
Abstract
Background:
The role of the cerebellum in motor function is well recognized. However, its role in higher nervous system activities such as cognition, emotion, endocrine, and autonomic activities is less known. The present study aims to show direct dento-amygdala projections using a biotinylated dextran amine (BDA) tracer in rats and 3-tesla (T) high-resolution diffusion tensor imaging (DTI)-based tractography in humans.
Materials and Methods:
The BDA tracer was pressure injected into the dentate nucleus of the cerebellum of Wistar albino rats. Labeled cells and axons were documented. High-resolution 3-T tractography data were obtained from the Human Connectome Project database. Dento-amygdala tracts were analyzed using diffusion spectrum imaging (DSI) Studio software.
Results:
The experimental study showed bilateral projections between the dentate nucleus and the central and basal nuclei and ipsilateral projections between lateral nuclei of the amygdala. The fibers from the dentate nucleus reached the amygdala through the superior cerebellar peduncle (SCP), and the contralateral fibers crossed in the decussation of SCP at the midbrain. The dento-amygdala results of the experimental study corresponded with the 3-T tractography findings on humans. Additionally, DTI findings showed that most of the dentate fibers passed through the hypothalamus before reaching the amygdala, and the amygdalae of the two sides are connected through the anterior commissure.
Discussion:
The 3-T DTI data of adult humans showed both direct dento-amygdala and indirect dento-hypothalamo-amygdala projections. Thus, this may indicate cerebellar contribution in modulation of emotional and autonomic functions. Furthermore, this can explain the emotional and cognitive deficits that occur in patients with cerebellar or SCP damage.
Impact statement
The present study showed direct dento-amygdala connections in the rat brain and human brain, which may provide evidence for cerebellar contribution in modulation of emotional and autonomic functions.
Introduction
The cerebellum forms ∼10% of the total intracranial volume and contains more than half of the total number of neurons in the human central nervous system (CNS) (Kandel et al., 2000). Traditionally, the cerebellum has a primary role in a wide variety of motor behaviors such as motor control, balance, posture, eye movements, control of volitional goal-directed movements, and motor learning (Middleton and Strick, 1998; Stoodley and Schmahmann, 2010).
Recent functional neuroimaging, experimental, electrophysiological, and clinical studies with advanced technical devices have increased our understanding regarding the complex nonmotor functions of the cerebellum (Çavdar et al., 2020; Middleton and Strick, 1997, 1998). The results of these researches demonstrated the role of the cerebellum in higher nervous system activities such as cognition, emotion, memory, endocrine, autonomic, and even immune functions (Adamaszek et al., 2017; Peng et al., 2006).
The direct and indirect connections of the cerebellum with limbic structures (hypothalamus, hippocampus, anterior thalamus, and amygdala) have been documented in literature (Bohne et al., 2019; Konarski et al., 2005). Our former and many other studies have described the presence of cerebello-hypothalamic connections in rats, cats, and human adults (Kamali et al., 2018; Konarski et al., 2005).
Moreover, electrophysiological and tract-tracing studies showed the presence of reciprocal cerebello-hippocampal connections (Liu et al., 2012; Snider and Maiti, 1976). Furthermore, the cerebellum is connected to brainstem areas (ventral tegmental area, periaqueductal gray, and locus coeruleus) that are connected with limbic and paralimbic regions (Heath and Harper, 1974).
The amygdala is the part of the brain responsible for coordination of behavioral and emotional responses (Sergerie et al., 2008). The amygdala interacts extensively with cortical–subcortical–limbic circuits. The anatomical location of the amygdala and its multiple connections with large areas of the CNS show that it is an integrating center for emotional and behavioral processing. The functional interactions between the amygdala and other brain regions are being widely studied to understand their involvement in amygdala-based neuropsychiatric disorders.
In recent years, evidence of the role of the cerebellum in emotional processing has increased substantially (Adamaszek et al., 2017; D'Angelo and Casali, 2013). Clinical studies have shown that lesions of the cerebellum result in neurological and neuropsychiatric disorders, particularly in schizophrenia (Phillips et al., 2015; Shakiba, 2014), autism (Fatemi et al., 2012), and depression (Schmahmann, 2004). The data from neuroimaging and clinical studies identify the cerebellum as a key region among the emotion-relevant structures (Baumann and Mattingley, 2012; Stoodley and Schmahmann, 2010).
The earliest acknowledged cerebellar involvement in emotion and emotional disorders was in the late 1970s (Heath et al., 1978; Snider and Maiti, 1976). Early studies showed that electrical stimulation of the cerebellum produced evoked responses in the amygdala (Heath and Harper, 1974). However, a clear demonstration of cerebello-amygdala connections was not described in the literature. In our previous studies, we have clearly defined the cerebello-hypothalamic connections in rats and human adults using tract tracing and fiber dissection and diffusion tensor imaging (DTI) techniques (Çavdar et al., 2020).
In the present study, we aimed to expose the dento-amygdala projections in the rat brain and human brain. The connections of the cerebellum with limbic structures can be clinically important and can further enrich our understanding of cerebellar contribution to limbic circuits. The present study was designed in two parts; in the experimental part, we aimed to inject the biotinylated dextran amine (BDA) tracer into the dentate nucleus of the cerebellum of the rat brain and document the dento-amygdala projections. The second part of the study using 3-tesla (T) DTI tractography reveals dento-amygdala pathways in humans.
Materials and Methods
Animal experiment
Adult Wistar albino rats weighing 250–400 g were fed standard laboratory rat chow and tap water ad libitum and housed in plexiglass cages with a 12-h light–12-h dark cycle in a temperature-controlled room (20°C ± 3°C). The Institutional Animal Care and Use Committee approved all procedures. A total of 11 animals were used for the BDA injections into the dentate nucleus of the cerebellum.
Animals were deeply anesthetized with pentobarbital (Narcoren) administered as 35 mg/kg body weight (BW) together with a mixture (1 mL/kg BW) of ketamine (25 mg/mL), xylazine (1.3 mg/mL), and acepromazine (0.25 mg/mL) intraperitoneally (ip). The heads of the animals were placed in a stereotaxic frame (Stoelting Model 51600 [Fisher Scientific], Wood Dale, IL). The scalp was incised longitudinally, and the skull was exposed posterior to lambda. A small hole was drilled in the skull at a position appropriate for unilateral injection of the tracer into the dentate nucleus.
Injections were made at the following coordinates of the dentate nucleus: 10.8 mm posterior to the bregma, 3.6 mm lateral to the midline, and 6.5 mm ventral to the surface of the skull (Fig. 1), according to the rat brain atlas by Paxinos and Watson (2006). Pressure injection of 0.4 μL of BDA solution (10% BDA in 0.1 M phosphate-buffered saline [PBS]; Molecular Probes, Eugene, OR) into the dentate nucleus was performed using a Hamilton syringe (HAM-80008, 1701 SN 10 μL Syr).

Schematic illustration of the BDA injection site in the dentate nucleus (lateral nucleus) of the cerebellum in the five rats included in the study. The illustrations show the anteroposterior of the extension of the tracer within the dentate nucleus between bregma −10.52 and −11.60. BDA, biotinylated dextran amine.
After survival of 10 days, the animals received a lethal dose of pentobarbital (Narcoren) (100 mg/kg BW i.p.) and were transcardially perfused with saline (0.9% NaCl, pH 7.45; 200 mL), followed by 500 mL of phosphate-buffered fixative containing 2% paraformaldehyde (PFA) and 1.5% glutaraldehyde (GA) solution (in 0.1 M PBS, pH 7.45; PBS Amresco E404-200TABS; PFA MERCK 1.04005.1000; and GA MERCK 8.20603.1000).
Brains were removed from the skull, postfixed in the same fixative overnight, and stored in 30% sucrose solution in PBS at +4°C for 2 days. Coronal cryostat sections (40 μm) were cut using a cryomicrotome (Leica CM 1950) and collected in PB in tissue culture plates.
BDA histochemistry
Histochemistry was applied to every third section to visualize the BDA solution in the tissue, and sections were run in the avidin-biotinylated HRP (ABC) procedure, followed by the metal-enhanced diaminobenzidine (DAB-Ni) reaction.
The procedure started with 10 min of sodium borohydride (Sigma-Aldrich S38714-486) incubation; PBS 0.1 M washes, twice for 10 min each; ABC standard solution (Vector Laboratories PK6100; Vectastain Elite ABC Standard Kit) incubation for 3 h at room temperature; PBS 0.1 M wash for 10 min; Tris buffer (TB) 0.05 M (pH: 8.0) washes, twice for 10 min each; DAB-Ni [DAB Sigma D5905, ammonium nickel(II) sulfate hexahydrate Sigma A1827] preincubation in a dark room for 15 min; H2O2 reaction for 3–15 min in a dark room; TB 0.05 M (pH: 8.0) wash for 10 min, followed by PBS 0.1 M wash for 10 min; and finally, the sections were mounted.
The sections were examined at 40 × magnification under a light microscope.
Fiber tractography
Three-tesla high-resolution DTI data of 35 (20–59 years old; mean age = 31 years old) healthy subjects were obtained from open-source connectome datasets from the Human Connectome Project (HCP) (Mass General Hospital-University of Southern California [MGH-USC] Consortium). These DTI datasets were examined using a customized 3-T Connectome Scanner (Siemens, Skyra, Erlangen, Germany) unit with a 64-channel brain array coil and a novel gradient system (gradient strength: 70–100 mT/m) (Fan et al., 2016). Details of the HCP data are given on the internet.
Each dataset consists of isotropic 3D-T1W MPRAGE (time of repetition [TR]/time of echo [TE]/time of inversion [TI]: 2530/1.15/1100 ms, field of view [FOV]: 256 × 256 mm, voxel size: 1 × 1 × 1 mm3, Integrated Parallel Acquisition Techniques [IPAT]: 2, and acquisition time: 6 min), 3D-T2W SPACE (TR/TE: 3200/561 ms, FOV: 224 × 224 mm, voxel size: 0.7 × 0.7 × 0.7 mm3, IPAT: 2, and acquisition time: 6.48 min), and diffusion-weighted echo-planar imaging (with four different b-values) sequences. Details of diffusion scans were as follows; TR/TE: 8800/57 ms, FOV: 210 × 210 mm, matrix: 140 × 140, slice number: 96, voxel size: 1.5 × 1.5 × 1.5 mm, IPAT: 3, echo spacing: 0.63 ms, b-values: 1000, 3000, 5000, and 10,000 s/mm2, and total acquisition time: 89 min.
The MGH-USC Consortium has developed new methods for diffusion preprocessing (e.g., eddy current correction, distortions, and motion correction) of the different diffusion directions. Scanning details of 3-T MR and DTI acquisition protocols can be found in the appendix of the MGH-USC Consortium webpage.
In the present study, a probabilistic atlas and normalization for deep cerebellar nuclei were used to show the fibers of the dentate nucleus. Fiber tracking was performed by deterministic tractography, using seed points placed along with the amygdala and dentate nucleus, bilaterally. A probabilistic atlas of the human cerebellum was used for determination of the dentate nucleus.
The diffusion spectrum imaging (DSI) Studio tractography tool was used for data analysis. The tractography images of the dentate nucleus in these atlases were superimposed on T1-weighted images in the HCP data with the help of the DSI Studio software. The location of the dentate nucleus was determined. As a result, region of interest (ROIs) of the dentate nucleus were drawn both by manual segmentation with the help of the atlas and by using the previously determined automatic ROIs in the atlas.
In addition, ROIs were placed on the amygdala region using the DSI Studio software with the automated anatomical labeling (AAL3) atlas in the present study.
After corrections of eddy current-induced and motion artifacts, all images were coregistered to one another. After skull stripping or brain extraction, images were subsequently normalized to the Montreal Neurological Institute coordinates.
The parameters used for fiber orientation in this study are as follows: tracking index = 0, FA threshold = 0, turning angle = 0, step size = 0, min-max lengths = 20–300, track count = 5,000,000, tracking plan = 0, autotip = 16, autotrack tolerance = 16, dt_index = 0, dt_threshold = 0.2, tracking_method = 0, smoothing = 0, interpolation = 0, initial_direction = 0, seed_plan = 0, random_seed = 0, check_ending = 0, otsu_threshold = 0.6, track_format = 2, and thread_count = 1.
Results
BDA tract tracing
Of the 11 BDA injections, the results presented are from 5 animals in which the centers of the injection sites were optimal with minimal contamination of adjacent structures or along the pipette tract. The results of six animals were excluded from the study due to incorrect injection site (n = 4) or significant contamination of the BDA to the surrounding structures (n = 2).
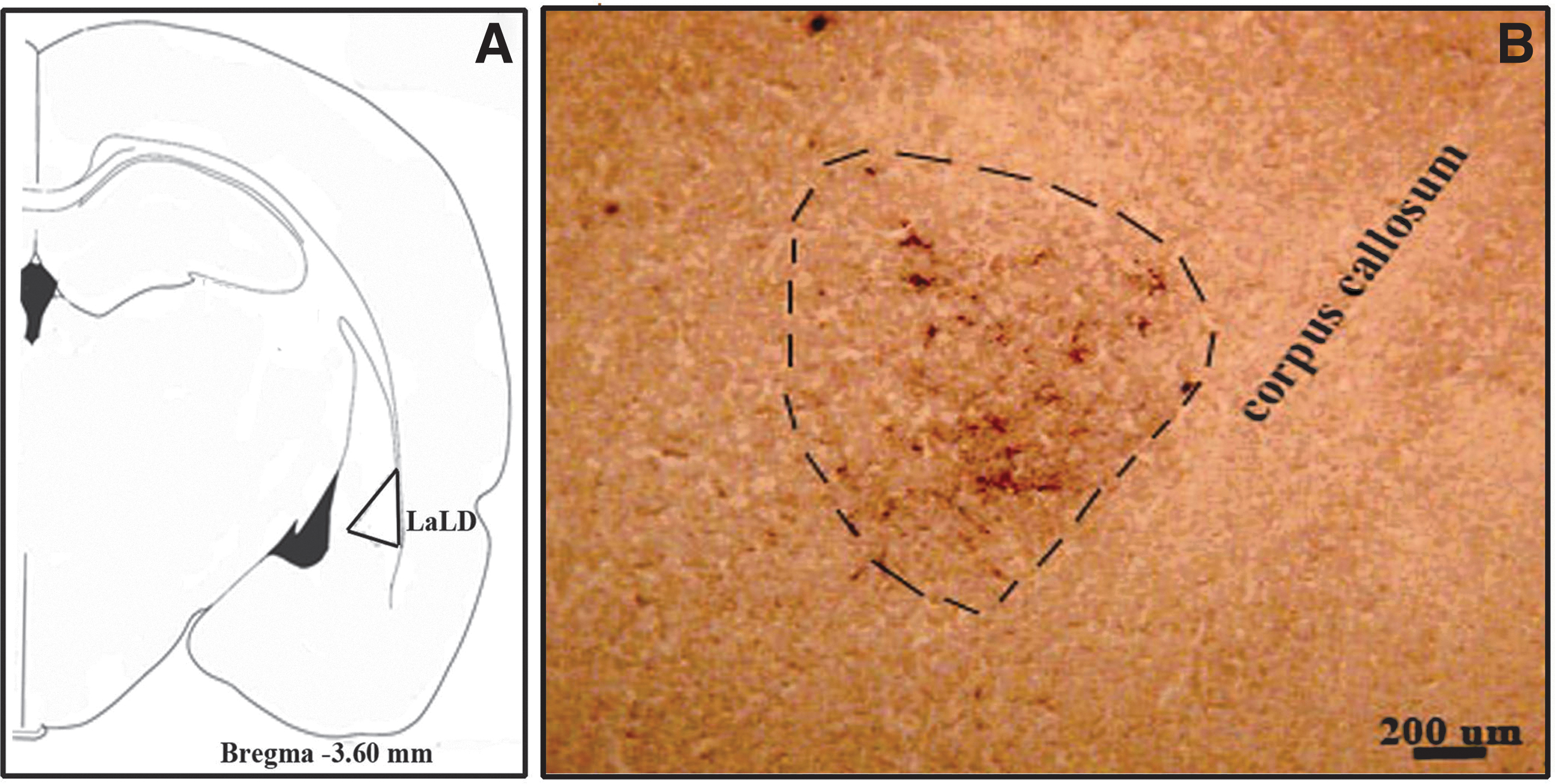
The injection sites of five animals are shown in Figure 1. The labeled axons and cells in amygdala nuclei, which were present in at least four animals, were included in the results. The amygdala is a nuclear complex located deep in the anteromedial temporal lobe in the rat brain. It is related dorsomedially to the striatum and laterally to the insular cortex. The nuclei of the amygdala extend between 1.60 and 4.80 mm posterior to the bregma in the rat brain.
Subsequent to BDA injections into the dentate nucleus, labeled cells and axons were documented in the central (CeC, CeM, and CeC), basal (BLA), and lateral (LaLD) nuclei of the amygdala. Bilateral labeled cells and axons were observed in the central (Fig. 2A–C) and basal (Fig. 3A–C and Table 1) nuclei of the amygdala, being dense on the ipsilateral side and sparse on the contralateral side.


The Density of Bilateral and Ipsilateral Projections of the Dento-Amygdala Projections
Numerous labeled cells: indicates >25 labeled cells.
Moderately labeled cells: indicates 15–20 labeled cells.
Sparsely labeled cells: indicates <10 labeled cells.
—, no labeled cells.
However, only ipsilateral labeled cells and axons were observed in the lateral amygdala nucleus (Fig. 4A, B, and Table 1). The contralateral fibers crossed in the decussation of the cerebellar peduncle at the midbrain, and ipsilateral dentate fibers reached the amygdala through the superior cerebellar peduncle (SCP) (Fig. 5A, C, D).

DTI of cerebello-amygdala connections
Analogous to the data obtained from the rat tract-tracing study, unilateral and bilateral dento-amygdala fibers were observed in most of the cases (Figs. 6–8). The dento-amygdala fibers ascended within the SCP and most fibers reached the ipsilateral amygdala, and fewer fibers crossed at the level of the midbrain and reached the contralateral amygdala nuclei (Figs. 7 and 8).
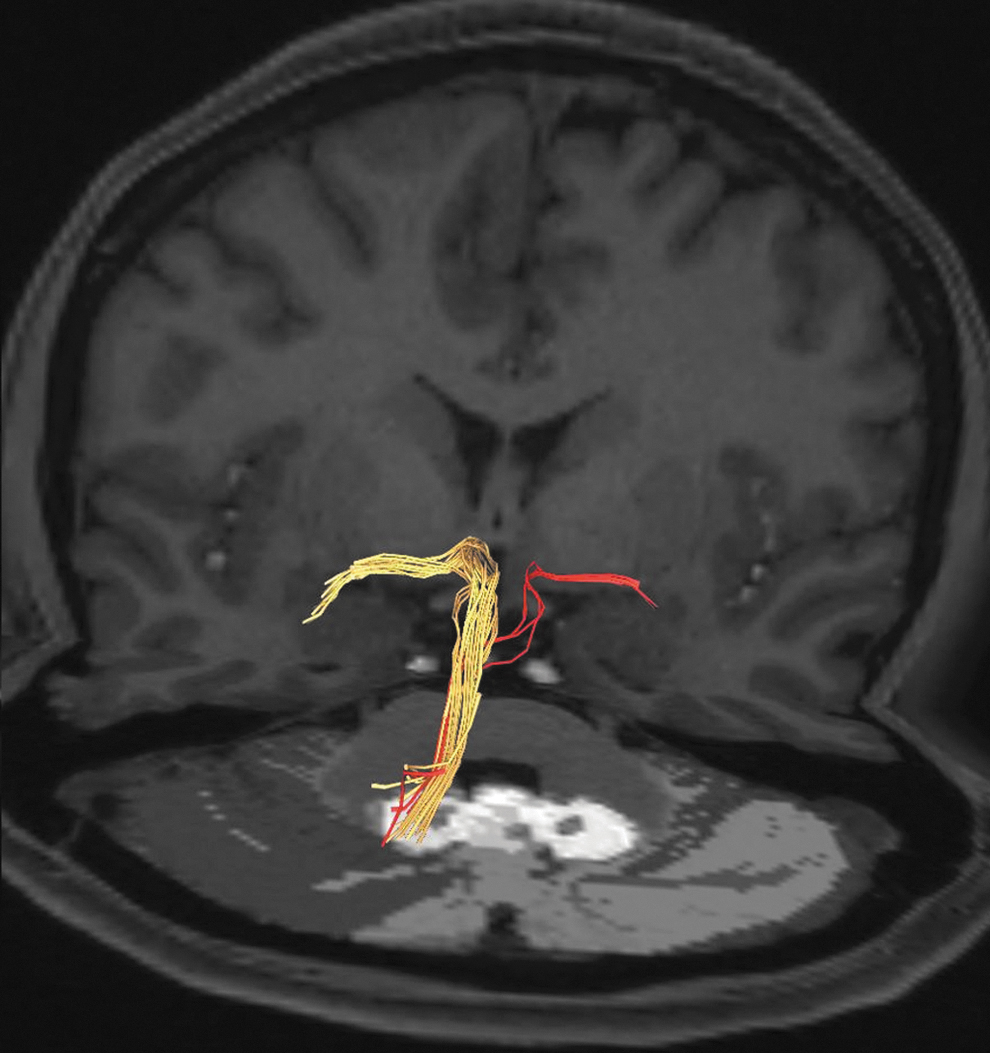
Tractography images of a 39-year-old female showing the dentate fibers reaching both left and right amygdalae, the majority of the fibers to the ipsilateral (yellow-colored fiber), and few fibers (light-colored fibers) to the contralateral amygdala.

The tractography image of a 44-year-old woman showing the pathways from both dentate nuclei to the ipsilateral (white arrow) and contralateral (yellow arrow) amygdala. The majority of the fibers from the dentate nucleus pass through the hypothalamus to reach the amygdala (double-headed yellow arrow). The decussation of the SCP at the midbrain is indicated by the red arrow. In addition, the tracts between the fastigial nucleus and amygdala are highlighted (blue arrow).

The image of a 44-year-old female showing left dento-amygdala projections from the dentate nucleus to the amygdala through the hypothalamus (yellow arrows) and without passing through the hypothalamus (red arrows), and their bifurcation.
In the majority of cases, we also observed that fibers arising from the dentate nucleus passed through the hypothalamus before reaching the amygdala, and few fibers reached the amygdala directly (Figs. 7–9). Additionally, the DTI analyses show that the left and right amygdalae were connected to each other through transverse fibers traveling within the anterior commissure (Fig. 9).
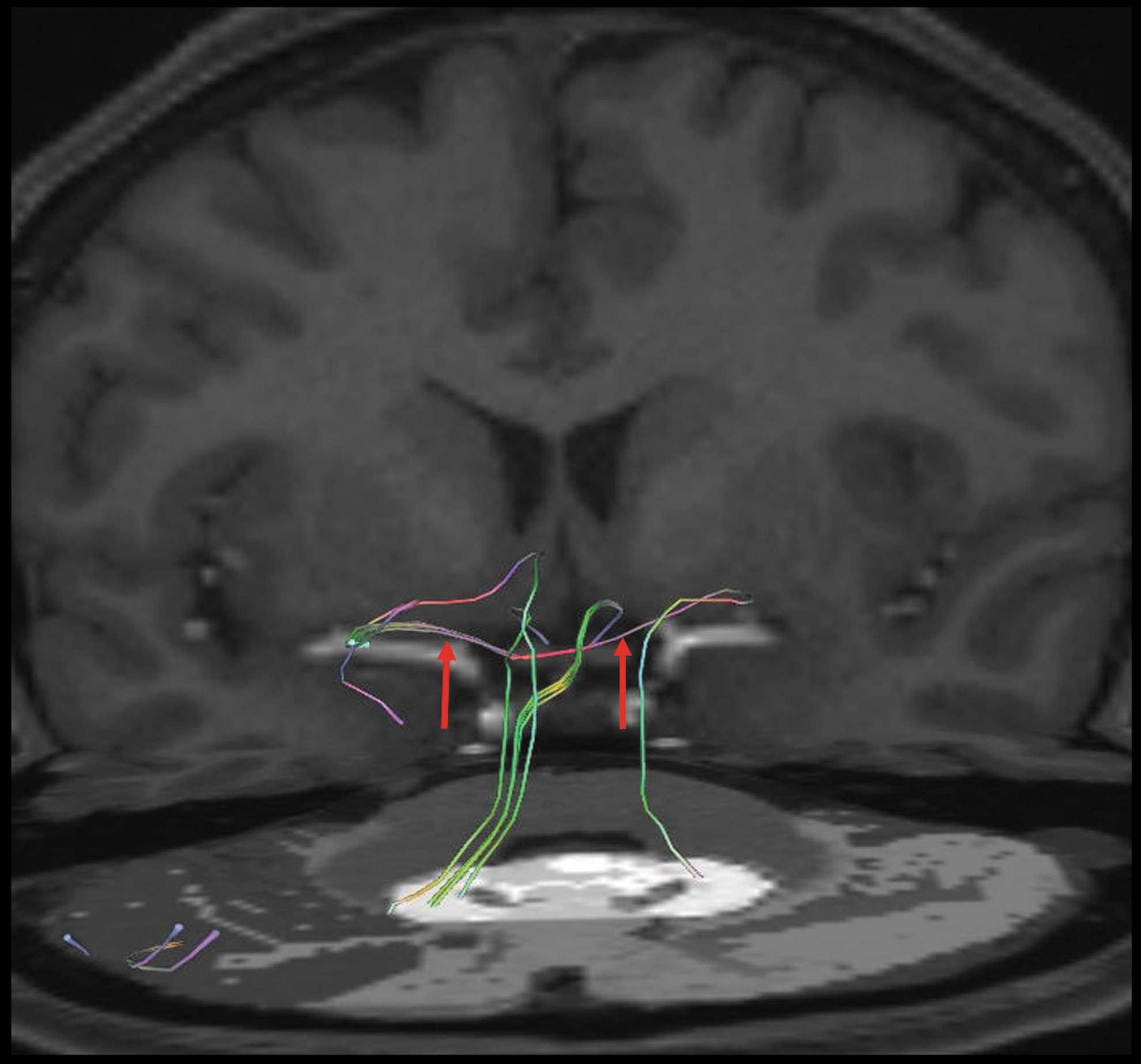
Tractography images of a 39-year-old female demonstrating the pathway connecting the left and right amygdalae through the anterior commissure (red arrows).
Unfortunately, we were unable to define specific amygdala nucleus termination of the dento-amygdala fibers.
Discussion
Major findings
The results of the present experimental tract-tracing study clearly showed projections between the dentate nucleus and the central, basal, and lateral nuclei of the amygdala. The projections to the central and basal nuclei of the amygdala were bilateral, dense on the ipsilateral and sparse on the contralateral amygdala nuclei. However, only ipsilateral projections were observed with the lateral nucleus of the amygdala.
The fibers from the dentate nucleus reached the amygdala through the SCP, the contralateral fibers crossed in the decussation of the SCP at the midbrain. The results of the experimental study on rats corresponded with the DTI study on humans. Furthermore, tractography results showed that dentate fibers pass through the hypothalamus before reaching the amygdala.
Dento-amygdala connections
Experimental tract-tracing studies showed that the cerebellum not only projects to cortical and subcortical motor areas but also nonmotor regions of the brain (Çavdar et al., 2020; Strick et al., 2009). Early studies showed evidence of cerebellar connections with the amygdala (Heath et al., 1978; Nashold and Slaughter, 1969).
Nashold and Slaughter (1969) stimulated the dentate nucleus and SCP in a man and the subject experienced an unpleasant sensation of fear. Furthermore, vermal lesions of the cerebellum in animal experiments resulted in a decrease in the reactivity of animals to fear stimuli (Nashold and Slaughter, 1969).
Furthermore, electrical stimulation of the anterior cerebellum induced responses such as arousal, predatory attack, and feeding, which mimic those obtained by amygdala stimulation (Snider and Maiti, 1976).
An experimental, double-tracer labeling study on rats by Dietrichs and Haines (1986) showed that retrogradely labeled neurons from the cerebellum or amygdala were found in the same hypothalamic nuclei. They concluded that some hypothalamic neurons may project to the cerebellum and also give collaterals to the amygdala (Dietrichs and Haines, 1986).
Moreover, supporting the former study, an evoked response in the amygdala and hypothalamus has been recorded through stimulation of the cerebellum (Anand et al., 1959; Heath et al., 1978). In our previous research, we clearly defined the dento-hypothalamic projections (Çavdar et al., 2020). In the present study, we used a single tracer, therefore we could not detect whether the hypothalamic labeling were collaterals of dento-amygdala projections or direct projections.
However, the 3-T DTI data clearly show that most of the dentate fibers pass through the hypothalamus before reaching the amygdala. Thus, this may indicate a circuit between the dentate nucleus, hypothalamus, and amygdala, which may show the role of the dentate nucleus in modulation of autonomic and emotional functions.
The BDA tracer used in the present study is not only a sensitive anterograde tracer but also a retrograde marker. A clinical dynamic causal modeling study demonstrated that during rhyming judgment tasks the cerebellum and the anterior temporal lobe are strongly and bidirectionally interconnected.49 Thus, the present experimental study on rats is in accordance, showing bidirectional projections between the dentate and amygdala nuclei. DTI holds enormous potential to study the white matter of the human brain; however, it has some limitations (Çavdar et al., 2020; Jbabdi and Johansen-Berg, 2011). One of the limitations is the discrimination of afferent and efferent fibers; unfortunately, it does not show whether these fibers are afferent or efferent from the dentate nucleus.
An early study on monkeys and cats showed that the fastigial nucleus projected to the amygdala (Heath and Harper, 1974). Furthermore, stimulation of the fastigial nucleus produces drowsiness and electroencephalogram changes, which resemble sleep patterns resulting from stimulation of the ventral amygdala (Snider and Maiti, 1976). The central amygdala (CeA) has been shown to modulate cerebellar learning by facilitating sensory input to the cerebellar cortex and interpositus nucleus (Farley et al., 2016).
We have shown the presence of fastigial nucleus-amygdala connections in the DTI of human subjects. These results indicate that not only the dentate but also other deep cerebellar nuclei are connected to the amygdala, which may serve different functions.
Cerebellar dysfunction and emotions
An early study by Papez (1937) has shown bidirectional connections of the cerebellum with a large portion of the limbic lobe and the associated subcortical nuclei, which are regions of interest to psychiatry through their association with emotional processing. Clinical studies on 31 patients with degenerative cerebellar diseases showed that 77% of the patients had psychopathology, particularly depression and personality change (Leroi et al., 2002; Papez, 1937).
A recent study described a resting-state, bilateral cerebello-amygdala network using independent component analysis and suggested that these connections could be in charge of sensorimotor, emotional, and motivational integration of somesthetic stimuli before recruiting more specialized circuits such as the ventral striatum or attentional and salience networks (Habas, 2018).
Furthermore, radiological studies demonstrated the involvement of the cerebellum in psychiatric and cognitive disorders in patients with degenerative cerebellar diseases, including schizophrenia, anxiety disorders, depression, and mania (Lauterbach, 1996; Leroi et al., 2002; Picard et al., 2008). Studies of patients with major depressive disorder showed a reduction in the connectivity of the amygdala with brain regions involved in emotional processing and regulation, including the ventrolateral prefrontal cortex; insula; caudate, middle, and superior temporal regions; occipital cortex; and cerebellum (Phillips et al., 2015).
The results of the present study clearly show the dento-amygdala connections. Moreover, the dentate nucleus has extensive connections with the thalamus and brainstem nuclei (brainstem reticular nucleus, locus coeruleus, and substantia nigra) (Cohen et al., 1958; Haubrich et al., 2020), which are connected to the amygdala. Thus, the dentate nucleus can influence the amygdala indirectly through dento-thalamo-amygdala or dento-brainstem nucleus-amygdala connections as well.
Brain areas interact with each other to orchestrate cognition and behavior. Specific differences in brain connectivity among species have been documented (Barrett et al., 2020). It has been shown that the fronto-occipital connections and the arcuate fasciculus were more prominent in humans than in monkeys (Barrett et al., 2020).
Furthermore, Schaeffer et al. (2020) compared the medial frontal cortex (MFC) organization between rodents, nonhuman primates (marmosets), and humans and showed similar intrinsic functional organization of the MFC across the three species, but clear differences between rodent and primate MFC whole-brain connectivity were observed (Schaeffer et al., 2020).
We were unable to compare the dento-amygdala connections between rats and humans because we do not have an MRI device specially designed for rats and no literature was available regarding the tractography of dento-amygdala connections in the rat brain.
Limitations of the study
The DTI-based deterministic tractography technique shows limitations in the quantitation of measurements and reconstruction of crossing fibers. Quantitative anisotropy techniques may be more useful in detecting the crossing fibers. We may not have been able to detect all ipsilateral or contralateral fibers due to the intrinsic limitations of the 3-T DTI technique.
There is a need for new and more comprehensive studies using higher tesla MR devices with dedicated coils/techniques and up-to-date hardware related to these subjects.
Conclusions
The present experimental rat and human DTI study showed direct amygdala projections. Detailed knowledge of dento-amygdala connections can explain the emotional and cognitive deficits that occur in patients with cerebellar or SCP damage.
Furthermore, being aware of the course of the dento-amygdala fiber can potentially minimize damage during regional surgery.
Footnotes
Acknowledgments
We would like to thank Koc University for allowing us to use the facilities of Research Center for Translational Medicine (KUTTAM). The DTI data was provided from the Human Connectome Project (HCP).
Ethical Approval
All procedures performed in the studies involving human participants were approved by the institutional ethics committee.
Informed Consent
Informed consent was obtained from all individual participants for the DTI part of the study by the Human Connectome Project, WU-Minn Consortium.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this study.