
Abstract
Introduction:
Infantile hydrocephalus (HCP) is a condition in which there is an abnormal buildup of cerebrospinal fluid in the ventricles within the first few months of life, which puts pressure on surrounding brain tissues. Compression of the developing brain increases the risk of secondary brain injury and cognitive disabilities.
Methods:
In this study, we used diffusion-weighted imaging and resting-state functional magnetic resonance imaging to investigate the effects of ventricle dilatation on structural and functional brain networks in children with shunted infantile HCP and examined how these brain changes may impact executive function.
Results:
We found that children with HCP have altered structural and functional connectivity between and within large-scale networks. Moreover, hyperconnectivity between the ventral attention and default mode network in children with HCP correlated with reduced executive function scores. Compared with typically developing age-matched control participants, our patient population also had lower fractional anisotropy in posterior white matter.
Discussion:
Overall, these findings suggest that infantile HCP has long-term effects on brain network connectivity, white matter development, and executive function in children at school age. Future work will examine the relationship between ventricular volumes before shunt placement in infancy and brain network development throughout childhood.
Impact statement
Infantile hydrocephalus (HCP) increases the risk of cognitive disability and secondary brain injury due to compression of the developing brain. In this study, we report that children with HCP show hyperconnectivity between the ventral attention and default mode network, which correlated with reduced executive function scores. HCP children also had lower fractional anisotropy in posterior white matter. This work highlights the long-term impact of infantile HCP on brain network connectivity and executive function in children at school age.
Introduction
Infantile hydrocephalus (HCP) is a rare condition in which there is an abnormal buildup of cerebrospinal fluid (CSF) in the ventricles of the brain within the first few months of life (Del Bigio, 1993; Erickson et al., 2001; Mataró et al., 2001). Epidemiological studies of infantile HCP have reported prevalence estimates that vary from 1 to 32 per 10,000 live births (Tully and Dobyns, 2014), with the most recent estimated prevalence of 1.1 per 1000 live births (Munch et al., 2012).
There are several causes of infantile HCP, including spina bifida, Dandy–Walker malformation, genetic abnormalities, complications of premature birth leading to bleeding in the brain (e.g., intraventricular hemorrhage), or an infection of the brain and spinal cord such as bacterial meningitis. The excess CSF increases the size of the ventricles (ventricle dilatation), which then increases pressure on surrounding brain tissue (Del Bigio et al., 2003).
A major difference between infantile HCP and HCP acquired later in life is the ratio of posterior to anterior ventricular horn dilatation; more specifically, that infantile HCP results in a greater expansion of the posterior horns (Kishimoto et al., 2018; Qiu et al., 2017). Studies using three-dimensional ultrasound in preterm infants with HCP, due to intraventricular hemorrhage, have shown that the posterior horns dilate more than the anterior horns and have suggested that there is a stronger effect on the posterior cortex (Kishimoto et al., 2018; Qiu et al., 2016, 2017). There are two major types of HCP: communicating HCP, which is due to impaired CSF reabsorption without obstructing CSF flow between the ventricles, and noncommunicating HCP, which is due to an obstruction blocking CSF flow between the ventricles.
In communicating HCP, the pressure within the ventricles is constant throughout since there is no obstruction; as such, the mechanism behind the increased posterior horn dilatation should be due to decreased resistance from surrounding posterior structures, possibly due to a relative lack of myelination of these structures compared with structures surrounding the anterior horns. In the case of noncommunicating HCP, greater dilatation occurs in areas proximal to the obstruction of CSF flow.
Greater expansion of the posterior horns puts the surrounding posterior cortex under prolonged compression and results in increased inflammation, cortical thinning, and increased CSF volumes (Dorner et al., 2018; Fletcher et al., 1992, 1996b; McAllister et al., 1991). Overall, this is consistent with indications that children with HCP have sustained greater damage to structures adjacent to the posterior horns compared with the anterior horns (Brouwer et al., 2010, 2016; Dorner et al., 2018).
Compression of the developing brain increases the risk of secondary brain injury, especially in parietal and occipital areas, and often results in cognitive and developmental disabilities (Del Bigio, 2010; Erickson et al., 2001; Mataró et al., 2001). Despite surgical treatment with ventriculoperitoneal (VP) shunt, infantile HCP remains an incurable condition with lasting cognitive and behavioral impairments that appear during infancy and early childhood in domains, such as memory, language, and motor skills (Behjati et al., 2011; Bigler, 1988; Mataró et al., 2001). However, deficits in higher order executive function behaviors typically present later in childhood and their relationship with disruptions in structural and functional brain networks in children with infantile HCP are not well understood (Dorner et al., 2018; Fletcher et al., 1992, 1996a; Hannay, 2000).
Neuroimaging techniques, such as diffusion-weighted imaging (DWI) and resting-state functional magnetic resonance imaging (fMRI), offer the ability to noninvasively examine in vivo structural or functional brain network connectivity. DWI is sensitive to the rate of water diffusion in biological tissue, which can then be modeled using tensor analysis to characterize white matter microstructure and estimate local fiber orientations for tractography and structural connectivity analyses (Basser et al., 1994a,b; Thomsen et al., 1987).
Several studies on HCP patients have used DWI to examine the effects of early ventricular dilatation and increased intracranial pressure on white matter microstructure and its relation with clinical deficits (Air et al., 2010; Assaf et al., 2006; Ben-Sira et al., 2015; Isaacs et al., 2019; Kulkarni et al., 2015; Rajagopal et al., 2013; Scheel et al., 2012; Tan et al., 2018; Williams et al., 2015; Yuan et al., 2009, 2013b, 2015).
Resting-state fMRI can be used to measure functional connectivity, which is the correlation between fluctuations in the blood-oxygen level-dependent (BOLD) signal in remote brain areas (Biswal et al., 1995). Since functional connectivity largely reflects white matter structural connectivity (Hagmann et al., 2008; Honey et al., 2009), functional networks in children with previous infantile HCP may be altered given the extensive structural damage sustained in the early stages of brain development.
Previous imaging research on infantile HCP has largely focused on early outcomes in young children up to 4 years old. Only a few imaging studies have investigated long-term outcomes of infantile HCP in children at school age (Mangano et al., 2016; Tan et al., 2018; Williams et al., 2015); however, these studies grouped patients with large differences in age, which may blur age-specific findings and have only examined changes in structural connectivity, but not functional connectivity. Since impairments in executive functions are better detected in children with HCP at school age (Burmeister et al., 2005; Fletcher et al., 1996a), it is necessary to assess HCP patients in this age group to improve prognosis.
Executive functions broadly include the ability to regulate emotions, plan, organize, monitor, inhibit, and initiate behaviors to solve complex tasks (Alvarez and Emory, 2006). Several higher order brain networks are involved in executive function, including the task-positive frontoparietal control, dorsal attention, and ventral attention networks (Dosenbach et al., 2008; Vossel et al., 2014); the task-negative default mode network (Andrews-Hanna, 2011; Fox et al., 2005); and the limbic network involved in motivation, affect, learning, and memory (Mega et al., 1997).
In the present study, we used DWI and resting-state fMRI to investigate the effects of ventricle dilatation on structural and functional brain networks and fractional anisotropy (FA) in anterior and posterior white matter in school-aged children with shunted infantile HCP compared with age-matched typically developing controls. We also examined the relationship between these imaging metrics and executive function. We hypothesized that infantile HCP alters the structural and functional connectivity of brain networks involved in executive function and results in greater damage to the posterior white matter.
Materials and Methods
Participants
We performed a retrospective study with all data obtained from an ongoing neuroimaging and behavioral study of children with previous infantile HCP. Patients were recruited from a follow-up pediatric neurosurgery clinic at the London Health Sciences Centre in London, Ontario. All patients had been treated with a VP shunt within the first year of life and were clinically stable at the time of study recruitment. Typically developing children were recruited as the control group through Western University's Cognitive Neuroscience Research Registry (OurBrainsCAN) and the Western Psychology's Child Developmental Participant Pool. Parents provided written informed consent for their child to participate in the study and children provided verbal assent. This study was approved by the Western University Health Sciences Research Ethics Board.
Our study sample included 6 children with shunted infantile HCP (1 girl, 5 boys) who were 6 to 10 years of age (mean [M] = 8.18, standard deviation [SD] = 1.61) and 20 typically developing children (8 girls, 12 boys) who were also between 6 and 10 years of age (M = 8.37, SD = 1.60). Demographic information of HCP patients is listed in Table 1. The study protocol was repeated in Patient 4A about 1 year later (356 days); this participant is included in our analyses as Patient 4A and 4A2.
Demographic Information of Hydrocephalus Patients Included in the Study
F, female; HCP, hydrocephalus; IVH, intraventricular hemorrhage; M, male; VP, ventriculoperitoneal; XLBW, extra-low birth weight.
Parental socioeconomic status (SES) was measured using the Hollingshead Four Factor Index of Social Status, which uses parental education, occupation, sex, and marital status to determine a composite SES score (Cirino et al., 2002; Hollingshead, 2011). An unpaired two-sample t-test comparing parental SES scores between patients (M = 45.33, SD = 12.73) and controls (M = 46.71, SD = 13.91) revealed no significant group differences (p = 0.41).
Behavioral assessments
Executive function behaviors were evaluated using the Behavior Rating Inventory of Executive Function, Second Edition (BRIEF2) parent-rating form (Gioia et al., 2015). The BRIEF2 is a revised version of the BRIEF, which is a well-validated and gold standard test of executive function behaviors outside the laboratory environment (Mahone et al., 2002a,b). BRIEF2 scores show high internal consistency, test–retest stability, internal validity, and concurrent validity (Hendrickson and McCrimmon, 2019).
Each assessment form includes 63 items related to 9 clinical scales: inhibit, self-monitor, shift, emotional control, initiate, working memory, plan/organize, task-monitor, and organization of materials. These clinical scales are combined into three composite indexes: Behavior Regulation Index (inhibit, self-monitor), Emotion Regulation Index (shift, emotional control), and the Cognitive Regulation Index (initiate, working memory, plan/organize, task-monitor, organization of materials).
The Global Executive Composite comprises all clinical scales into an overall executive function score. Raw scores are then converted into age- and sex-standardized t-scores (mean t-score = 50) in which higher scores indicate lower executive function scores. The BRIEF2 scoring manual interprets t-scores between 60 and 64 as “mildly elevated,” scores between 65 and 69 as “potentially clinically elevated,” and scores above 70 as “clinically elevated.” Statistical differences in BRIEF2 composite scores between patients and controls were determined using unpaired two-sample t-tests (p < 0.05).
Image acquisition
In a training session before image acquisition, participants were exposed to a 10- to 15-min mock scan that simulated the space and sounds of a magnetic resonance imaging (MRI) scanner to reduce potential anxiety or fear during the real scan. MRI data were acquired in a second session on a 3-Tesla Siemens MAGNETOM Prisma Fit 60-cm horizontal bore scanner using a 32-channel head coil at the Centre for Functional and Metabolic Mapping in Robarts Research Institute.
Participants watched cartoon video clips during image acquisition to reduce motion artifacts—a standard practice in pediatric neuroimaging (Greene et al., 2018; Vanderwal et al., 2015). A high-resolution T1-weighted magnetization prepared-rapid gradient echo anatomical image was obtained with the following parameters: time to repetition (TR) = 2300 ms, time to echo (TE) = 2.93 ms, inversion time = 900 ms, flip angle = 9°, field of view = 256 × 256 mm, matrix size = 256 × 256, voxel size = 1.0 mm isotropic, number of slices = 160.
DWI data were obtained using a multi-band echo planar imaging (EPI) sequence with the following parameters: TR = 2500 ms, TE = 77.4 ms, field of view = 192 × 192 mm, matrix size = 96 × 96, number of averages = 4, voxel size = 2.0 mm isotropic, number of slices = 64, number of averages = 4. We acquired 30 diffusion-encoding gradient directions (b-value of 1000 sec/mm2) and 1 volume without diffusion-weighting (b-value = 0 sec/mm2). Functional images were collected using a multi-band EPI sequence at an acceleration factor of 4 with the following parameters: TR = 1000 ms, TE = 30 ms, flip angle = 40°, field of view = 208 × 208 mm, matrix size = 84 × 84, voxel size = 2.5 mm isotropic, number of slices = 48. Each functional run was 5 min in length and consisted of 300 EPI volumes.
Image preprocessing
Raw images were converted from DICOM to NIfTI format and organized according to the standard Brain Imaging Data Structure (BIDS) (Gorgolewski et al., 2016) using the “dcm2bids” package (Bedetti et al., 2019). All preprocessing steps were then carried out using the FMRIB Software Library (FSL) version 6.0 (Woolrich et al., 2009). DWI data were preprocessed using FMRIBs Diffusion Toolbox (FDT) implemented with FSL. Subject motion and eddy current-induced distortion were corrected using FSLs “eddy” tool (Andersson and Sotiropoulos, 2016). Diffusion tensor models were then fit at each voxel using “DTIFIT” to obtain scalar maps of the three major eigenvalues (λ1, λ2, λ3), which correspond to the magnitude of diffusion for each eigenvector of each fitted tensor.
Diffusion tensor-based FA scalar maps include voxel values between 0 and 1 representing the degree of anisotropic diffusion and are calculated as the relative difference of the first eigenvalue compared with the other two eigenvalues (Alexander et al., 2007; Basser and Pierpaoli, 1996; Basser et al., 1994a; Beaulieu, 2002). Finally, FSLs “BEDPOSTX” was used to account for crossing fibers by fitting a multiple tensor model at each voxel and generating more accurate probability distributions of voxel-wise fiber orientations (Behrens et al., 2003, 2007).
Functional data were preprocessed using FSLs fMRI Expert Analysis Tool which included brain extraction (Smith, 2002), MCFLIRT rigid-body motion correction (Jenkinson et al., 2002), and high-pass temporal filtering (cutoff at 100 sec). Next, nuisance covariates were regressed out from the BOLD signal, including six motion parameters and the average time series from white matter and CSF masks (which included the shunt). Spatial smoothing was then applied using a 5 mm full width at half maximum Gaussian kernel.
Regional FA analysis
We investigated FA differences in anterior and posterior white matter between patients and controls using a region of interest (ROI)-based approach. First, the diffusion tensor-based scalar FA map for each participant was thresholded to maintain values above 0.2 to exclude gray matter or CSF voxels (Smith et al., 2013). Next, white matter ROIs were defined using the Johns Hopkins University (JHU) white matter atlas (Hua et al., 2008; Wakana et al., 2007) after it was transformed from adult to pediatric template space using registration procedures described in the “Network Construction” section below. Anterior white matter ROIs were defined as the JHU parcels located anterior to the midline of corpus callosum and included the genu of corpus callosum, anterior corona radiata, and the anterior limb of the internal capsule (Fig. 1).

Anterior and posterior white matter regions of interest. Regions of interest defined from the JHU white matter atlas. CC, corpus callosum; FA, fractional anisotropy; JHU, Johns Hopkins University; L, left hemisphere; R, right hemisphere; ROI, region of interest; WM, white matter.
Posterior white matter ROIs were defined as the JHU parcels located posterior to the midline of corpus callosum and included the splenium of corpus callosum, posterior thalamic radiation, and the tapetum of corpus callosum (Fig. 1). FA values were then obtained by masking the thresholded FA maps with each white matter ROI. The mean FA value was then calculated for both anterior and posterior white matter by averaging across the three anterior or posterior ROIs. Statistical differences in anterior and posterior white matter FA between patients and controls were determined using the Wilcoxon rank sum test (p < 0.05).
Network construction
Cortical network nodes were defined in native space using the Schaefer2018 100 parcel parcellation scheme (Schaefer et al., 2018), with each parcel belonging to a large-scale cortical network, including the dorsal attention, ventral attention, limbic, frontoparietal control, and default mode networks (Yeo et al., 2011).
Network ROI labels and coordinates of the Schaefer2018 100 parcel parcellation in standard adult MNI152 template space are listed in Table 2. First, we transformed the Schaefer2018 100 parcel parcellation from the standard adult MNI152 template to age-appropriate pediatric templates, specifically one template for participants aged 4.5–8.5 years old and another for those aged 7.5–13.5 years old (Fonov et al., 2009, 2011). The Schaefer2018 parcellation in standard pediatric space was then transformed into native functional or diffusion space through a five-step process.
List of the Network Node Labels and Coordinates in Standard MNI152 Space
DMN, default mode network; DOR, dorsal attention network; FPC, frontoparietal control network; Hemi, hemisphere; L, left; LIM, limbic network; MNI, Montreal Neurological Institute; R, right; VENT, ventral attention network.
Each participant's functional or diffusion EPI images were linearly registered to native structural T1 space. Next, structural T1 images in native space were transformed to age-appropriate standard pediatric space using FSLs linear and nonlinear registration tools (Andersson et al., 2007; Jenkinson and Smith, 2001; Jenkinson et al., 2002). We then combined the two transformation matrices from (1) native functional/diffusion space to native structural T1 space and from (2) native structural T1 space to standard pediatric space. The output transformation matrix was then inversed and applied to the Schaefer2018 parcellation in standard pediatric space to transform it into native functional and diffusion space for each participant. Visual inspection confirmed that the parcellation scheme was successfully registered to native space.
Diffusion-weighted probabilistic tractography
Probabilistic tractography was implemented using FDTs “probtrackx2” command line tool to obtain diffusion-based measures of structural connectivity between all cortical ROIs (Behrens et al., 2003, 2007). For the HCP population, shunts were manually traced from structural T1-weighted images in FSLeyes and confirmed by a pediatric neurosurgeon to create termination masks. Shunt masks were included in the tractography analysis to terminate streamlines once they reached the mask to avoid having streamlines confounded by the directional diffusion of water along shunts. Standard tractography parameters (curvature threshold = 0.2, step length = 0.5, steps = 2000, subsidiary fiber volume threshold = 0.01) were applied to sample 5000 streamlines per voxel within each ROI mask (i.e., for n voxels in an ROI, 5000*n streamlines were sampled).
A “successful” streamline was one that was generated from a seed ROI voxel and reached a target ROI voxel; all cortical ROIs were used as seed–target pairs. This algorithm generated a matrix where each cell contained the total number of successful streamlines from seed ROI A to target ROI B. We normalized the streamline counts for each seed–target pair by dividing the total number of successful streamlines by the total number of streamlines sampled from the seed ROI. Since diffusion-weighted tractography is dependent on the seed ROI location, the probabilistic streamline count from seed A to target B is slightly different from that from seed B to target A. Thus, we averaged the probabilistic streamline counts from A to B and from B to A to obtain an undirected (symmetric) streamline probability matrix.
Next, we applied a common logarithm transformation to improve the normality of the data because streamline probabilities are skewed toward smaller values and vary across multiple orders of magnitude. Log-transformed streamline probabilities between each pair of ROIs were used as a measure of structural connectivity. More specifically, streamline probabilities between ROIs within the same brain network were classified as a measure of within-network structural connectivity, and streamline probabilities between ROIs from different networks were used as a measure of between-network structural connectivity.
Statistical differences in structural connectivity (log-transformed) between patients and controls were determined using unpaired two-sample t-tests with false discovery rate (FDR) correction (p < 0.05) for multiple comparisons using the Benjamini–Hochberg procedure (Benjamini and Hochberg, 1995).
Resting-state functional connectivity analysis
The BOLD signal time series was averaged across all voxels within each parcel ROI, and the pairwise linear correlation coefficient was calculated between the BOLD time series of each pair of ROIs. Fisher's r to z transformation was then applied to normalize correlation coefficients into z-scores, where z-scores denote the functional connectivity between each pair of ROIs. A z-score between ROIs from the same brain network was classified as a measure of within-network functional connectivity, and z-scores between ROIs from different networks were used as a measure of between-network functional connectivity. All z-scores in the averaged functional connectivity matrices for patients and controls were positive.
Statistical differences in functional connectivity (|z-scores|) between patients and controls were determined using unpaired two-sample t-tests with FDR correction (p < 0.05) for multiple comparisons using the Benjamini–Hochberg procedure (Benjamini and Hochberg, 1995).
Correlation between executive function and imaging metrics
We investigated whether executive function correlated with anterior and posterior FA, structural connectivity, and functional connectivity. To minimize the number of correlation tests, we selected only the Global Executive Composite score from the BRIEF2 assessment as the measure of executive function. A linear correlation coefficient was calculated between the Global Executive Composite score and (1) anterior and posterior white matter FA, (2) within-network and between-network structural connectivity, and (3) functional connectivity of the five networks involved in executive function (dorsal attention, ventral attention, limbic, frontoparietal, and default mode networks).
Results
Executive function behaviors
Parental reports on the BRIEF2 assessment revealed differences between HCP patients and healthy controls on executive function behaviors (Fig. 2). An unpaired two-sample t-test comparing age- and sex-standardized mean t-scores between patients and controls revealed a statistically significant difference on the Global Executive Composite (p = 0.024) with a large effect size (Hedges' g = 1.14), with patients showing mildly elevated to potentially clinically elevated t-scores. No significant differences between patients and controls were found on the Behavior Regulation Index (p = 0.078), the Emotion Regulation Index (p = 0.079), or the Cognitive Regulation Index (p = 0.097).

Composite scores on the BRIEF2 assessment of executive function. Age- and sex-standardized mean t-scores are shown for healthy controls and HCP patients. Error bars represent the standard deviation. t-Scores near 50 are considered average. t-Scores between 60 and 64 are 1–1.5 standard deviations above the mean and are considered “mildly elevated.” t-Scores between 65 and 69 are 1.5–2 standard deviations above the mean and are considered “potentially clinically elevated.” t-Scores above 70 are more than 2 standard deviations above the mean and are considered “clinically elevated.” Statistical differences between patients and controls were determined using unpaired two-sample t-tests (p < 0.05). Statistically significant differences are indicated by an asterisk (*). Patients and controls only differed significantly on the Global Executive Composite score, with patient scores ranging from mildly elevated to potentially clinically elevated. BRIEF2, Behavior Rating Inventory of Executive Functions, Second Edition; HCP, hydrocephalus.
FA of anterior and posterior white matter
FA values in anterior and posterior white matter were statistically compared between HCP patients and controls and were included in a correlation analysis with Global Executive Composite scores. In HCP patients, FA was significantly higher in anterior white matter (p = 0.018) with a medium effect size (Hedges' g = 0.43) and significantly lower in posterior white matter (p < 0.001) with a large effect size (Hedges' g = 1.81) compared with controls (Fig. 3). There were no significant correlations between the Global Executive Composite and FA in anterior or posterior white matter.
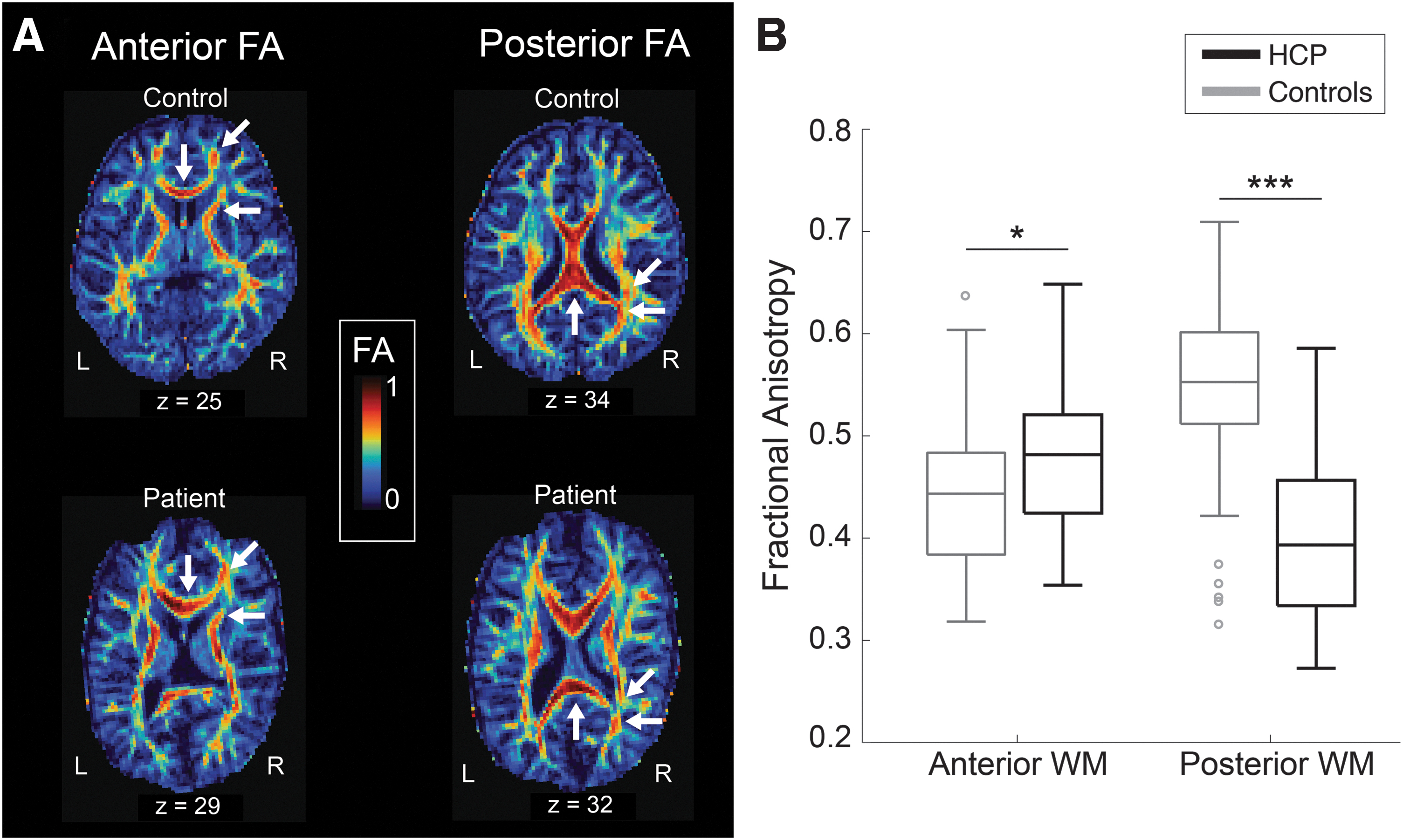
FA of anterior and posterior white matter.
Structural connectivity of large-scale cortical networks
DWI data were used to perform probabilistic tractography between all cortical ROIs assigned to one of five brain networks involved in executive function behaviors: dorsal attention, ventral attention, limbic, frontoparietal control, or default mode network. A structural connectivity matrix composed of log-transformed probabilistic streamline counts was generated for each group. Statistically significant differences in structural connectivity between patients and controls are illustrated in Figure 4, and the numerical data are listed in Supplementary Table S1. Effect sizes for all significant structural connectivity differences were measured using Hedges' g, and each had at least an effect size of g = 1.2, which is considered a large effect.

Differences in structural connectivity between HCP patients and controls.
We found that HCP patients had weaker within-network structural connectivity in the right dorsal attention network (Fig. 4). Patients also had weaker interhemispheric between-network structural connectivity between the right frontoparietal network ROIs and the left dorsal attention, left ventral attention, left limbic, and left default mode network ROIs. Reduced structural connectivity was also found between the left frontoparietal network and the right dorsal attention and left ventral attention networks. Finally, HCP patients also had weaker between-network structural connectivity between the left dorsal attention network ROIs and the right ventral attention and left default mode network ROIs.
Increased interhemispheric between-network structural connectivity was found between the dorsal attention and default mode networks in patients compared with controls (Fig. 4). We also found evidence of increased between-network structural connectivity between the left frontoparietal (precuneus) and right default mode networks (ventral prefrontal cortex) and between the left limbic and left dorsal attention network.
Functional connectivity of large-scale cortical networks
Resting-state fMRI data were used to obtain pairwise absolute functional connectivity values (|z-scores|) between all cortical ROIs assigned to one of five brain networks involved in executive function behaviors: dorsal attention, ventral attention, limbic, frontoparietal control, or default mode network. A functional connectivity matrix was generated for each group, and the statistically significant differences between patients and controls are illustrated in Figure 5, and the numerical data are listed in Supplementary Table S2. Effect sizes for all significant functional connectivity differences were measured using Hedges' g, and each had at least an effect size of g = 0.8, which is considered a medium effect.
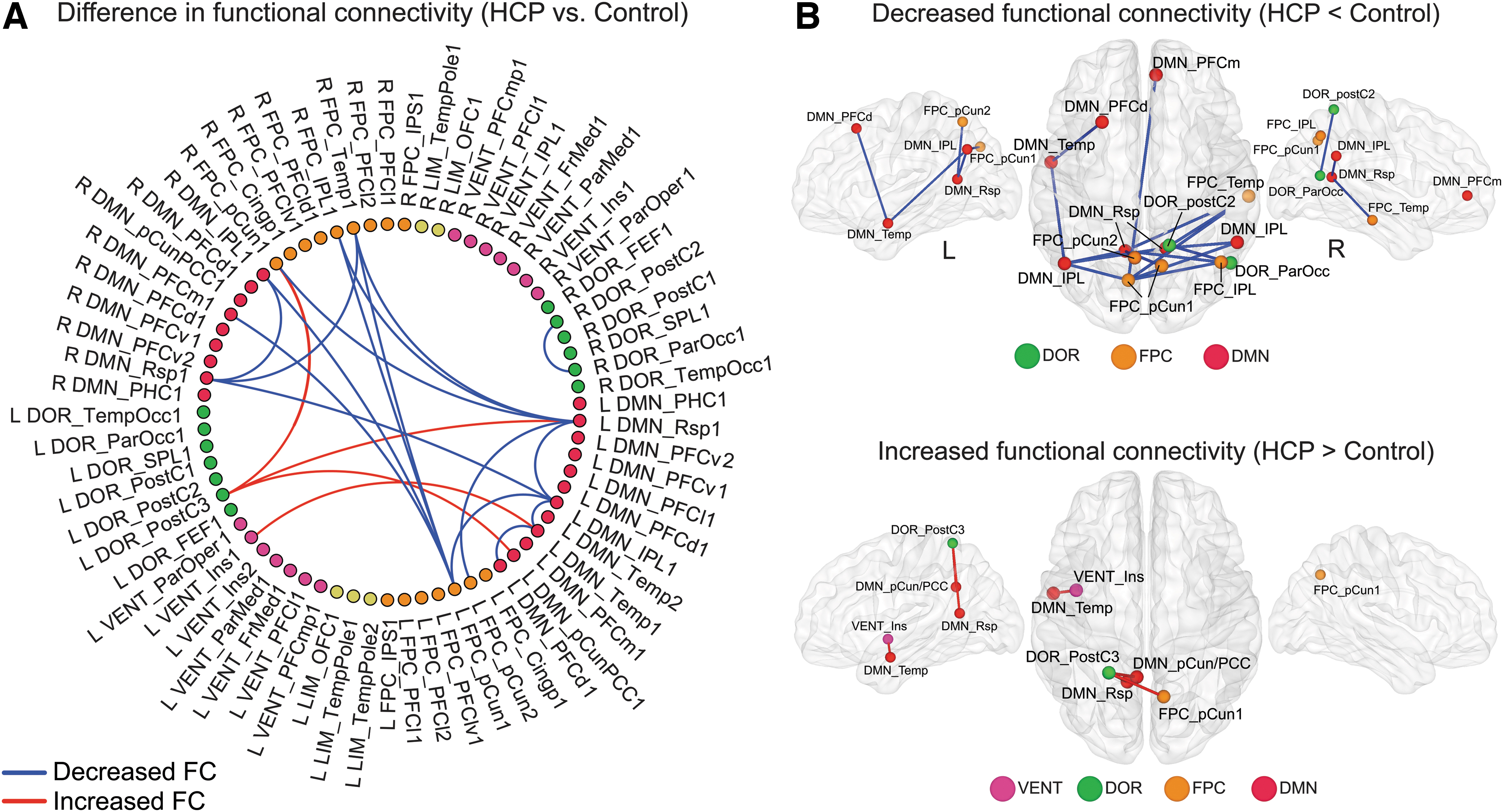
Differences in functional connectivity between HCP patients and controls.
Compared with typically developing controls, HCP patients showed reduced within-network functional connectivity in the dorsal attention (between right hemisphere ROIs), default mode (between left hemisphere ROIs and between right and left hemisphere ROIs), and frontoparietal network (between right and left hemisphere ROIs). HCP patients had reduced between-network functional connectivity between the bilateral frontoparietal network and default mode network ROIs compared with controls (Fig. 5). Stronger between-network functional connectivity in HCP patients was found between the left dorsal attention network ROIs and the right frontoparietal and left default mode network ROIs as well as between the left ventral attention network and the left default mode network (Fig. 5).
Correlation between executive function and brain network connectivity
A linear correlation analysis was performed between the BRIEF2 Global Executive Composite scores and the within- and between-network functional and structural connectivity of the five resting-state networks involved in executive function: dorsal attention, ventral attention, frontoparietal control, default mode, and limbic networks. We found a significant correlation in HCP patients between the Global Executive Composite and the between-network functional connectivity between the ventral attention–default mode networks (r = 0.88, p = 0.019), but not in the control group (r = 0.26, p = 0.326; Fig. 6). There were no significant correlations between the Global Executive Composite and the within- or between-network structural connectivity values in either the HCP group or the control group.

Correlation between the Global Executive Composite and functional connectivity. Higher scores on the Global Executive Composite indicate reduced executive function behaviors. Functional connectivity between the ventral attention and default mode network significantly correlated with scores on the Global Executive Composite in HCP patients, but not controls. HCP patients with lower executive function scores showed greater functional connectivity between these cortical networks. p, probability value; r, correlation coefficient.
Discussion
The present study identified several associations between infantile HCP and subsequent brain network connectivity and executive function at school age. We found that children with infantile HCP show deficits of global executive function compared with typically developing children, which correlated with between-network functional connectivity between the ventral attention and default mode networks. Children with infantile HCP had decreased between-network structural connectivity between the ventral attention network and frontoparietal and dorsal attention networks, between the limbic and frontoparietal network, and decreased within-network structural and functional connectivity within the right dorsal attention network.
In contrast, increased between-network structural connectivity was found between the limbic and dorsal attention network in HCP patients. Mixed between-network connectivity was found between the default mode network and frontoparietal and dorsal attention network ROIs and between the frontoparietal and dorsal attention network ROIs. Finally, children with infantile HCP showed increased FA values in anterior white matter and decreased in posterior white matter compared with typically developing children. Findings suggest that infantile HCP has long-lasting effects on children's brain network and white matter development and executive function.
Parental reports of impaired executive function on the BRIEF2
The Global Executive Composite score provides an overall measure of executive function by accounting for several clinical scales and is often the sole measure used as an index of children's executive function (Hawkey et al., 2018; King et al., 2020). In our study, children with HCP had higher scores (i.e., lower performance) on the Global Executive Composite compared with typically developing children without HCP; however, patient scores were only mildly to potentially clinically elevated and did not reach the clinically elevated range.
Reports on executive function in this patient population have been varied across studies (Erickson et al., 2001), with some finding little to no differences between patients and controls (Fletcher et al., 1996a; Holwerda et al., 2016) and others finding impaired executive function in children with shunted HCP and normal intellectual quotient on scales of planning, problem solving, working memory, mental flexibility, and self-monitoring (Fletcher et al., 1996b; Lacy et al., 2012).
Moreover, Lacy and colleagues (2012) examined children aged 6–16 years and found that problems with executive function tended to increase with age. Thus, discrepancies in the literature regarding executive functions may be related to within-group age differences. Future work with larger sample sizes will be useful to determine which domain of executive function (behavior, emotion, or cognition) from the BRIEF2 accounts for the elevated Global Executive Composite score. In addition, since performance-based measures and parental ratings of executive function have been shown to assess different constructs (Ten Eycke and Dewey, 2016; Toplak et al., 2013; Vriezen and Pigott, 2002), future studies may include both assessment types for a more comprehensive measure of executive function in this population.
Altered FA in anterior and posterior white matter
Compared with a healthy control population, children with HCP had lower FA in posterior white matter ROIs, which included the splenium and tapetum of the corpus callosum and the posterior thalamic radiation, and higher FA in anterior white matter ROIs, which included the genu of the corpus callosum, anterior corona radiata, and the anterior limb of the internal capsule. This finding supports our hypothesis that the posterior brain suffers greater damage in children with HCP due to greater expansion of posterior horns of the lateral ventricles or enlargement of the fourth ventricle. Reduced FA is thought to reflect demyelination, decreased fiber density, and/or axonal loss (Beaulieu, 2002; Sotak, 2002), which may be due to increased pressure on periventricular white matter (Assaf et al., 2006; Yuan et al., 2013a).
In a recent study, Tan and colleagues (2018) investigated children and adults with HCP (age range: 9–39 years) who received shunt surgery within the first 2 years of life and similarly found lower FA in posterior white matter structures. However, studies that used diffusion tensor analysis in infants with HCP pre-shunt found lower FA in the genu of the corpus callosum (Air et al., 2010; Assaf et al., 2006; Mangano et al., 2016; Yuan et al., 2009), which normalized after 1–2 years following shunt surgery in one study (Air et al., 2010) but remained low in others (Assaf et al., 2006; Mangano et al., 2016). In contrast with these studies in infants with HCP, Tan and colleagues (2018) found no difference in FA values of the genu of corpus callosum (anterior white matter) in their older HCP patient population compared with healthy controls.
Since our patient group is more than 5 years post-shunt, one possible explanation for our finding of higher FA in the anterior white matter (including the genu) is that it reflects long-term compensatory remodeling of white matter fibers (e.g., increased myelination) after sustaining damage from ventricular enlargement (Beaulieu, 2002; Del Bigio, 2010). However, given that our patient group is younger than patients in the study by Tan and colleagues, the increased FA that we found may normalize and return to baseline in later years as seen in the study by Tan and colleagues (2018).
Altered structural and functional brain network connectivity
We found evidence of both hypo- and hyperconnectivity within and between brain networks in children with HCP. Within-network hypoconnectivity was found within the dorsal attention, default mode, and frontoparietal control networks. Specifically, hypoconnectivity in the dorsal attention network involved the superior parietal lobule (SPL) node, and hypoconnectivity within the default mode network involved the dorsal prefrontal cortex (PFCd) node. Both areas play a role in higher cognitive functions, with the SPL involved in the top-down control of spatial attention and visuospatial working memory (Giesbrecht et al., 2003; Klingberg et al., 2002; Shomstein et al., 2010; Yantis et al., 2002) and the PFCd involved in working memory and inhibitory control (Arnsten, 2011; Corbetta et al., 2002; Goldman-Rakic, 1996).
Imaging studies in children with HCP are scarce, which limits our ability to contextualize these specific findings with the existing literature. However, one such study by Yuan and colleagues (2016) applied a graph theory analysis to post-shunt DTI data from HCP patients aged 1–18 years and found abnormalities in network degree and betweenness centrality in the SPL and PFCd (specifically in the superior frontal gyrus and middle frontal gyrus). Although degree and betweenness centrality are not directly comparable to the strength of connectivity, this study supports our findings that the SPL and PFCd are altered in children with HCP.
Between-network hypoconnectivity was found between the ventral attention network and frontoparietal control and dorsal attention networks and between the limbic network and frontoparietal control network. This hypoconnectivity between the ventral attention and frontoparietal control networks included the medial frontal gyrus and precuneus/posterior cingulate cortex (pCun/PCC) nodes, which have also shown abnormal network degree and betweenness centrality in previous DTI studies in children with HCP (Yuan et al., 2015, 2016).
The medial frontal gyrus is important for response inhibition (Floden and Stuss, 2006; Verfaellie and Heilman, 1987), and the pCun/PCC plays a role in cognition, self-awareness, emotion, memory, and visuospatial functions (Cavanna and Trimble, 2006; Leech and Sharp, 2014; Maddock et al., 2003; Wallentin et al., 2006). These outlined visuospatial and cognitive functions carried out by brain areas with lower structural or functional connectivity are most often impaired in the HCP population (Erickson et al., 2001; Mataró et al., 2001). It is possible that these brain areas sustain secondary damage due to ventricular enlargement and increased intracranial pressure, which then disrupts the normal functioning carried out by these brain regions.
Conversely, between-network hyperconnectivity was found between the limbic (temporal pole) and dorsal attention network and between the ventral attention and default mode network in children with HCP. The temporal pole is part of an extended limbic network and plays a role in social and emotional processing (Olson et al., 2007). As for the hyperconnectivity between the ventral attention and default mode network, one of the areas involved is the insula which has also shown abnormal degree and betweenness centrality in a previous DTI study in children with HCP (Yuan et al., 2015) and plays a role in a variety of functions including sensory and affective processing (Uddin et al., 2017).
Although our patient sample was not significantly different from controls on the BRIEF2 Emotional Regulation Index, Iglesias and colleagues (2020) reported that social–emotional scores on the Hydrocephalus Outcome Questionnaire were lower in older children with shunted HCP. Thus, it is possible that this abnormal network hyperconnectivity may precede the onset of social–emotional impairments (Khundrakpam et al., 2016).
Imaging studies in infants with post-hemorrhagic HCP and intraventricular hemorrhage have also reported altered structural and functional network connectivity, with some patients showing increased connectivity before and after VP shunt insertion (Smyser et al., 2013, 2019). Hyperconnectivity in HCP patients may reflect brain reorganization following injury due to ventricular dilatation as a compensatory response or may be maladaptive and lead to future neurodegeneration due to increased brain metabolism from chronic hyperconnectivity (Hillary and Grafman, 2017).
Interestingly, we found that higher between-network functional connectivity between the ventral attention and default mode network correlated with lower scores of executive function in children with HCP. The ventral attention network is involved in reorienting attention toward salient stimuli (i.e., bottom-up processing) and is considered a task-positive network since it becomes activated during a task (Corbetta and Shulman, 2002; Corbetta et al., 2008; Fox et al., 2005). However, the default mode network is involved in self-monitoring, introspection, and mind wandering and is considered a task-negative network since it becomes active in the absence of a task and is anticorrelated with task-positive networks (Fox et al., 2005; Raichle, 2015).
Consistent with our findings, previous studies have shown that behavioral performance on high-level cognitive tasks is reduced with higher positive functional connectivity between task-positive and task-negative networks (DeSerisy et al., 2021; Keller et al., 2015; Kelly et al., 2008).
Finally, we found evidence of both hypo- and hyperconnectivity between areas of the (1) default mode and frontoparietal network, (2) default mode and dorsal attention network, and (3) frontoparietal and dorsal attention network. These inconsistent connectivity findings may reflect the evolving interactions between large-scale networks that continue to develop throughout childhood (Betzel et al., 2014; Fair et al., 2007; Jolles et al., 2011; Marek et al., 2015; Zhong et al., 2014). Alternatively, some areas of hyperconnectivity between networks may be a compensatory response to the decreased connectivity between areas of the same networks due to white matter damage in HCP.
Caveats and limitations
Indeed, the small patient sample size limits the generalizability of our findings to those individual children included in this study. However, small sample sizes are common in clinical pediatric research due to challenges in recruitment and training of pediatric populations, especially for imaging studies. In addition, infantile HCP is a relatively rare condition, which further limits patient recruitment. Nonetheless, this combined imaging and behavioral study is an important step to elucidating the effects of infantile HCP on children's brain networks and functioning.
It is worth mentioning that although patient 4A was included twice in this study sample 1 year later as patient 4A2, our findings remained largely unchanged when we ran analyses without two instances of patient 4A. When the structural connectivity analysis included only one instance of patient 4A, significance was lost for the increased between-network structural connectivity between limbic and dorsal attention networks and for the decreased between-network connectivity between frontoparietal and dorsal attention networks.
As for functional connectivity, when the analysis included only one instance of patient 4A, significance was lost for the increased between-network functional connectivity between the right frontoparietal and left dorsal attention networks. The rest of the changes in structural and functional connectivity as well as all other major findings remained significant and unchanged.
Finally, it will be important for future studies to use direct measures of executive function when assessing neurodevelopmental outcomes of HCP patients. Since performance-based measures and parental ratings of executive function have been shown to assess different constructs (Ten Eycke and Dewey, 2016; Toplak et al., 2013; Vriezen and Pigott, 2002), future studies may include both assessment types for a more comprehensive measure of executive function in this population.
Conclusions
This study reveals that shunted infantile HCP has long-term effects on school-aged children's structural and functional brain networks and executive functioning. Our findings show that children with HCP have abnormal brain connectivity between and within large-scale networks that may be a result of the sustained damage from ventricular enlargement and increased intracranial pressure. Moreover, between-network hyperconnectivity between the ventral attention and default mode network was correlated with lower scores of executive function.
In addition, our diffusion tensor analysis revealed that children with HCP sustain greater damage to posterior white matter than anterior white matter and have lower FA in posterior white matter compared with typically developing controls. Overall, this work highlights a need to continue investigating the long-term outcomes of children with previous infantile HCP as they mature into later childhood and adolescence. Future studies will examine the relationship between ventricle volumes before shunt placement in infancy and the later brain network connectivity at school age.
Footnotes
Acknowledgments
We thank Estelle Ansermet and Ikhlas Hashi for assisting with data collection.
Authors' Contributions
R.A.: Methodology, software, formal analysis, writing—original draft, visualization. D.G.: Methodology, investigation, recruitment, data acquisition. J.B.M.: Methodology, supervision, writing—review and editing. R.E.: Methodology, conceptualization, supervision, writing—review and editing. S.d.R.: Conceptualization, methodology, writing—review and editing, supervision, funding acquisition.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Canada First Research Excellence Fund (CFREF) BrainsCAN Supported MRI Rates and the Western Strategic Support for CIHR Success Grant.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.