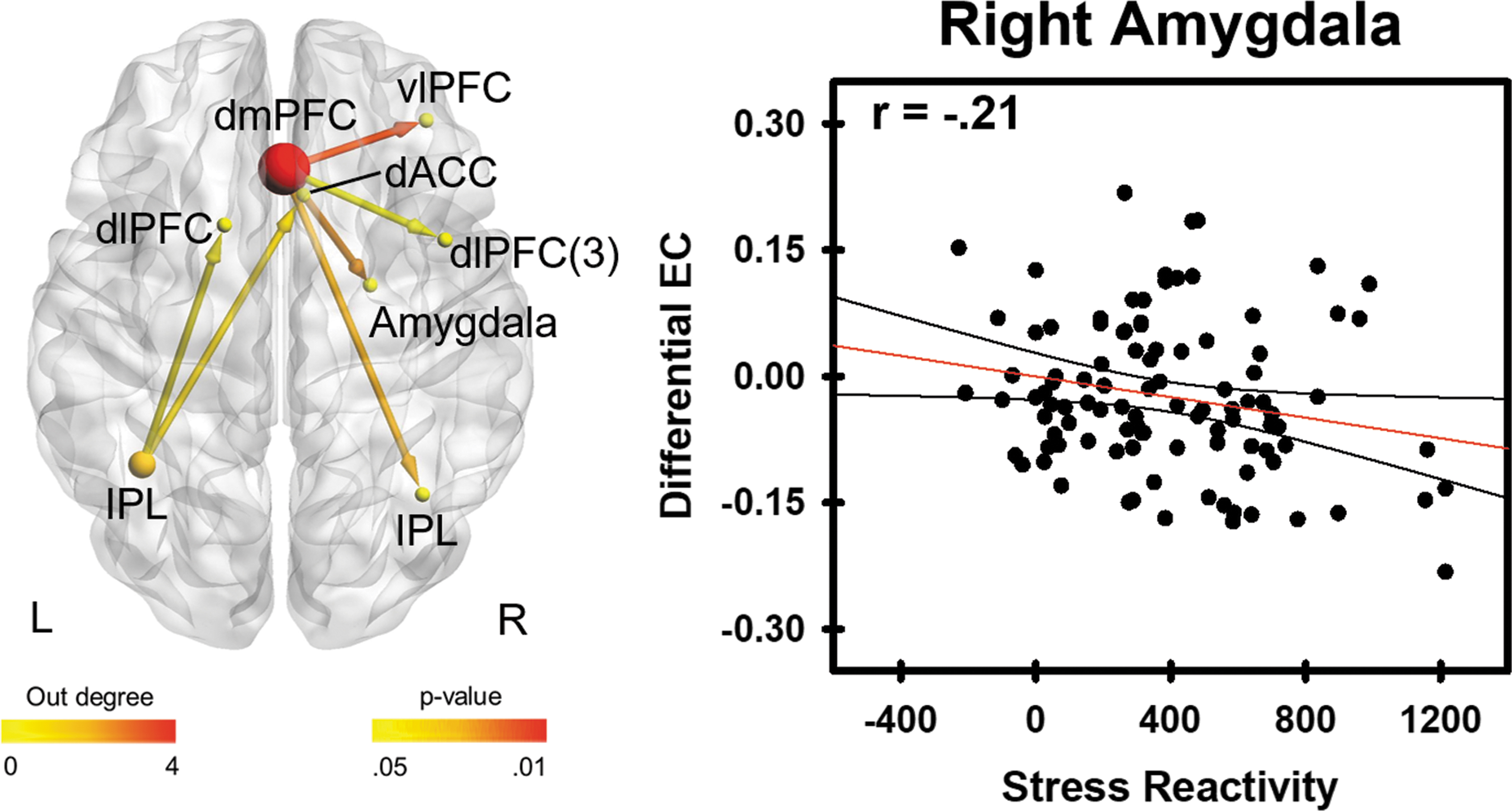
Relationship between self-reported stress reactivity and differential effective connectivity (UCS alone − CS+UCS). The left panel shows that changes in effective connectivity vary negatively with stress reactivity (p < 0.05). The right panel shows data demonstrating the negative relationship between self-reported stress reactivity and differential effective connectivity values between dmPFC and amygdala for unpredictable (i.e., UCS alone) compared with predictable (i.e., CS+UCS) threat trials. Color images are available online.
Correlation of Stress Reactivity (Stress2 − Control2) and Differential Effective Connectivity (UCS Alone − CS+UCS)
CS, conditioned stimulus; dACC, dorsal anterior cingulate; dlPFC, dorsolateral PFC; dmPFC, dorsomedial PFC; IPL, inferior parietal lobule; UCS, unconditioned stimulus; vlPFC, ventrolateral PFC.
Discussion
Stress-related disruption of emotion regulation processes appears to be modulated by the function of the PFC and amygdala. However, the mechanism by which these brain regions interact to mediate these processes remains unclear. Determining the connectivity of these brain regions during the stress-related disruption of the neurobehavioral response to threat may provide novel insights into the neural mechanisms of stress-induced emotional dysfunction. Therefore, the current study assessed the impact of acute stress exposure on the effective connectivity of brain regions that support emotion regulation during Pavlovian fear conditioning. Our prior work has demonstrated that stress modifies the activity of brain regions that support emotion regulation processes (Goodman et al., 2018b). However, the dynamic causal interplay between these brain regions (i.e., effective connectivity) was not assessed. The current study extends our previous findings by conducting follow-up effective connectivity analyses and comparing the data with individual differences in self-reported stress reactivity. These analyses revealed that the left insula and bilateral vmPFC served as neural hubs during the presentation of both predictable and unpredictable threats. However, the right dmPFC demonstrated greater outbound connectivity during predictable than unpredictable threats. Furthermore, comparisons of stress reactivity with differential connectivity (i.e., during unpredictable vs. predictable threats) revealed that dmPFC outbound connectivity varied negatively with stress reactivity. In particular, we found a negative relationship between stress reactivity and differential connectivity from the dmPFC to amygdala, consistent with our hypotheses. Although our interpretation is that acute stress disrupts subsequent brain connectivity underlying emotion regulation processes, it is possible that stress reactivity during the MIST and dmPFC-amygdala connectivity during the threat response are each separately influenced by a third variable (e.g., trait anxiety). The influence of a third variable could produce a spurious correlation between these variables (i.e., stress reactivity and effectivity connectivity between brain regions) that does not reflect a causal relationship. However, the causal influence of acute stress on subsequent emotion regulation processes in humans has been well-documented (Antov et al., 2013; Bentz et al., 2013; Jackson et al., 2006). Furthermore, prior work has identified variability in dmPFC and amygdala function underlying fear conditioning that varies with prior stress reactivity (Merz et al., 2013). Taken together, prior research and the current study results suggest that high stress reactivity may interfere with the regulation of the emotional response by increasing dmPFC connectivity to the amygdala during predictable threats, compared with unpredictable threats. Accordingly, the current study provides a more comprehensive and dynamic systems neuroscience perspective on the disruption of emotion regulation processes that arise from acute stress.
CS+UCS and UCS-alone effective connectivity findings
The bilateral vmPFC and left insula appear to be important neural hubs during both predictable and unpredictable threats in the present study (Fig. 2). These findings portray important similarities in the brain regions that process information about predictable and unpredictable threat. However, the right dmPFC showed greater outward effective connectivity throughout the node network during predictable threats than unpredictable threats (Fig. 3). This finding suggests that dmPFC may support greater top-down regulation during predictable than unpredictable threats and may underlie the conditioned diminution of the emotional response to threat observed in prior work (Domjan, 2005; Dunsmoor et al., 2008; Harnett et al., 2015; Knight et al., 2010; Rust, 1976; Wood et al., 2012). Alternatively, the right dmPFC showed greater outward effective connectivity to the right amygdala and dlPFC during unpredictable threats than predictable threats (Fig. 3). Conditioned diminution of the emotional response to threat is evident when the response to the unpredictable threat is greater than the response to the predictable threat (Goodman et al., 2018a). Thus, increased outbound connectivity from the dmPFC node to the dlPFC and amygdala during the unpredictable threat demonstrates evidence of conditioned diminution of this neural network during the response to predictable threat. Taken together, the present findings suggest the dmPFC may serve as a central node in the neural circuit that regulates the emotional response to threat. Prior work has also suggested that dmPFC projections regulate amygdala activity (Delgado et al., 2008). In turn, amygdala projections to midbrain regions, including the hypothalamus, support the expression of the peripheral emotional response, including sympathetic and parasympathetic activity (Cheng et al., 2003, 2006; Knight et al., 2005; Wheelock et al., 2014; Wood et al., 2012). Thus, the greater effectivity connectivity from dmPFC to amygdala during predictable threats in the current study suggests that the dmPFC plays an important role in the regulation of the amygdala as well as the emotional response to threat, which differs when the threat is predictable compared with unpredictable.
Effective connectivity and stress reactivity findings
The dmPFC and left IPL appear to serve as important neural hubs that support the regulation of the emotional response to threat following a stressful experience (Fig. 4). In particular, differences between connectivity paths (i.e., for unpredictable vs. predictable threat) from the dmPFC to amygdala were negatively correlated with stress reactivity. Our analyses revealed that differential dmPFC connectivity to the right amygdala during predictable threat compared with unpredictable threat decreased as stress reactivity increased. Although connectivity from the dmPFC to amygdala was greater during unpredictable than predictable threat when collapsed across all participants (Fig. 3), this pattern varied as a function of stress reactivity. The finding that dmPFC to amygdala connectivity varies with stress is consistent with our prior work (Goodman et al., 2018b). Specifically, we previously reported that activity within the dmPFC, amygdala, and other brain regions varied with stress reactivity in the same sample of participants. The present findings, however, extend our prior work by demonstrating that these brain regions act as a dynamic neural circuit with functional interactions that change with stress reactivity.
Conclusions
Prior work has identified stress-induced changes in the conditioned diminution of the fMRI response to threat within the dmPFC and amygdala (Goodman et al., 2018b). The present study builds upon this prior work by investigating the causal interplay (i.e., effective connectivity) of these brain regions. The current results revealed that differences in connectivity from dmPFC to amygdala during unpredictable and predictable threat varied negatively with stress reactivity. Thus, functional connectivity from the dmPFC to the amygdala may reflect an important neural process that is disrupted by acute stress, which may in turn disrupt adaptive regulation of the emotional response. Accordingly, the current study provides novel evidence for stress-induced changes in emotion regulation processes that appear to be mediated by altered communication from the PFC to the amygdala.
Footnotes
Acknowledgments
The authors thank Ethan Gossett, Chelsea Tompkins, Tyler Orem, Danielle Hurst, and Kimberly Wood for their personal and technical assistance during the course of the study.
Authors' Contributions
A.M.G. and D.C.K. drafted and revised the article. A.M.G. and M.D.W. performed all the statistical analyses. A.M.G., M.D.W., N.G.H., and E.S.D. prepared the data for analysis. G.D. provided computational training and scripts for the analysis. M.D.W., N.G.H., E.S.D., S.M., and G.D. provided feedback on various versions of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by the National Institute of Mental Health of the National Institutes of Health [Grant No. MH098348].
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.