
Abstract
Objective:
Absence seizures are the prototypic primarily generalized seizures, but there is incomplete understanding regarding their generation and maintenance. A core network for absence seizures has been defined, including focal cortical and thalamic regions that have frequency-dependent interactions. The purpose of this study was to investigate within-frequency coupling and cross-frequency coupling (CFC) during human absence seizures, to identify key regions (hubs) within the absence network that contribute to propagation and maintenance.
Methods:
Thirteen children with new-onset and untreated childhood absence epilepsy had over 60 typical absence seizures during both electroencephalography-functional magnetic resonance imaging (fMRI) and magnetoencephalography (MEG) recordings. The spatial map of the ictal network was defined using fMRI and used as prior information for MEG connectivity. A multilayer network approach was used to investigate within-frequency coupling and CFC for canonical frequency bands. A rigorous null-modeling approach was used to determine connections outside the noise floor.
Results:
Strong coupling between beta and gamma frequencies, within the left frontal cortex, and between the left frontal and right parietal regions was observed. There was also strong connectivity between left frontal and right parietal nodes within the gamma band. Multilayer versatility analysis identified a cluster of network hubs in the left frontal region.
Interpretation:
Cortical regions commonly identified as being critical for absence seizure generation (frontal cortex, precuneus) have strong CFC and within-frequency coupling between beta and gamma bands. As nonpharmacologic treatments, such as neuromodulation, become available for generalized epilepsies, detailed mechanistic understanding of how “diffuse” seizures are generated and maintained will be necessary to provide optimal outcomes.
Impact statement
This study provides functional magnetic resonance imaging (fMRI)-informed magnetoencephalography multilayer network analyses showing beta/gamma cross-frequency and within-frequency coupling in the frontoparietal and frontofrontal regions during childhood absence seizures, which has not previously been reported for any primary generalized seizure. These results offer a mechanistic explanation for previous fMRI and neurophysiology studies showing focal areas of the precuneus and frontal lobe being critically important for the initiation and maintenance of generalized rhythmic spike wave discharges. As nonpharmacologic treatments, such as neuromodulation, become available for generalized epilepsies, detailed mechanistic understanding of how “diffuse” seizures are generated and maintained will be necessary to provide optimal outcomes.
Introduction
Childhood absence seizures are the prototypic primarily generalized type, but there is incomplete understanding of the mechanisms responsible for the generation and propagation of these seizures. While they remain categorized as generalized (Fisher et al., 2005), absence seizures have been reported to involve networks that are broad but include focal abnormal functioning in bilateral thalami, basal ganglia, brainstem, frontal cortices, temporal cortices, precuneus, and cingulate gyri (Bai et al., 2010; Betting et al., 2006a,b; Carney et al., 2010, 2012; Holmes et al., 2004; Luttjohann et al., 2013; Masterton et al., 2013; Moeller et al., 2008; Salek-Haddadi et al., 2003; Tenney et al., 2013, 2014).
A lack of pathophysiologic understanding severely limits the treatment options for patients with drug-resistant generalized epilepsies. Surgical treatments for epilepsy have begun to evolve beyond traditional resection of brain tissue to more targeted therapies including neuromodulation, which could represent a new treatment paradigm for generalized epilepsies (Benbadis et al., 2018; Cukiert et al., 2020; Dell et al., 2019; Sisterson and Kokkinos, 2020). However, development of these approaches would require a mechanistic understanding of how “diffuse” seizures are generated and maintained.
Our previous work has demonstrated that the onset of human absence seizures is characterized by distinct parietal and frontal corticothalamic networks operating simultaneously (Tenney et al., 2013, 2014; Xiang et al., 2015; Youssofzadeh et al., 2018). In addition, we have shown specific frequency-dependent connectivity patterns that are associated with medication responsiveness in patients with childhood absence epilepsy (Tenney et al., 2018). Specifically, participants with ethosuximide (ETX) unresponsiveness had decreased connectivity in the precuneus region and increased connectivity in the frontal cortex, compared with ETX responders. Our hypothesis regarding absence seizure generation is that cortical sites, such as the precuneus region, are necessary to transform normal network connections into a pathologic and prolonged oscillatory state, which is supported by the frontal corticothalamic network. However, the mechanisms by which brain networks with different frequency oscillations interact to produce absence seizures remain unclear.
Cross-frequency coupling (CFC) is a way to describe the interactions between slow and fast oscillations, and it may help to characterize the mechanisms of communication across various spatial and temporal scales (Hyafil et al., 2015). However, to our knowledge, CFC has not been applied specifically to absence seizures. It has been suggested that CFC is a key feature in maintaining the inhibition/excitation balance and permitting information flow across the brain. While the application of CFC has shown to be powerful and reveals new insights into brain function, most of this work has been focused on cognitive functioning in otherwise healthy brains (Aru et al., 2015; Helfrich et al., 2016; Tewarie et al., 2016; Weaver et al., 2016). Only a few studies have applied these methods to human epilepsy, and none for primary generalized seizures (Amiri et al., 2016; Guirgis et al., 2015; Ibrahim et al., 2018; Jiang et al., 2019; Zhang et al., 2017). However, CFC during epileptic spasms has repeatedly been shown between pathologic high-frequency oscillations and delta slow waves with high-modulation index following perinatal stroke associated with the development of infantile spasms (Iimura et al., 2018; Nariai et al., 2011; Suzuki et al., 2021).
In this study, we utilize a multilayer network approach to investigate within-frequency coupling and CFC, to identify key regions (hubs) that participate in absence seizure propogation and maintainence, potentially at various timescales.
Methods
Participants
Children aged 6–12 years with new-onset childhood absence seizures were recruited from the Neurology Division at Cincinnati Children's Hospital Medical Center (13 participants [7 females, 6 males], mean age at diagnosis, and testing of 8.8 years [range = 6.4–11.8]). This research protocol received Institutional Review Board approval before study recruitment. The parent of each participant provided informed consent, and child. Inclusion criteria included a diagnosis of childhood absence seizures, normal development (based on parental report), normal neurological examination (which grossly assessed cognition), normal brain magnetic resonance imaging (MRI), and bilaterally synchronous 3–4 Hz spike and wave discharges on a normal background with at least one electroclinical seizure lasting 3 sec or more. Exclusion criteria included a history of seizures other than absence type (i.e., generalized tonic/clonic or myoclonic) or the presence of antiepileptic medication.
Electroencephalography-functional MRI
All participants had simultaneous electroencephalography (EEG) and functional MRI (fMRI) recording (Tenney et al., 2018). Echo planar imaging (EPI) was used with blood oxygenation level-dependent (BOLD) weighting and full brain coverage (repetition time [TR]/echo time = 2000/38 ms, field of view 24 × 24 cm, matrix 64 × 64, slice thickness = 4 mm, resulting in a voxel size of 3.75 × 3.75 × 4 mm, 37 axial slices). In addition, a three-dimensional (3D) T1-weighted 1 mm isotropic anatomical image with fiducial markers was acquired for coregistration with both the fMRI and magnetoencephalography (MEG) data. MRI images were collected on a 3T Phillips Achieva MRI scanner using a 32-channel head coil. EEG was recorded using a 68-channel MRI-compatible EEG acquisition system (Maglink by Neuroscan; Division of Compumedics Ltd., El Paso, TX).
Following recording, scanner and cardioballistic artifacts were removed using automated procedures (Scan 4.3 software; Compumedics Neuroscan, Charlotte, NC). EEG data were notch filtered at 60 Hz and EPI gradient artifacts were averaged over the first 9 TR periods and then subtracted from each epoch of the raw data following methods that have been previously described. A standard bipolar EEG montage with 16 channels was used to identify the absence seizures, which were then used for creating epochs for fMRI data analysis. EEG-fMRI was recorded in 10-min blocks (300 EPI volumes) while participants were watching a movie.
MEG recordings
Simultaneous EEG and MEG recordings were conducted using a 275-channel (axial gradiometers) CTF MEG system (CTF Systems Inc., Coquitlam, BC, Canada) with a sampling rate of 4 kHz (Tenney et al., 2013, 2018). Participants were in a supine position throughout the recording. Continuous head localization was used to ensure <5 mm of movement for any recording. Continuous MEG recordings were completed as 5-min sessions, repeated 8 times for a total recording duration of 40 min.
Network definition
EEG-fMRI analysis began with participant EEGs that were reviewed by an experienced epileptologist (J.R.T.) and generalized spike wave (GSW) bursts lasting ≥3 sec were included in the subsequent analysis (Tenney et al., 2018). The fMRI images were preprocessed using statistical parametric mapping version 8 and included spatial realignment to correct participant motion, slice timing to align slices within each volume to the first slice, normalization into Montreal Neurological Institute space, and spatial smoothing with a Gaussian kernel that was full-width half-maximum equal to 8 mm. An event-related independent component analysis (ICA) method was used to detect the brain areas responsible for the absence seizures identified on EEG (Masterton et al., 2013). This method consists of two processing steps—first, a linear deconvolution that provides the BOLD response at each voxel for the time of interest, and second, an ICA that determines the spatial extent of the event-related BOLD changes. The ICA decomposition was performed separately for each participant based on his or her temporally concatenated fMRI data. In other words, each participant's event-related data were concatenated rather than the entire raw fMRI time series.
Group-wise spatial maps were constructed based on an ICA decomposition of this concatenated event-related data. These spatial maps show positive and negative changes in z statistic maps (p < 0.05, family-wise error correction) with respect to the time course for each of the five independent components. Only components with reliable event-related time courses, across the entire group, were used to spatially represent the absence seizure network. The between-subject variance in the estimated time courses was used to help separate true physiologic components from artifact. So, components were included if they showed a consistent group response shape accounting for a significant amount of the total temporal variance (F-test, p < 0.05, corrected for the number of components).
Parcellation
The absence seizure network was spatially defined for subsequent MEG connectivity using the group-wise fMRI activation maps. Areas with BOLD time courses that positively correlated with the seizure time course included the inferior/middle/posterior frontal, temporal, and thalamus. The posterior cingulate, parietal, angular, precuneus, and occipital regions were found to have a negative correlation with the seizure time course. First, the fMRI group activation maps were parcellated for each of the five identified components. These activation maps were binarized so that only suprathreshold voxels remained and this activation was multiplied by a random 200 U parcellation scheme (Craddock et al., 2012). The size of this parcellation allows for a good balance between anatomical interpretations and functional homogeneity. Centroids of parcels with >10 active voxels (each 2.0 mm isotropic) provided the spatial coordinates of the seizure network nodes.
fMRI-informed MEG connectivity analysis
MEG preprocessing was carried out using FieldTrip routines running in MATLAB (Oostenveld et al., 2011). A total of 25 sec of absence seizure data were used from each participant, using the fewest seizures possible per subject. So, the integer value (rounding down) of the longest seizure was extracted, and then, the integer value of the next longest seizure was added, etc., until a total of 25 sec of seizure activity was collected, per subject. The data were filtered from 0.5 to 100 Hz, and a very sharp discrete Fourier transform filter was used to suppress power line noise at 60 Hz. The data were then epoched into 1s trials, before source localization. Fiducial locations were identified on each subject's 3D-T1, which was then segmented before construction of a realistic single-shell headmodel (Nolte and Muller, 2010; Nolte et al., 2008). The absence network, represented as nodes in template space, was nonlinearly warped to individual space using SPM12 routines. Neuronal population activity at each node was estimated using a linearly constrained minimum variance beamformer, with 0.1% regularization (VanVeen et al., 1997). Noise was also projected through the spatial filter, which was used to normalize estimated source activity (i.e., we computed the “neural activity index,” at each position).
MEG within-frequency coupling and CFC with surrogate normalization
Source-level activity (41 nodes) was downsampled to 1000 Hz and symmetrically orthogonalized to eliminate the impact of source leakage on connectivity estimates (Colclough et al., 2015). The data were filtered into canonical frequency bins: 1–4 Hz (delta), 4–8 Hz (theta), 8–12.5 Hz (alpha), 12.5–30 Hz (beta), and 30–59 Hz (gamma). Amplitude envelopes were calculated using absolute Hilbert transform and Z-transformed to stabilize variance. Amplitude/amplitude coupling (AAC) was obtained for each node pair both within- and between-frequency bins (Brookes et al., 2012, 2016; Mandke et al., 2018; O'Neill et al., 2015; Tewarie et al., 2016). For between-frequency coupling, only low-to-high frequency AAC was computed.
One hundred surrogate time series were generated to obtain null distributions of AAC, used to compute z-statistics for the observed AAC for each node pair, both within and between frequencies. The null modeling involved iterative amplitude-adjusted Fourier transform with a maximum of 100 iterations for convergence (Schreiber and Schmitz, 1996). These surrogates underwent exactly the same filtering and AAC analyses, as the observed data. This approach resulted in 15 adjacency matrices weighted by z-statistic (5 within-frequency and 10 between-frequency matrices).
Multilayer network analysis
Connectivity matrices were thresholded by the orthogonal minimum spanning tree method (Dimitriadis et al., 2017). Our previous work has shown an advantage in using this data-driven method over proportional/sparsity approaches (Williamson et al., 2021). In between-frequency matrices, the maximum connectivity was taken between each pair of nodes to ensure that the matrices remained symmetric and undirected. Adjacency matrices were then combined to form a full multilayer network, represented by a supra-adjacency matrix (Fig. 1; De Domenico et al., 2013). Multilayer versatility (i.e., multilayer PageRank centrality) was obtained for each node and averaged across participants. All MEG connectivity, surrogate analyses, and multilayer analyses were performed with in-house scripts and the MuxViz package in R (De Domenico et al., 2015; R Core Team, 2018).
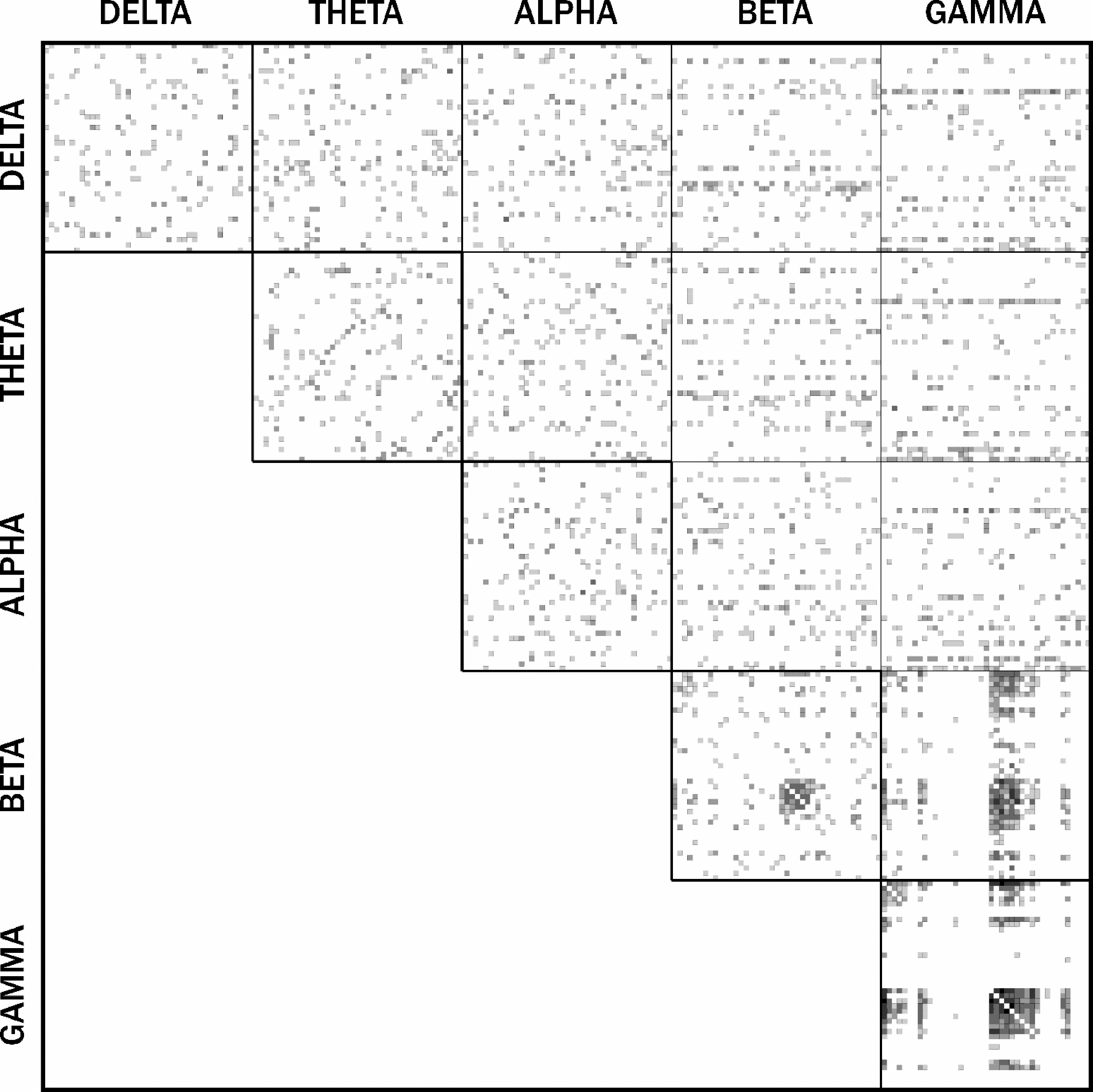
Supra-adjacency matrix of absence seizures. For the between-frequency matrices, the maximum connectivity was taken between each pair of nodes to ensure that the matrices remained symmetric and undirected. Clusters of cross-frequency coupling can be seen between beta and gamma bands with increased within-frequency connectivity in the gamma band.
Results
Participants
Sixteen participants were recruited and completed the study procedures. Three participants were excluded from CFC analysis since the total duration of seizures recorded was <25 sec. The remaining 13 participants (7 females, 6 males) had a mean age at diagnosis and testing of 8.8 years (range = 6.4–11.8). These participants had an average of 4.9 seizures (range = 2–8) lasting an average of 11.2 sec, each (range = 3.3–21.3; Table 1). All seizures used for analysis were spontaneous and not provoked by hyperventilation. All participants had MEG recording first followed by EEG-fMRI. Nine participants had both recordings on the same day and four participants had the recordings on separate days (range of 1–7 days apart).
Participant Characteristics
MEG, magnetoencephalography; R, right; SD, standard deviation.
MEG connectivity
Supra-adjacency matrices were binarized, summed, and thresholded such that only connections present in three or more participants were passed on for visualization. All analyses were carried out at the individual level but summarized for the group. We observed strong coupling between beta and gamma frequencies, within the left frontal cortex, and between the left frontal and right parietal regions. There was also strong connectivity between the left frontal and right parietal nodes within gamma (Fig. 2A).
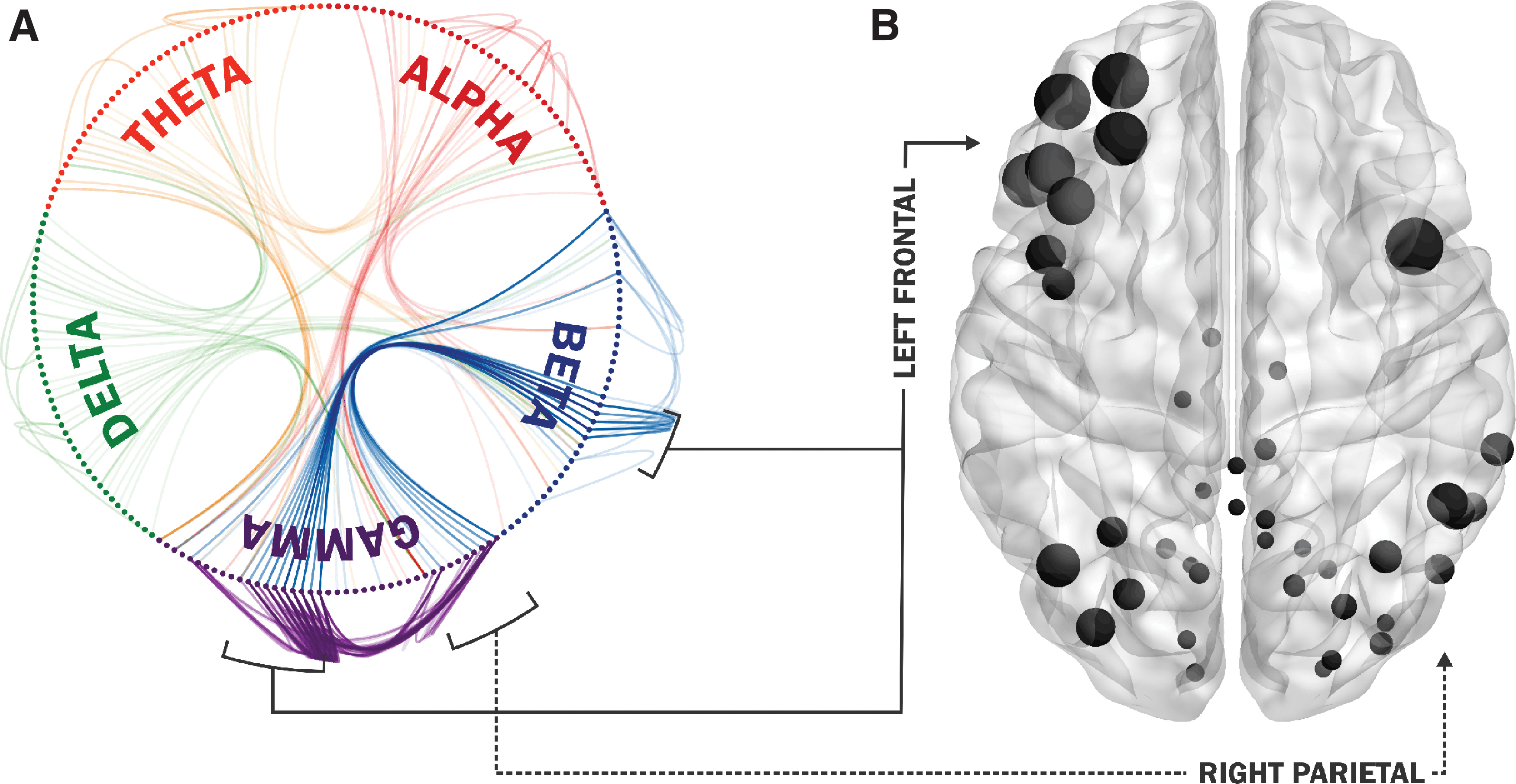
Magnetoencephalography absence network connectivity.
Multilayer versatility
Group-wise multilayer versatility analyses identified a dominant cluster of network hubs in the left frontal cortex, specifically the left frontal gyrus (Table 2). Other nodes with high versatility were observed diffusely in cortical regions of the absence seizure network (Fig. 2B).
Absence Seizure Nodes Ranked by Versatility
L, left; MNI, Montreal Neurological Institute; R, right.
Discussion
The findings reported in this study that beta/gamma amplitude-envelope CFC in the frontofrontal and frontoparietal regions is present during childhood absence seizures provide new insight into the pathophysiology of human absence seizures and support the view that absence epilepsy is regulated by multiple brain networks coupled in the frequency domain. This helps bring together several previous neuroimaging and neurophysiology findings related to absence seizure generation and propagation (Bai et al., 2010; Carney et al., 2010, 2012; Luttjohann et al., 2013; Masterton et al., 2013; Moeller et al., 2008; Ossenblok et al., 2019; Vaudano et al., 2009). Specifically, we have shown that cortical regions commonly identified as being critical for absence seizure generation (frontal cortex, precuneus) have strong CFC and within-frequency coupling between the beta and gamma bands. These results may have important future clinical treatment implications since surgical epilepsy may eventually evolve to include neuromodulatory paradigms for drug-resistant generalized epilepsies.
Our study identified clear cross-frequency and within-frequency amplitude envelope correlations between the frontofrontal and frontoparietal lobes during absence seizures. We used a network approach that was multilayered and included both within-frequency and cross-frequency analyses to investigate integration of multiple frequency band-specific information in the dynamics of absence seizures. A rigorous null-modeling approach ensured that only connections outside the noise floor were identified. A recent report of MEG connectivity during absence seizures found commonality between all participants with focal patterns of high local connectedness within the frontal and parietal areas during the maintainence of spike wave discharges (Ossenblok et al., 2019). Our findings add to this understanding by suggesting that focal frontofrontal and frontoparietal regions of CFC between the beta and gamma bands may coordinate the control mechanisms associated with the maintainence of absence seizures, leading to absence seizure generalization. This likely reflects the maintenance of the diffuse rhythmic 3 Hz GSW discharges, primarily perpetuated by the left frontal region. In addition, the right parietal nodes were found to have higher versatility due to ongoing connectivity with the left frontal nodes.
Previous fundamental work has demonstrated that the frequency of neuronal oscillations is a main determinant of the distance over which neuronal populations can communicate (Brunel and Wang, 2003; Kopell et al., 2000). Low-frequency oscillations tend to facilitate synchronization over long distances, while high-frequency activity is optimal for rapid information flow over short distances (Aru et al., 2015; Buzsaki, 2006). CFC provides a different method to investigate the mechanisms of information flow, across multiple timescales, during human absence seizures. Our data show the clear advantage to performing multilayer connectivity analyses for patients with epilepsy, since only the within-frequency gamma connectivity would have been captured in tradition, monolayer analysis.
There are several limitations of the current study to consider. Our analysis was limited by the small size of the current cohort and these group level results allow no conclusions to be drawn regarding CFC differences between participants. This is an important area of future investigation as our group and others have shown that treatment response is associated with differences in connectivity during absence seizures (Ossenblok et al., 2019; Tenney et al., 2018). Second, although we have previously reported thalamic sources detected with MEG during absence seizures, the current study did not find significant thalamic CFC or within-frequency coupling (Tenney et al., 2014). While MEG can provide high spatiotemporal details about corticocortical oscillations, signal-to-noise is certainly reduced in deep subcortical regions. Previous neuroimaging studies have identified important components of thalamic, basal ganglia, and brainstem nuclei that likely participate in the generation and propagation of absence seizures and are less well studied with MEG (Bai et al., 2010; Carney et al., 2010; Masterton et al., 2013; Moeller et al., 2008; Salek-Haddadi et al., 2003). This is an important reason for continued multimodal neuroimaging and neurophysiology studies.
Conclusion
This study provides fMRI-informed MEG multilayer network analyses showing beta/gamma CFC and within-frequency coupling in the frontoparietal and frontofrontal regions during childhood absence seizures, which has not previously been reported for any primary generalized seizure. These results provide a possible mechanistic explanation for previous fMRI and neurophysiology studies showing that focal areas of the precuneus and frontal lobe are critically important for the initiation and maintenance of the generalized rhythmic spike wave discharges characterizing absence seizures. As nonpharmacologic treatments, such as neuromodulation, become available for generalized epilepsies, detailed mechanistic understanding of how “diffuse” seizures are generated and maintained will be necessary to provide optimal outcomes. We hope that these findings will encourage further studies that aim to understand the mechanisms of all generalized epilepsies.
Footnotes
Authors' Contributions
J.R.T.: Qualifies as an author based on (1) drafting/revising the article for content, (2) study concept and design, and (3) analysis and interpretation of data. B.J.W.: Qualifies as an author based on (1) revising the article for content and (2) analysis and interpretation of data. D.S.K.: Qualifies as an author based on (1) revising the article for content and (2) analysis and interpretation of data.
Author Disclosure Statement
All authors have no conflicts to disclose.
Funding Information
Portions of this study were funded by a “Taking Flight” award from the Citizens United for Research in Epilepsy (J.R.T.).