
Abstract
Introduction:
Deceptive intentions may be realized by imparting false (simple deception) or true (manipulative truth) information. Both forms of deception require inferring others' thoughts and are underpinned by the theory of mind (TOM) neural system. Manipulative truth is thought to more strongly recruit these processes. However, the organization of functional interactions underlying simple deception and manipulative truth remains unclear.
Materials and Methods:
We performed psychophysiological interaction analysis for a key node in the TOM system, the right temporoparietal junction (rTPJ), using functional MRI data obtained from 23 volunteers (14 men and 9 women, age range 18–45 years) during the sender-receiver game. During the game, participants sent true, simple deceptive, or manipulative truthful messages to another player according to their own choice. A Bayesian approach to statistics was employed to perform statistical inference and define voxels with significant changes in functional interactions.
Results:
We observed functional interactions between nodes of the TOM system (bilateral TPJ, left precuneus, left dorsomedial prefrontal cortex, and right superior temporal sulcus) characterizing both forms of deception. We identified an increment in functional interactions of the rTPJ with the left TPJ (lTPJ) and right precuneus associated with manipulative truth. Furthermore, we demonstrated that a higher rate of manipulative truthful actions was associated with weaker functional interactions between the rTPJ and lTPJ, left precuneus, and left dorsomedial prefrontal cortex.
Discussion:
Compared with simple deception, manipulative truth is associated with a higher demand for socio-cognitive processes that contributes to the cognitive load of this form of deception.
Impact statement
This study contributes significantly to our understanding of the fundamental characteristics of socio-cognitive brain mechanisms. Analysis of psychophysiological interactions is highly relevant to the neurophysiology field to promote a shift from studying local activity changes in brain structures to elucidating the activity-related characteristics of functionally connected regions. The presented research for the first time demonstrates functional interactions between nodes of the theory of mind system underpinning deception through communication of false information and telling the truth to deceive (manipulative truth). Understanding the neural substrates of deception may be applied in the forensic field to develop techniques for detecting deception.
Introduction
Deception is an important component of social interactions. It is defined as a successful or unsuccessful deliberate attempt to generate a belief in another person, which the communicator considers wrong (Vrij, 2008). Accordingly, deception encompasses various forms of misleading, even when imparting truthful information, which is termed manipulative truth (Sutter, 2009). Recent studies have demonstrated that this form of deception is more sophisticated because of a higher demand for socio-cognitive processes (Volz et al., 2015; Zheltyakova et al., 2020). Although simple deception is relatively well studied, manipulative truth has not been examined in detail. In this regard, identifying the neurophysiological correlates of sophisticated deception may be relevant for understanding its nature and developing technologies for detecting deception.
Compared with telling the truth, deception is considered a cognitively more demanding task. Indeed, in contrast to remaining honest, the decision to deceive takes longer (Suchotzki et al., 2017) and involves increased neuronal activity in frontal and parietal areas of the human brain (Abe, 2011; Christ et al., 2009). Different components of deception, such as socio-cognitive processes associated with the intention to create a false belief in an interlocutor, may contribute to its higher cognitive load (Vrij, 2008). To attain this goal, one needs to assume thoughts, intentions, and beliefs of the opponent (Bhatt et al., 2010; Lisofsky et al., 2014; Sip et al., 2008, 2012).
In psychophysiological literature, the ability to infer mental states of another person is associated with the theory of mind (TOM) neural system (Bahnemann et al., 2009; Dodell-Feder et al., 2011; Frith and Frith, 2006; Saxe and Kanwisher, 2003). Brain regions repeatedly activated by tasks requiring these socio-cognitive processes constitute nodes of the TOM neural system, comprising the right temporoparietal junction (rTPJ) and left TPJ (lTPJ); precuneus; dorsal, middle, and ventral components of the medial prefrontal cortex; and right superior temporal sulcus (Bzdok et al., 2012; Dufour et al., 2013; Mar, 2011; Molenberghs et al., 2016).
The involvement of a key node in the TOM system, the rTPJ, in deliberate deception in an interactive social context was demonstrated by a meta-analytic study (Lisofsky et al., 2014). Furthermore, compared with truth-telling in a strategic game against a human opponent, both deception by manipulative truth and deception by false messages (simple deception) were associated with increased blood-oxygen-level-dependent (BOLD) signals in areas of the TOM system: the rTPJ, right precuneus (Volz et al., 2015; Zheltyakova et al., 2020), lTPJ, left precuneus, and right superior temporal sulcus (Zheltyakova et al., 2020). Moreover, compared with nonmanipulative control actions, both manipulative truth and simple deception were characterized by increased functional interactions between the rTPJ and left middle frontal gyrus (MFG) (Kireev et al., 2017). Therefore, deception accomplished by imparting both false and true information requires higher cognitive effort and involves a greater load on TOM-related socio-cognitive processes.
Compared with manipulative truth, deception is considered a cognitively more demanding action that requires greater executive control, thus eliciting greater neural responses in areas associated with this function (Ding et al., 2014; Kireev et al., 2013). However, manipulative truth is thought to involve a greater load on socio-cognitive processes, which has led researchers to name this phenomenon “sophisticated deception” (Sutter, 2009). In contrast to simple deception, “sophisticated deception” elicits increased BOLD signals in nodes of the TOM system, including the bilateral TPJ, left precuneus, and right superior temporal sulcus (Volz et al., 2015; Zheltyakova et al., 2020). Thus, the literature suggests that simple and sophisticated deception differs in their cognitive demand relative to that for honest behavior. Nevertheless, it is challenging to tease apart this distinction solely using data from brain activation studies. Therefore, this study aimed to investigate the organization of distant neural interactions underlying simple deception and manipulative truth focusing on functional interactions of the TOM network.
Several functional MRI (fMRI) studies have analyzed changes in functional interactions between brain areas underlying deception and reported an enhancement of coupling (Jiang et al., 2015; Kireev et al., 2015, 2017; Knyazeva et al., 2020). Previous analyses of functional interactions associated with “sophisticated deception” did not identify significant changes when comparing “sophisticated” and simple deception, but both forms of deception were associated with increased interaction of the rTPJ with the prefrontal cortex (Kireev et al., 2017). This may have been owing to the experimental paradigm, which did not focus specifically on social interactions given that subjects played with computer opponents.
Therefore, this research has focused on previously unexplored changes in functional interactions with nodes of the TOM system to obtain a deeper understanding of the neural correlates of different forms of deception. Efforts have been made to characterize deception by imparting false and true information in the settings of a strategic game in social conditions (Volz et al., 2015; Zheltyakova et al., 2020).
We hypothesized that simple deception and manipulative truth would elicit greater coupling between nodes of the TOM system compared with honest actions. Furthermore, if the strength of functional interactions of TOM network nodes represent the degree of involvement of socio-cognitive processes, superior ability to manipulate others and a higher tendency to lie by reporting formal truth would be associated with a greater load on the TOM network and enhanced functional integration of TOM network activity. This is corroborated by the fact that taking the perspective of others is crucial for deception (Kireev et al., 2017; Lisofsky et al., 2014; Sip et al., 2008; Volz et al., 2015; Zheltyakova et al., 2020) and only children who have developed TOM ability are able to lie (Lewis and Osborne, 1990; Polak and Harris, 1999). Therefore, functional interaction of TOM network nodes during “sophisticated deception” may be greater than that during simple deception.
Based on the assumption that deception is a cognitively more demanding task than telling the truth, simple deception would elicit greater functional interactions between frontal areas, parietal areas, and basal ganglia (regions traditionally associated with deception) compared with manipulative truth. Indeed, studies have suggested that TOM ability in manipulative truth is associated with the involvement of cognitive control processes (Leng et al., 2019). For instance, behavioral TOM test scores were positively correlated with N450 amplitude recorded during the condition of sophisticated deception (Carrión et al., 2010). These findings may reflect greater involvement of cognitive control and conflict processing when processing manipulative truth and may also decrease the occurrence of this form of deception.
Two opposing hypotheses regarding the individual variability in the tendency to deceive can be formulated. First, greater involvement of socio-cognitive processes and hence greater functional integration between brain areas for manipulative honest actions (sophisticated deception), including TOM system nodes, could be associated with an increased rate of manipulative truth. This would support the view that a greater ability to attribute mental states to others is associated with a bias to use truth to mislead others. Second, greater functional interactions of TOM system nodes underscoring manipulative truth could be associated with a decreased rate of manipulative truthful actions. This would corroborate the notion that the involvement of socio-cognitive processes increases cognitive load.
Thus, the objective of the study was to specify the neural substrates and functional characteristics underlying TOM-related socio-cognitive mechanisms of simple deception and manipulative truth. We hypothesized that, compared with telling the truth, deception by imparting false and true information would be characterized by increased functional connectivity with the nodes of the TOM system. Furthermore, compared with simple deception, manipulative truth would be associated with increased functional interactions between areas of the TOM network. Simple deception would elicit greater functional interactions between frontal areas, parietal areas, and basal ganglia (regions traditionally associated with deception) compared to manipulative truth. Finally, we tested two opposite hypotheses regarding the role of manipulative truth-associated functional connectivity between TOM network nodes, whether its increase would be associated with higher or lower rate of manipulative truth.
Materials and Methods
Participants
This study used fMRI data previously collected from 33 volunteers (19 men and 14 women, age range 18–45 years). All participants were native Russian speakers without any history of psychiatric and neurological diseases or current medication intake. Handedness was assessed with the Edinburgh Handedness Inventory (Oldfield, 1971). Only right-handed volunteers were included in the study. After conclusion of the experiment, participants received a monetary reward (1000 rubles) and an additional reward of up to 300 rubles based on the results of the experimental game. All procedures were performed in accordance with the Declaration of Helsinki and were approved by the Ethics Committee of the N.P. Bechtereva Institute of the Human Brain, St. Petersburg, Russia. Before commencing the study, all participants provided their written informed consent.
Stimuli and procedure
The experimental design has been described previously by Volz and colleagues (2015) and Zheltyakova and colleagues (2020). Stimulus presentation, recording of responses, and synchronization with the fMRI registration were performed using NordicNeuroLab и E-Prime software (version 1.1; Psychology Software Tools, Inc., Pittsburg, Pennsylvania, USA).
During the experiment, participants in the scanner played a strategic game against an opponent. The main goal of the game was to maximize their reward by sending truthful or deceptive messages to influence the opinion of the opponent. The players were informed that the purpose of the experiment was to study decision-making processes in interactive game settings. Furthermore, they were instructed that they would be competing with a human; however, this information was misleading. All the actions of the opponent were randomly generated by a computer.
The game consisted of 90 trials, each comprising several stages. Participants first viewed a table with two options of monetary rewards, both for themselves and for the opponent. Options were designated either blue or red (Fig. 1a). Participants were required to send a message to the second player, stating either “Blue option is more profitable to You” or “Red option is more profitable to You,” by pressing the corresponding left or right button. In each trial, one of the options was more profitable for the opponent, which could be the same or different option to the one more profitable for the participant. Therefore, the participant had to switch between sending true and false message to maximize gains. Finally, participants were required to respond to the question “Which state do you expect the receiver to choose? The red column or the blue column?” by pressing the corresponding left or right button on the controller. A new trial would then commence.
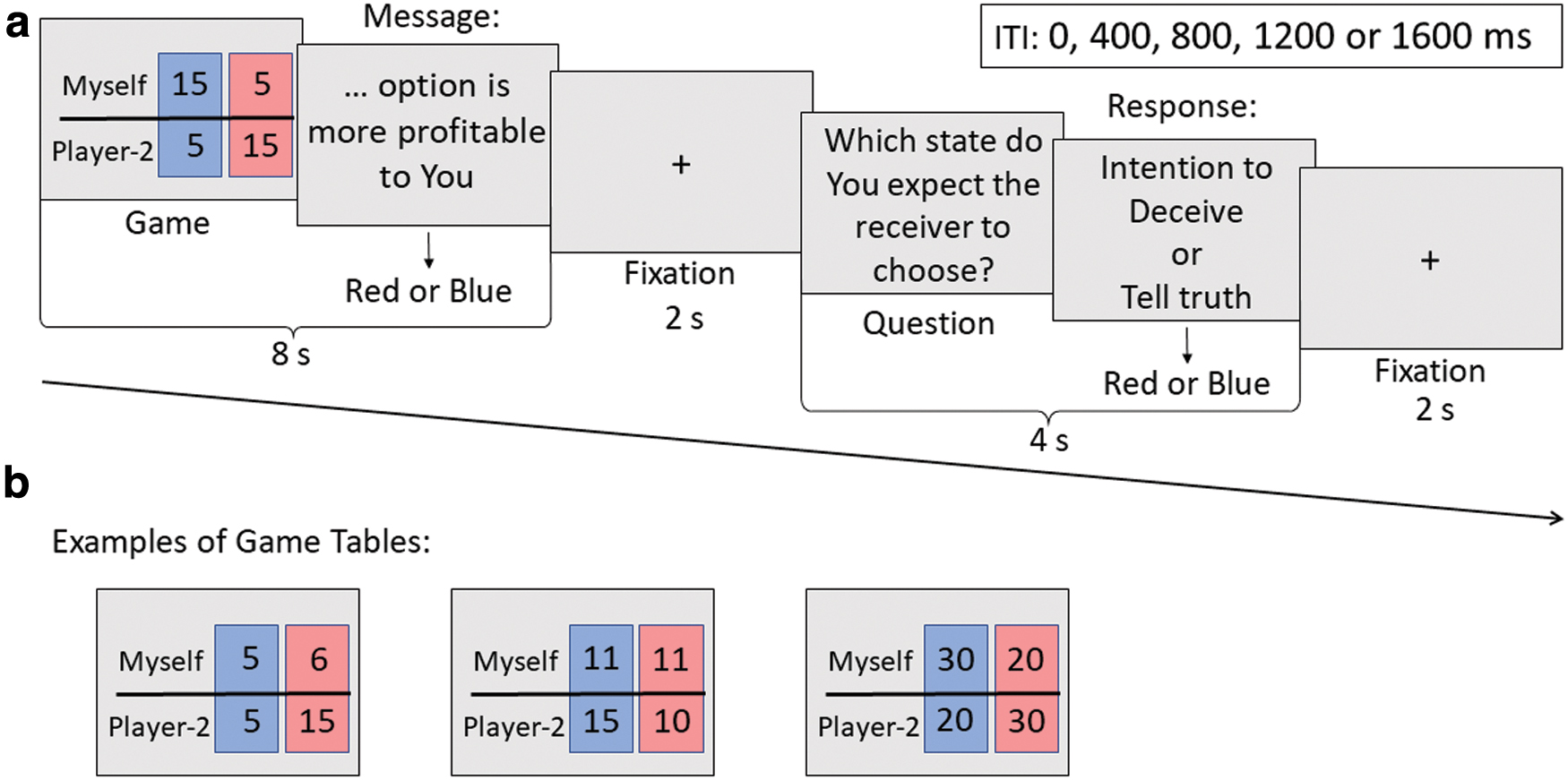
Participants were informed that the opponent was unable to view the tables and distribution of monetary rewards but would receive a message and select the red or blue option accordingly. This choice determined the result of the trial. No feedback was presented during the game to prevent learning and strategy adjustment. Therefore, participants could not be caught sending false messages and were free to choose between being truthful or deceptive. Of note, when sending a truthful message, participants would occasionally try to mislead the second player. Responses to the second question reflected the intentions of participants and allowed us to distinguish messages sent with and without manipulative intentions. After all 90 tables were presented, one table was randomly selected, and participants received an additional monetary reward according to the result of that trial.
The incentive to deceive changed during the game, as tables varied in distributions of game points between two players (Fig. 1b). In 45 of 90 trials, the option that was more profitable for the participant was less profitable for the opponent and vice versa. In 27 trials, only the payoff for the opponent changed between both options, whereas the participant received the same amount of game points in both cases, which could be higher, lower, or the same as the payoff of the opponent. In the remaining 18 trials, the same option was more profitable for both players. During the game, tables were presented to participants in random order.
The temporal characteristics of trials are illustrated in Figure 1a. Trials lasted for 16 sec. For the first 8 sec, the table with distributions of monetary rewards was presented on the screen, and participants were instructed to make a decision and press a button to send the message during this time. A fixation cross was then presented for 2 sec. The question “Which state do you expect the receiver to choose? The red column or the blue column?” then appeared on the screen for 4 sec, during which participants were required to press a button to select the response. A fixation cross was then shown for 2 sec, after which a new trial began. The intertrial interval varied between 0, 400, 800, 1200, and 1600 msec in a randomized manner.
The experiment comprised three types of trials: Truth, Simple Deception, and Manipulative Truth (Fig. 2). The trial was named Truth if the participant correctly informed the opponent about the more profitable option (sent a true message) and expected the same option to be chosen. In Simple Deception trials, the participant sent a false message and expected it to be chosen by the opponent. In Manipulative Truth trials, a true message was sent, but the participant expected the opponent not to believe this information and to select the opposite (false option). Trials in which participants sent a false message and expected the opponent to choose the opposite (true option) were not classified in this study and were used as conditions of no interest in the statistical analysis. Trials in which participants failed to send a message and/or respond to the question were regarded as errors.

Trial classification. “Message” refers to the information sent to the opponent by the first button press: “Red/Blue option is more profitable to You.” “Question” refers to the answer to the second question: “Which state do you expect the receiver to choose? The red column or the blue column?” Reprinted with permission from Zheltyakova and associates (2020).
fMRI image acquisition procedure and image processing
fMRI data were recorded using a 3 Tesla Philips Achieva scanner. Structural images were acquired using a T1-weighted pulse sequence (T1W-3D-FFE; repetition time [TR] = 2.5 msec; echo time [TE] = 3.1 msec; 30° flip angle), measuring 130 axial slices (field of view [FOV] = 240 × 240 mm; 256 × 256 scan matrix) of 0.94 mm thickness. Functional images were obtained using an echo planar imaging (EPI) sequence (TE = 35 msec; 90° flip angle; FOV = 208 × 208 mm; 128 × 128 scan matrix). In total, 32 continuous 3.5-mm-thick axial slices (voxel size = 3 × 3 × 3.5 mm) covering the entire cerebrum and most of the cerebellum were oriented with respect to structural images. The images were acquired using a TR of 2000 msec.
An MR-compatible cervical collar was used to prevent head movements. Data preprocessing and subsequent statistical analysis were performed in the SPM12 toolbox run in MATLAB (Mathworks, Inc., Natick, MA, USA). The preprocessing of raw fMRI data for each participant included the following stages: realignment, slice-time correction, co-registration, normalization, and smoothing.
Psychophysiological interaction analysis and region of interest selection
The generalized form of psychophysiological interaction analysis (gPPI-analysis) was used to determine the organization of functional interactions between brain areas (McLaren et al., 2012). This analysis was performed for every voxel in the brain to detect regions that changed their functional interactions with the predefined region of interest (ROI) in a context-dependent manner. The gPPI toolbox was applied to run the gPPI analysis.
Given the focus of the present study on socio-cognitive mechanisms of deception, the rTPJ was selected as the ROI for gPPI analysis. The ROI represented a sphere with the center at MNI coordinates x = 42, y = 49, z = 20 and radius = 4 mm.
In a previous analysis by Zheltyakova and associates (2020), these coordinates were identified as peak coordinates in a cluster with a significantly increased BOLD signal associated with the intention to deceive (simple deception and manipulative truth/“sophisticated deception”) compared with that for the intention to tell the truth (truth). However, previous analyses have not identified significant differences in the BOLD signal in the rTPJ between simple deception and manipulative truth (“sophisticated deception”) experimental conditions. A potential explanation for this is that the differential involvement of the rTPJ in both forms of deception may only manifest through changes in functional interactions. Therefore, gPPI analysis was conducted to test this hypothesis.
Psychophysiological interaction analysis
Statistical analysis was first performed for each participant separately, and individual general linear models (GLMs) were generated. To create gPPI regressors in these models, BOLD signal time series from the ROI were deconvolved with the hemodynamic response function (HRF) to reveal underlying neuronal activity (Gitelman et al., 2003). The obtained signal was multiplied by the temporal characteristics of experimental events: Truth, Simple Deception, Manipulative Truth trials, unclassified trials, errors, and questions (responses to questions: “Which state do you expect the receiver to choose? The red column or the blue column?”). Events were modeled with the onset at the time of sending messages or responding to questions (button press) and duration equal to zero. Results obtained for multiplication vectors were then convolved with the standard HRF.
GLMs also included regressors of no interest, including: (1) regressors, modeling BOLD signal changes, used in the classical analysis and obtained by multiplying temporal characteristics of experimental events by the standard HRF; (2) head movement parameters, obtained during preprocessing (realignment); and (3) regressors, representing time series of the mean BOLD signal in all voxels in the ROI. Beta values of regression coefficients for the gPPI regressors in GLMs were estimated from individual-level gPPI analysis. For these coefficients, t-contrasts were calculated, including Truth > baseline, Simple Deception > baseline, Manipulative Truth > baseline, and Manipulative Truth > Simple Deception.
To test hypotheses regarding the relationship between the individual rate of Manipulative Truth and strength of functional interactions within the TOM system, we performed correlation analysis using a one-sample t-test. We used the percentage of Manipulative Truth experimental trials over the total number of trials obtained individually for each participant as the covariate of interest.
A Bayesian approach to statistics was used to perform a voxel-wise statistical inference. A zero-effect size threshold and posterior probability threshold defined as log-odds >3 were applied in accordance with reports by Zheltyakova and associates (2020) and Volz and associates (2015). Xjview Toolbox was used to identify the anatomical location of voxels with significant changes in functional interactions.
To identify clusters within the TOM neural system, statistical maps of seven TOM-related regions (the rTPJ and lTPJ; precuneus; dorsal, middle, and ventral components of the medial prefrontal cortex; and right STS) were used [Dufour et al. (2013)]. The cluster was considered a TOM region if the coordinate of its maximum lied within one of seven clusters of the TOM neural system. REX toolbox was applied to illustrate differences in values of regression coefficients in obtained clusters of changes in functional interactions. The gPPI analysis results were illustrated using MRIcroGL toolbox.
Results
Behavioral results
For the statistical analysis, data from 23 participants (14 men and 9 women) were used. In total, 10 excluded volunteers did not perform all types of experimental trials (Truth, Simple Deception, and Manipulative Truth); of these, six participants did not perform a sufficient number of Manipulative Truth trials, and four participants did not perform a sufficient number of Simple Deception trials. Therefore, their game strategy did not fit the concept of social interactions and did not align with the purpose of the study.
The behavioral data analysis included a comparison of the time taken to send messages in different types of trials, termed response times (RTs). Mean RTs for Truth, Simple Deception, and Manipulative Truth trials were 2565 msec (standard deviation [SD] = 726 msec), 2817 msec (SD = 872 msec), and 3091 msec (SD = 1190 msec), respectively. Paired Student's t-tests revealed that RTs were significantly shorter for Truth trials than for Simple Deception trials (t = 4.32, p < 0.001) and Manipulative Truth trials (t = 4.01, p < 0.001). No significant differences in RTs were observed between Simple Deception and Manipulative Truth trials (t = 1.39, p = 0.2). These findings suggested that telling the truth without the intention to deceive took less time regardless of the way the deception was fulfilled (i.e., via false or true messages).
PPI results
Functional interactions underscoring the intention to deceive (simple deception and manipulative truth) versus intention to tell the truth
Compared with truthful actions, actions with deceptive intentions were associated with increased functional interactions between the ROI in the rTPJ and other TOM system structures ([Simple Deception + Manipulative Truth] > [Truth trials]), including the lTPJ and rTPJ, dorsomedial prefrontal cortex, precuneus, and superior temporal sulcus (Fig. 3 and Table 1).

Results of the group level gPPI analysis with the ROI in the rTPJ. Clusters of the increased functional interactions with the rTPJ, associated with Simple Deception and Manipulative Truth, compared with Truth, are shown in red color (effect size = 0 Hz, log odds = 3, k > 40). The TOM system clusters are shown in green color, according to results of the meta-analysis (Dufour et al., 2013). L/R, left/right hemisphere; g., gyrus; TPJ, temporoparietal junction; gPPI analysis, generalized form of psychophysiological interaction analysis; ROI, region of interest; rTPJ, right temporoparietal junction; TOM, theory of mind. Color images are available online.
Results of the Group Level Generalized Form of Psychophysiological Interaction Analysis for the Contrast “the Intention to Deceive (Simple Deception and Manipulative Truth)> Truth” (Effect Size = 0 Hz, Log Odds >3, k > 40)
k, cluster size in voxels; L/R, left/right hemisphere; g., gyrus; TPJ, temporoparietal junction; TOM, theory of mind; *, clusters with coordinates of maximums lying within TOM-related brain regions (Dufour et al., 2013).
Functional interactions characterizing manipulative truth versus simple deception
To test hypotheses concerning changes in functional interactions depending on the form of deception (by relating false or true information), t-contrasts of Manipulative Truth and Simple Deception conditions were used. Compared with Simple Deception, Manipulative Truth was characterized by increased functional interactions between the ROI in the rTPJ and TOM system structures, including the lTPJ and right precuneus (Fig. 4 and Table 2). These areas also exhibited increased functional coupling with the rTPJ in association with the intention to deceive compared with the intention to tell truth, as reported in the section “Functional interactions underscoring the intention to deceive (simple deception and manipulative truth) versus intention to tell the truth” (Table 1). Compared with Manipulative Truth, Simple Deception was characterized by an increase in functional interactions between the ROI in the rTPJ and one area in the brainstem (midbrain) (Table 2).
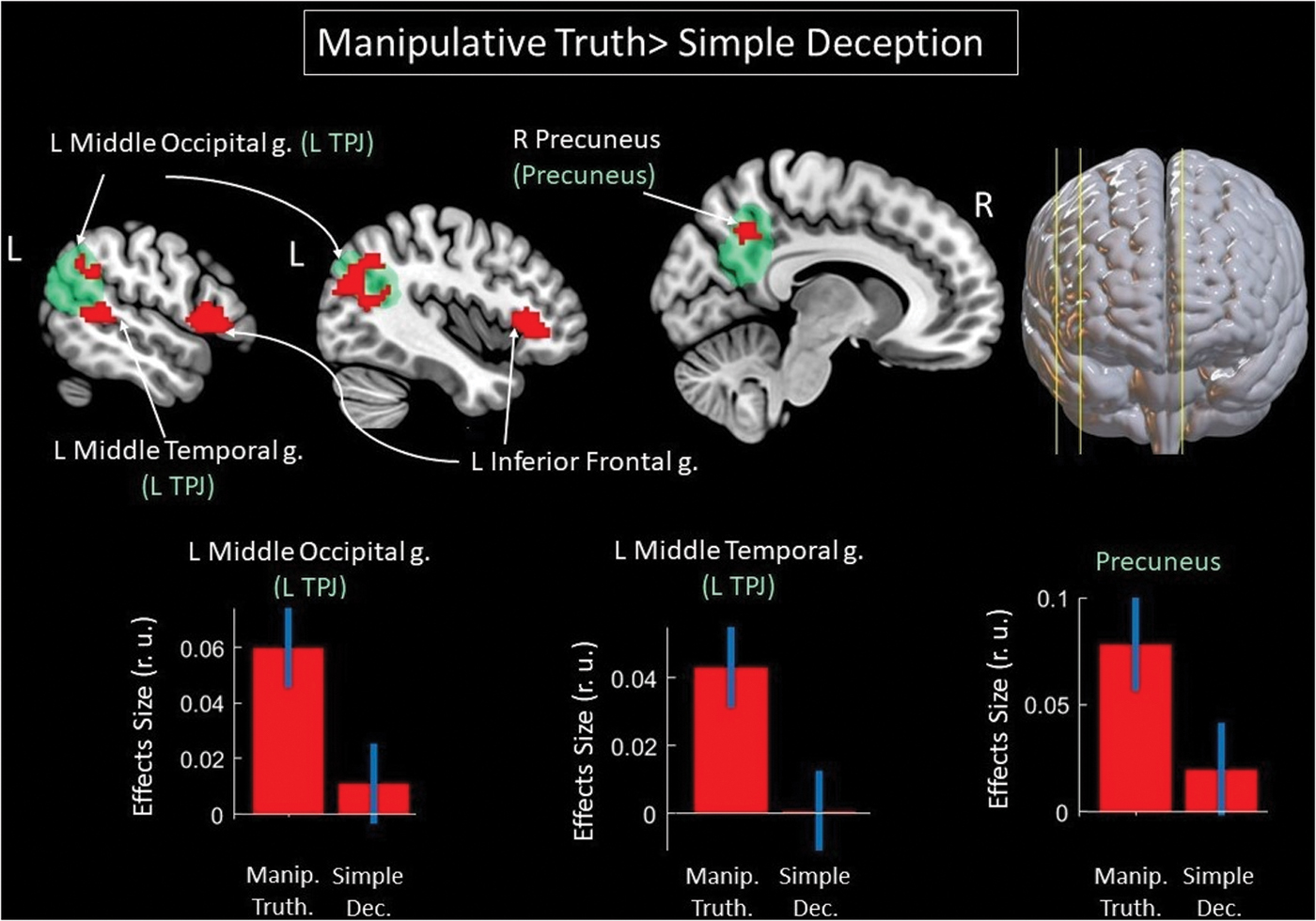
Results of the group level gPPI analysis with the ROI in the rTPJ. Clusters of the increased functional interactions with the rTPJ, associated with Manipulative Truth, compared with Simple Deception, are shown in red color (effect size = 0 Hz, log odds = 3, k > 30). The TOM system clusters are shown in green color, according to results of the meta-analysis (Dufour et al., 2013). L/R, left/right hemisphere; g., gyrus. Color images are available online.
Results of the Group Level Generalized Form of Psychophysiological Interaction Analysis for the Contrast Between Manipulative Truth and Simple Deception (Effect Size = 0 Hz, Log Odds = 3, k > 30)
k, cluster size in voxels; L/R, left/right hemisphere; g., gyrus; *, clusters with coordinates of maximums lying within TOM-related brain regions (Dufour et al., 2013).
Relationship between the rate of manipulative truth and strength of functional interactions characterizing manipulative truth
Correlation analysis of the rate of Manipulative Truth and PPI parameters obtained from the comparison of Manipulative Truth > Simple Deception revealed a negative correlation between the rate of Manipulative Truth and strength of functional interactions between the ROI in the rTPJ and left precentral gyrus, left supramarginal gyrus, right cerebellum, left precuneus, left hippocampus, and left superior frontal gyrus (Fig. 5 and Table 3). Among these structures, the left, left precuneus, and dorsomedial prefrontal cortex were related to the TOM system. These areas exhibited increased functional interactions with the rTPJ during both forms of deception (Simple Deception and Manipulative Truth) compared with that during acting without the intention to deceive (Truth). These interactions were stronger during Manipulative Truth than during Simple Deception (Fig. 5). Positive correlations of the rate of Manipulative Truth and PPIs revealed in the Manipulative Truth>Simple Deception comparison for the rTPJ ROI were observed with the right medial occipitotemporal gyrus, left medial occipitotemporal gyrus, left superior frontal gyrus, right paracentral lobule, and left claustrum; these areas are not nodes within the TOM system.

Results of the group level gPPI analysis with the ROI in the rTPJ. Clusters, in which increased functional interactions with the rTPJ characterizing Manipulative Truth, compared with Simple Deception, negatively correlated with the rate of Manipulative Truth, are shown in red color (effect size = 0 Hz, log odds = 3, k > 30). The TOM system clusters are shown in green color, according to results of the meta-analysis (Dufour et al., 2013). L/R, left/right hemisphere; g., gyrus. Color images are available online.
Results of the Group Level Generalized Form of Psychophysiological Interaction Analysis, Demonstrating the Relationship Between the Rate of Manipulative Truth and Changes in Functional Interactions of the Region of Interest in the Right Temporoparietal Junction Associated with the t-Contrast “Manipulative Truth > Simple Deception” (Effect Size = 0 Hz, Log Odds = 3, k > 30)
k, cluster size in voxels; L/R, left/right hemisphere; g., gyrus; PPI, psychophysiological interaction; *, clusters with coordinates of maximums lying within TOM-related brain regions (Dufour et al., 2013).
Discussion
This study reports for the first time the functional interactions between TOM system nodes, including the bilateral TPJ, left precuneus, left dorsomedial prefrontal cortex, and right superior temporal sulcus, that are associated with both simple and sophisticated forms of deception. Our results support the hypothesis that, compared with telling the truth, deception by imparting both false and true information is characterized by a greater load on socio-cognitive processes. This finding corresponds to previously observed greater local brain activity in loci of the TOM system, including the rTPJ, right precuneus (Volz et al., 2015; Zheltyakova et al., 2020), lTPJ, left precuneus, and right superior temporal sulcus (Zheltyakova et al., 2020). These findings collectively suggest that socio-cognitive brain mechanisms underscore deceptive intentions through changes in local neuronal activity and an increase in distant neural interactions within the TOM neural system.
In addition to TOM nodes, several brain regions previously reported to be involved in deceptive actions, including the left caudate nucleus and inferior frontal gyrus (IFG), exhibited increased functional interactions. The current finding expands upon previous results by revealing the involvement of these areas in the execution of deceptive intentions (both simple and sophisticated forms of deception) by increasing functional connectivity with the rTPJ. The caudate nucleus has been identified in studies on simple deception that analyzed local BOLD signal changes (Harada et al., 2009; Kireev et al., 2013; Lee et al., 2010; Ofen et al., 2017; Yin and Weber, 2019; Yu et al., 2019; Zheltyakova et al., 2020) and functional interaction analysis (Kireev et al., 2015; Yin and Weber, 2019). Several studies have suggested its role in social interactions (Fett et al., 2014, 2019; Kemp et al., 2013; King-Casas et al., 2005; Lemmers-Jansen et al., 2018, 2019; Martin et al., 2016; Myznikov et al., 2021; Sakaiya et al., 2013; Speer and Delgado, 2020; Wardle et al., 2013; Williams et al., 2005). Therefore, the involvement of the caudate nucleus may be associated with an increased load on socio-cognitive processes underpinning simple deception and manipulative truth. The IFG is implicated in multiple aspects of executive control in deception (Christ et al., 2009; Kireev et al., 2015, 2017; Vartanian et al., 2013; Yin and Weber, 2019). In this regard, increased interactions between the rTPJ and lIFG may reflect the interplay between TOM system nodes and executive control neural systems underlying the intention to deceive.
Results of the gPPI analysis for the comparison between two forms of deception confirmed the assumption that, compared with simple deception, manipulative truth is associated with a higher demand for socio-cognitive processes. This manifested as changes in functional connectivity within the TOM system, that is, increased functional interactions of the rTPJ with the lTPJ and right precuneus. These results partly concur with previously reported increases in the BOLD signal in TOM system nodes underscoring manipulative truth compared with that for simple deception (Volz et al., 2015; Zheltyakova et al., 2020).
In contrast to simple deception, manipulative truth is characterized by an increased demand for additional processes other than socio-cognitive processes, as evidenced by the increased functional interaction of the rTPJ ROI with the lIFG and MFG. In deception research, the MFG is typically associated with general cognitive functions (Christ et al., 2009; Ding et al., 2013, 2014; Farah et al., 2014; Ito et al., 2011; Kireev et al., 2017; Pornpattananangkul et al., 2018; Sip et al., 2010; Vartanian et al., 2013; Yin and Weber, 2019; Yin et al., 2016), whereas the IFG is associated with executive control (Christ et al., 2009; Kireev et al., 2015, 2017; Vartanian et al., 2013; Yin and Weber, 2019). This indicates that changes in local neuronal activity and reorganization of distant neural interactions intrinsic and extrinsic to the TOM neural system underscore the execution of manipulative truth.
This study demonstrates that compared with simple deception, manipulative truth requires greater integration within the TOM system and of TOM system nodes with brain regions associated with executive control and general cognitive functions (IFG and MFG). However, in individuals who resort to manipulative truth more often, functional interactions between loci of the TOM system associated with manipulative truth (compared with those associated with deceiving by false information) are weaker. This may be associated with less cognitive resources recruited during deception by telling the truth and its facilitation in these individuals. These results support the hypothesis that functional interactions between TOM network nodes are linked to cognitive control processes involved in performing manipulative truth.
Moreover, individuals exhibiting a higher rate of manipulative truth demonstrate higher functional integration of the rTPJ with areas outside the TOM system, including the left superior frontal gyrus associated with executive control functions such as working memory (Alagapan et al., 2019; Boisgueheneuc et al., 2006). A potential heuristic for these findings is that a higher rate of manipulative truth is associated with a deceased demand for socio-cognitive processes and increased demand for cognitive control required to deceive by telling the truth. Further research is warranted to clarify the roles and brain mechanisms underlying socio-cognitive processes and cognitive control involved in the performance of manipulative truth.
One of limitations of the current study is that gPPI analysis does not allow to reveal directions of observed functional connections. Further research using different methods of analysis would expand upon present findings. Another limitation is that this article dealt specifically with socio-cognitive mechanisms and the rTPJ was selected as the ROI for gPPI analysis. Future studies with broader list of ROIs are needed to reveal interactions between socio-cognitive mechanisms and other neural systems underpinning simple deception and manipulative truth.
Conclusion
In conclusion, we have specified the neural substrates and functional characteristics underlying TOM-related socio-cognitive mechanisms of simple deception and manipulative truth. Our results support the hypothesis that, compared with telling the truth, deception by imparting false and true information is characterized by higher load on socio-cognitive processes. This was reflected by increased functional interactions between the rTPJ and nodes of the TOM neural system (bilateral TPJ, left precuneus, left dorsomedial prefrontal cortex, and right superior temporal sulcus) observed in this study.
Furthermore, compared with simple deception, manipulative truth was associated with a higher demand for socio-cognitive processes. This manifested in changes in functional connectivity within the TOM system, that is, increased functional interactions of the rTPJ with the lTPJ and right precuneus.
Finally, a decrease in manipulative truth-associated functional connectivity of the rTPJ with TOM network nodes in the lTPJ, left precuneus, and dorsomedial prefrontal cortex was associated with a higher rate of manipulative truth. Our findings corroborate the notion that the involvement of socio-cognitive processes is associated with the cognitive load of manipulative truth.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the state assignment of the Ministry of Education and Science of Russian Federation [theme number AAAA-A19-119101890066-2].