
Abstract
Background:
Recent neuroimaging studies on upper-limb amputation have revealed the reorganization of bilateral sensorimotor cortex after sensory deprivation, underpinning the assumption of changes in the interhemispheric connections. In the present study, using functional magnetic resonance imaging (fMRI) and diffusion tensor imaging (DTI), we aim to explore the alterations in the interhemispheric functional and structural connectivity after upper-limb amputation.
Methods:
Twenty-two upper-limb amputees and 15 age- and sex-matched healthy controls were recruited for MRI scanning. The amputees were further divided into subgroups by amputation side and residual limb pain (RLP). DTI metrics of corpus callosum (CC) subregions and resting-state functional connectivity (FC) between the bilateral sensorimotor cortices were measured for each participant. Linear mixed models were carried out to investigate the relationship of interhemispheric connectivity with the amputation, amputation side, and RLP.
Results:
Compared with healthy controls, upper-limb amputees showed lower axial diffusivity (AD) in CC subregions II and III. Subgroup analyses showed that the dominant hand amputation induced significant microstructural changes in CC subregion III. In addition, only amputees with RLP showed decreased fractional anisotropy and AD in CC, which was also correlated with the intensity of RLP. No significant changes in interhemispheric FC were found after upper-limb amputation.
Conclusion:
The present study demonstrated that the interhemispheric structural connectivity rather than FC degenerated after upper-limb amputation, and the degeneration of interhemispheric structural connectivity was shown to be relevant to the amputation side and the intensity of RLP.
Impact statement
Neuroimaging studies have revealed the functional reorganization of bilateral sensorimotor cortex after amputation, with expanded activation from the intact hemisphere to the deprived hemisphere. Our findings indicated a degeneration of interhemispheric white matter connections in upper-limb amputees, unveiling the underlying structural basis for bilateral functional reorganization after amputation.
Introduction
The loss of unilateral limb not only results in structural and functional degeneration in the deprived sensorimotor cortex (Bao et al, 2021; Hamzei et al, 2001; Molina-Rueda et al, 2019), but also induces changes in the cortical representation of the intact limb. Expanded activation to the deprived sensorimotor cortex has been widely observed during sensory stimulation or movement of the intact limb (Bogdanov et al, 2012; Makin et al, 2013a), underpinning the assumption of changes in the interhemispheric connections. Interhemispheric connectivity has been reported to be disrupted either functionally or structurally in lower-limb amputees (Bramati et al, 2019; Li et al, 2017). However, there is relatively poor understanding of interhemispheric connectivity change after upper-limb amputation.
A pilot study has reported reduced interhemispheric functional connectivity (FC) in sensorimotor network after upper-limb amputation (Makin et al, 2013b), but this change was found to be primarily correlated with phantom limb pain (PLP). The neuroimaging correlates of upper-limb amputation itself still remain unclear. The objectives of this study were to explore the alternations in both interhemispheric structural and FC after upper-limb amputation.
In addition, potential factors affecting interhemispheric connectivity are also of interest. Besides PLP, multiple amputation-related factors have shown their power in brain reorganization, such as amputation side (Guo et al, 2019a), residual limb pain (RLP) (Guo et al, 2019b), and prosthesis use (Granata et al, 2020; Lotze et al, 1999). These factors could also play roles in disrupting interhemispheric connectivity. In this study, the amputation side and RLP were thus selectively analyzed according to the characteristics of recruited samples. Dominant-side amputation and nondominant-side amputation impact differently on daily activities (Yancosek and Mullineaux, 2011), and thus would likely lead to different long-term reorganization in the brain. A study on dominant-hand immobilization found a decrease of fractional anisotropy (FA) in the corticospinal tract (Langer et al, 2012), while another study on most subjects (13/18) with the loss of nondominant hand did not observe significant reduction of FA (Makin et al, 2013a).
Our previous study also suggested that the extent of reorganization of whole-brain structural network was associated with the amputation side (Guo et al, 2019a). Therefore, we hypothesized that the dominant-hand amputation may induce more disruption of interhemispheric connectivity than the nondominant-hand amputation. Another factor we are interested in is RLP, a type of pain felt in the residual limb. RLP causes greater interference in the daily life activities than PLP for its high intensity (Gallagher et al, 2001). Multiple types of pain experiences have shown their correlation with disruption of interhemispheric connectivity (Kim et al, 2014; Sevel et al, 2016), including PLP (Makin et al, 2013b). RLP has been correlated with brain reorganization from the aspects of brain metabolites and activity (Guo et al, 2019b), and likely takes an effect on interhemispheric connectivity.
In this study, interhemispheric connectivity was assessed both functionally and structurally by resting-state functional magnetic resonance imaging (fMRI) and diffusion tensor imaging (DTI). The major structural commissure connecting the bilateral sensorimotor areas is the corpus callosum (CC) (de Lacoste et al, 1985). CC plays an important role in the interhemispheric functional integration of the perceptual, cognitive, learned, and volitional information (Clarke and Zaidel, 1994). Previous studies have established a topographical distribution of interhemispheric fiber connections to different brain regions in the midsagittal section of the CC (Hofer and Frahm, 2006), and the microstructure of CC subregions, which is assessed by DTI, can be used to characterize interhemispheric structural connectivity between homologous brain regions (Li et al, 2017; Serbruyns et al, 2015).
We hypothesized that (1) upper-limb amputation may disrupt interhemispheric structural and FC between the bilateral sensorimotor cortices, and (2) RLP and the amputation side may play a role in the disruption of interhemispheric connectivity after upper-limb amputation.
Methods
Participants
Twenty-three upper-limb amputees and 16 healthy controls were recruited in this study. All participants reported no history of neurological or mental disorders. All participants were right-handed (handedness before amputation for amputees) according to a Chinese version of a standardized handedness inventory (Li, 1983), except for one amputee (A19) who was amputated at the age of two and could not complete the handedness inventory. For each amputee, PLP and RLP were assessed by a phantom- and stump-phenomena interview proposed by Winter et al (2001). If amputees had experienced pain on their injury limb within the past 3 months, they were asked to first localize the pain to distinguish between RLP and PLP, and then report a score on a 0–10 scale (0 = no pain, 10 = worst pain imaginable) based on the average intensity of pain within the last month.
The details of the experiment were described in the previous study (Guo et al, 2019a). The IRB approval of study was thus waived for it is only a re-analysis of the existing data of that study. The use of data was in compliance with the Declaration of Helsinki.
One amputee and one healthy control with obvious motion artifacts on MRI images were excluded, and therefore, 22 upper-limb amputees (age: 52.95 ± 7.42 years, sex: 16M/6F, amputation side: 7L/15R, years postamputation: 23.95 ± 13.70 years) and 15 healthy controls (age: 50.13 ± 8.63 years, sex: 11M/4F) were included in the following MRI analysis. The included amputees and healthy controls were matched in age and sex (both p > 0.540). To investigate the influence of amputation side or pain experience on interhemispheric connectivity, amputees were divided into subgroups according to amputation side and RLP, respectively, that is, the right-side amputees (n = 15) and the left-side amputees (n = 7), or the amputees with RLP (RLP amputees, RLP score >0, n = 9) and the amputees without RLP (non-RLP amputees, RLP score = 0, n = 13). The detailed demographic and amputation-related information of the amputees is summarized in Table 1.
Demographic and Amputation-Related Information of Amputees
PLP and RLP were reported on a 0–10 scale based on the average intensity of his/her pain within the last month (0 = no pain, 10 = worst pain imaginable).
Dominant hand before amputation.
Age at amputation.
M, male; F, female; L, left; R, right; Prosthesis, whether or not wearing any prosthesis; Y, yes; N, no; PLP, phantom limb pain; RLP, residual limb pain.
Image acquisition
MRI was performed on a 3T scanner (Philips, Achieva 3.0T TX, Netherlands) with an 8-channel head coil in the Second People's Hospital of Changzhou, China. The MRI session included a T1-structural scan, a spectroscopy scan, a DTI scan, and two functional scans in resting and under tactile stimuli on the residual limb, respectively. DTI and rest fMRI images were analyzed in this study, and the T1-weighted images were used for coregistration. During the scanning, the subjects were instructed to keep awake and rest with eyes closed. T1-weighted images were acquired with a 3D-TFE sequence (repetition time [TR] = 8.2 msec, echo time [TE] = 3.7 msec, flip angle = 8°, field of view [FOV] = 250 × 250 mm2, number of slices = 162, imaging matrix = 256 × 256, voxel size = 0.976 × 0.976 × 1.00 mm3).
DTI images were obtained with a spin echo-planar imaging sequence (TR = 7444.428 msec, TE = 85 msec, flip angle = 90°, number of signal averages = 2, FOV = 224 × 224 mm2, number of slices = 60, slice thickness = 2 mm, no gap, matrix = 112 × 112, orientation = axial, 32 nonlinear diffusion weighting gradient directions with b = 800 sec/mm2, and 1 additional image without diffusion weighting). Rest fMRI images were acquired with a field echo echo-planar imaging sequence (TR = 2000 msec, TE = 32 msec, flip angle = 82°, FOV = 230 × 230 mm2, number of slices = 35, imaging matrix = 128 × 128, voxel size = 1.875 × 1.875 × 3.00 mm3).
DTI image preprocessing
DTI image preprocessing was performed using FSL (University of Oxford, United Kingdom) (Smith et al, 2004). After the corrections for head motion, eddy current distortions, and removal of skull and nonbrain tissue, tensor estimation was performed to get FA (the degree of anisotropy of diffusion), mean diffusivity (MD, the rotationally invariant magnitude of diffusion within brain tissue), axial diffusivity (AD, the magnitude of diffusion parallel to fiber tracts), and radial diffusivity (RD, the magnitude of diffusion perpendicular to fiber tracts) images in DTI space. DTI images were coregistered to T1 images and then converted into Montreal Neurological Institute (MNI) standard space according to the transformation relationship between T1 space and MNI standard space. The same transformations were applied in FA, MD, AD, and RD images.
DTI-derived estimates of CC structural characteristics
CC standard template in MNI space was extracted by combing its genu, body, and splenium parts based on Johns Hopkins University DTI-based white-matter atlases, and then segmented into five subregions according to a classification scheme proposed by Sabine Hofer et al (Hofer and Frahm, 2006). Subregion I connects the bilateral prefrontal regions, subregion II connects the bilateral premotor cortex (PMC) and supplementary motor areas (SMA), subregion III connects the bilateral primary motor cortex (M1), subregion IV connects the bilateral primary sensory cortex (S1), and subregion V connects the bilateral parietal, temporal, and occipital regions (Fig. 1). As amputation principally affects motor and sensory areas such as M1 (Hamzei et al, 2001; Jiang et al, 2015), S1 (Jiang et al, 2015), and PMC and SMA (Bao et al, 2021; Hamzei et al, 2001), the CC subregions II, III, and IV were thus selected as regions of interest (ROIs) to calculate the DTI metrics (FA, MD, AD, and RD).
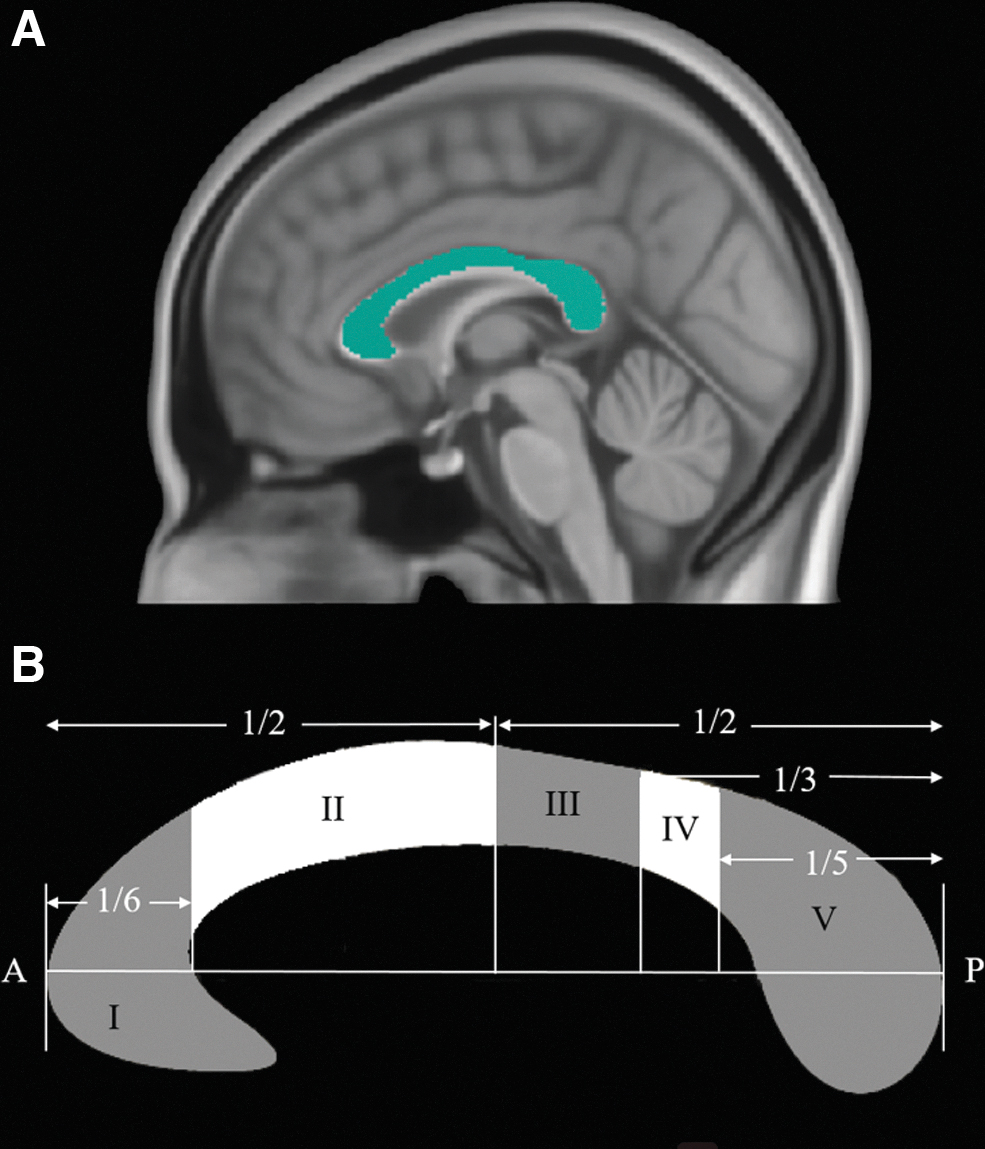
CC in the midsagittal slice.
fMRI image preprocessing and FC
Rest fMRI images were preprocessed using DPABI, a toolbox for Data Processing & Analysis for Brain Imaging (Yan et al, 2016). The preprocessing steps included discarding the first 10 volumes, slice timing correction, motion artifact correction, normalization to MNI space, smoothing with an 8 mm Gaussian blur, and filtering.
After preprocessing, six ROIs (including the bilateral M1 [Brodmann 4], S1 [Brodmann 3, 1, and 2], and PMC and SMA [Brodmann 6]) were selected for interhemispheric FC analysis. The blood oxygenation level dependent (BOLD) signals for all pixels in each ROI were averaged, and the Pearson correlation coefficient between the bilateral counterparts was calculated as the corresponding interhemispheric FC by GRETNA (Wang et al, 2015). All FC was measured in z-scores.
Statistics
All statistical analyses were performed using R Statistical Software (Team, 2011). Between-group differences in demographic and amputation-related information were assessed by using independent t-tests (for the variables conforming to normal distribution), Mann–Whitney U tests (for the variables not conforming to normal distribution), or chi-square tests (for sex and amputation side). The normality of variables was tested by Shapiro–Wilk's test. To identify the effects of amputation, amputation side, and RLP on the interhemispheric connectivity of sensorimotor areas, we fitted the data with two different types of linear mixed models (LMMs). First, when comparing amputees or subgroups of amputees with healthy controls, the LMMs included Group, Region, Region × Group interaction, Age, and Sex as the fixed effects, and Subject as the random effect (formula: Connectivity ∼ Group × Region + Age + Sex +1 | Subject).
Post hoc tests were used to test the effect of “Group” factor (amputees vs. controls, right-side amputees vs. left-side amputees vs. controls, RLP amputees vs. non-RLP amputees vs. controls), with the healthy control group as the reference on interhemispheric connectivity at different regions. Bonferroni correction was used in multiple comparisons. Second, to identify the effects of amputation side and RLP, we also made comparisons between subgroups of amputees (right-side amputees vs. left-side amputees; RLP amputees vs. non-RLP amputees) using another type of LMMs, adding Years postamputation and Age × Years postamputation interaction as additional fixed effects (formula: Connectivity ∼ Group × Region + Age × Years postamputation + Sex +1 | Subject). FA, MD, AD, RD, and FC were, respectively, fitted with the model as dependent variables.
Kenward–Roger test was used as post hoc test for testing linear hypotheses about fixed effects. All LMMs were performed with the R package lme4 lmerTest and emmeans (Bates et al, 2015; Kuznetsova et al, 2017; Piepho, 2004). Correlations of connectivity with significant effect of amputation with RLP and RLP intensity were evaluated by Spearman's correlation analysis. All data are presented as mean ± standard deviation.
Results
Comparison of DTI metrics of CC between upper-limb amputees and healthy controls
We found that upper-limb amputation was associated with the change in AD of the CC (Table 2; Supplementary Fig. S1). LMMs showed a significant effect of amputation on the mean AD of CC subregion II (β = −0.071, 95% CI [−0.125 to −0.0018], t(101) = −2.65, p = 0.011, Cohen's d = −1.67) and subregion III (β = −0.064, 95% CI [−0.117 to −0.010], t(101) = −2.36, p = 0.020, Cohen's d = −1.49). The effect of amputation was statistically nonsignificant on FA, MD, and RD.
Magnetic Resonance Imaging Measurements for Three Interhemispheric Connections in Both Amputees and Healthy Controls
p < 0.05.
AD, axial diffusivity; FA, fractional anisotropy; FC, functional connectivity; M1, primary motor cortex; MD, mean diffusivity; MRI, magnetic resonance imaging; PMC, premotor cortex; RD, radial diffusivity; S1, primary sensory cortex; SMA, supplementary motor areas.
Comparison of DTI metrics of CC among the right-side amputees, the left-side amputees, and healthy controls
The right-side amputees and the left-side amputees did not show significant differences in age, sex, years postamputation, amputation level, and RLP (all p > 0.258). Their age and sex were also comparable with healthy controls (all p > 0.459).
In the LMMs with the reference of healthy control group, the effect of both right-side amputation (β = −0.068, 95% CI [−0.128 to 0.0091], t(98) = −2.29, p = 0.024, Cohen's d = −1.599) and left-side amputation (β = −0.078, 95% CI [−0.152 to −0.0041], t(98) = −2.10, p = 0.039, Cohen's d = −1.827) was statistically significant and negative in the prediction of AD in CC subregion II (Table 3; Supplementary Fig. S2). Moreover, LMMs showed a significant effect of right-side amputation on the AD (β = −0.067, 95% CI [−0.126 to −0.0077], t(98) = −2.24, p = 0.027, Cohen's d = −1.567) and FA (β = −0.04, 95% CI [−0.07 to −0.0008], t(98) = −2.03, p = 0.045, Cohen's d = −1.459) of CC subregion III. The reduction of AD in CC subregion II in the right-side amputees was significant after the Bonferroni correction.
Magnetic Resonance Imaging Measurements for Three Interhemispheric Connections in Both Right-Side and Left-Side Amputees and Healthy Controls
Uncorrected p < 0.05.
Bonferroni corrected p < 0.05.
In the LMMs comparing the subgroups of amputees, the contrast between the right-side amputees and the left-side amputees was not significant (all p > 0.172).
Comparison of DTI metrics of CC among RLP amputees, non-RLP amputees, and healthy controls
The RLP amputees and the non-RLP amputees did not show significant differences in age, sex, amputation side, amputation level, and years postamputation (all p > 0.104). Both amputee subgroups were comparable with healthy controls in sex and age (all p > 0.191).
In the LMMs with the reference of healthy controls group, amputation with RLP contributed to a lower mean AD in CC subregion II (β = −0.096, 95% CI [−0.162 to −0.029], t(98) = −2.87, p = 0.005, Cohen's d = −2.249), III (β = −0.107, 95% CI [−0.173 to −0.041], t(98) = −3.21, p = 0.002, Cohen's d = −2.515), and IV (β = −0.087, 95% CI [−0.153 to −0.021], t(98) = −2.61, p = 0.011, Cohen's d = −2.040) (Table 4; Supplementary Fig. S3), and all these results passed the Bonferroni correction. In addition, the effect of RLP amputation on the mean FA of CC subregion II (β = −0.05, 95% CI [−0.09 to −0.0053], t(98) = −2.22, p = 0.028, Cohen's d = −2.040) and III (β = −0.05, 95% CI [−0.09 to −0.004], t(98) = −2.16, p = 0.033, Cohen's d = −1.866) was statistically significant and negative.
Magnetic Resonance Imaging Measurements for Three Interhemispheric Connections in Both Residual Limb Pain Amputees and Nonresidual Limb Pain Amputees and Healthy Controls
Uncorrected p < 0.05.
Bonferroni corrected p < 0.05.
The effect of amputation without RLP was statistically nonsignificant in FA, MD, AD, and RD. In the LMMs comparing the subgroups of amputees, the effect of RLP contributed to a lower MD in CC subregion IV (β = −0.06, 95% CI [−0.10 to −0.02], t(54) = −2.88, p = 0.006, Cohen's d = −1.693) and a lower AD in CC subregion III (β = −0.09, 95% CI [−0.17 to −0.004], t(54) = −2.10, p = 0.041, Cohen's d = −1.919) and IV (β = −0.09, 95% CI [−0.17 to −0.01], t(54) = −2.30, p = 0.025, Cohen's d = −2.103).
Further correlation analyses revealed that the RLP intensity was significantly and negatively correlated with FA in the subregion II (ρ = −0.681, p = 0.043; Fig. 2A), and AD in the subregion II (ρ = −0.672, p = 0.047; Fig. 2B), III (ρ = −0.732, p = 0.025; Fig. 2C), and IV (ρ = −0.894, p = 0.001; Fig. 2D).
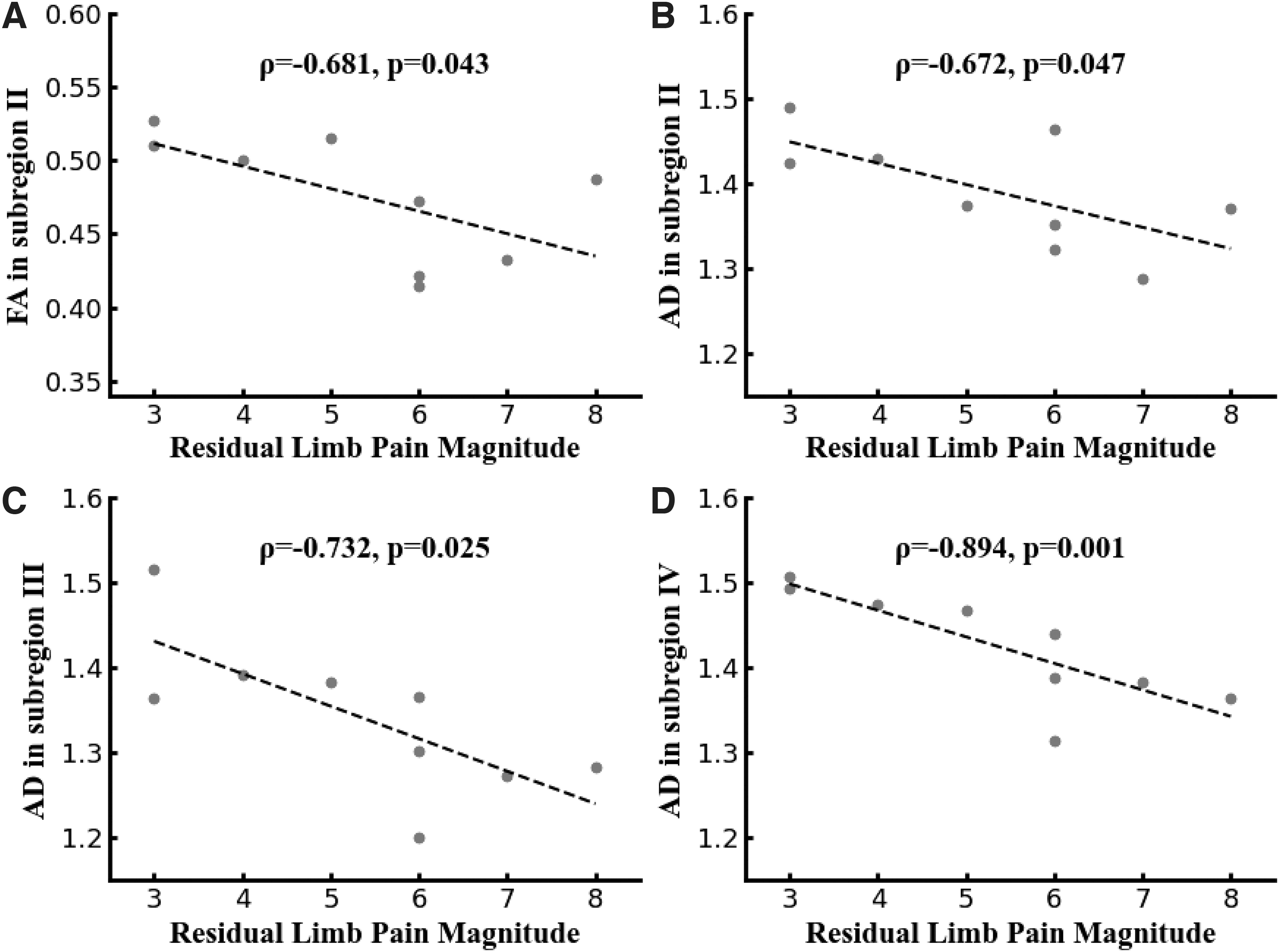
Spearman's correlation coefficients between RLP intensity and DTI metrics in three callosal subregions.
Comparison of interhemispheric FC
The interhemispheric FC between the bilateral M1, PMC and SMA, and S1 was predicted by LMMs (Tables 2–4), which showed no significant effect of amputation, amputation side, or pain experience (all p > 0.054).
Discussion
Generally, the microstructure of CC was found to be changed after upper-limb amputation, suggesting a disruption of interhemispheric structural connectivity in these subjects. Microstructural reorganization in CC has been observed in lower-limb amputees (Li et al, 2017; Simões et al, 2012). Through whole-brain tract-based spatial statistics, Simões et al (2012) found decreased FA in the body of the CC (mostly overlaps with the subregion II, III, and IV) in lower-limb amputees. Using subregion analysis, Li et al (2017) reported alteration in CC subregion II in lower-limb amputees. In this study of upper-limb amputees, a significant reduction of AD was found in the CC subregion II and III, which connect the bilateral PMC, SMA, and M1 (Hofer and Frahm, 2006).
Since AD reduction was often considered a biomarker of axonal degeneration (Castellano et al, 2016; Kim et al, 2011; Selvadurai et al, 2021; Song et al, 2003), our results suggested the degeneration of interhemispheric white matter connections after upper-limb amputation. With bilateral sensorimotor areas exhibiting interhemispheric inhibition (IHI) (Blankenburg et al, 2008; Ferbert et al, 1992), many studies have reported that amputation could decrease the level of IHI and promote the expanding representation of intact limb from ipsilateral hemisphere to contralateral hemisphere (Bogdanov et al, 2012; Makin et al, 2013a). Since CC provided the structural pathway of IHI (Cook, 1984; Denenberg et al, 1986), CC degeneration provides a structural basis for reduced IHI after amputation.
However, different from our observation in upper-limb amputees, the AD of CC was reported to be increased in lower-limb amputees (Li et al, 2017). We speculate that the inconsistency may be associated with prosthesis use. All the lower-limb amputees in Li et al (2017) relied on prothesis to stand and walk, but only four upper-limb amputees in this study used a prosthesis. In upper-limb amputees, extensive use of a myoelectric prosthesis in upper-limb amputees was associated with reduced cortical reorganization in M1, S1 (Lotze et al, 1999), and visual streams (Preißler et al, 2013). Di Pino et al (2009) suggested that the prosthetic system could be seen as a neurorehabilitation tool, which could induce reduction in “aberrant” plasticity and promote “good” plasticity. In our sample of upper-limb amputees mostly without a prosthesis, their decreased AD in CC can be interpreted as a kind of “aberrant” plasticity.
Our results suggest a possible axonal degeneration in white matter fibers that connect interhemispheric motor areas after upper-limb amputation.
It should be noted that the microstructural change in CC subregion III was likely dependent on the amputation side. Only the right-side amputees who lost their dominant hand showed decreased AD and FA in the subregion III compared with the healthy controls. This is reasonable as the dominant-side amputation would affect basic motor functions and daily life activities more extensively than the nondominant-side amputation (Yancosek and Mullineaux, 2011). The left-side amputees could still rely on their dominant (intact) hand as before, while the right-side amputees usually have to use their nondominant (intact) hand more excessively to compensate for the loss of the dominant hand. Brain reorganization after amputation seems use-dependent. Using the same DTI images, we found more reorganization of whole-brain structural network in the dominant-side amputees compared with the nondominant-side amputees (Guo et al, 2019a).
Previous fMRI studies found that amputees who relied more on their intact limb showed more profound expansion of activation from the intact cortex to the deprived cortex (Makin et al, 2013a). Such use-dependent reorganization has been suggested to be related to the decreased IHI (Werhahn et al, 2002). However, the nonsignificant differences between the left-side amputees and healthy controls might be due to the small sample size of the left-side amputees (n = 7), since the effect size was large for AD in subregion III (Cohen's d = −1.331).
Pain experience is another important factor for the microstructural change of CC after upper-limb amputation. Only the amputees with RLP showed decreased FA and AD in CC, which was also correlated with pain intensity. Several previous studies have suggested the association between chronic pain and CC microstructural change (Geha et al, 2008; Kim et al, 2014; Seo et al, 2019). Decreased FA in CC has been observed in chronic complex regional pain syndrome and fibromyalgia (Geha et al, 2008; Kim et al, 2014). Alterations in AD and RD have been suggested to be associated with PLP after upper-limb amputation (Seo et al, 2019). PLP has been believed to arise from maladaptive reorganization of the cortex after amputation (Flor et al, 2006); however, the etiology of RLP has mostly been attributed to peripheral pathological changes of the stump (Jensen et al, 1983; Ramanavarapu and Simopoulos, 2008) rather than brain reorganization.
Our study suggests that RLP may also play an important role in interhemispheric structural connectivity reorganization. Moreover, compared with the study of PLP, which only compared amputees with PLP and healthy controls (Seo et al, 2019), and thus could not distinguish the influences from pain and amputation, our study also recruited amputees without RLP. Both p-value and effect size suggested that the CC metrics of the amputees without RLP were more likely similar with healthy controls rather than with the amputees with RLP, implying that pain experience and interhemispheric structural connection reorganization are highly related.
This speculation was also supported by some pain-specific alterations only observed in the amputees with RLP, but not in all amputees. First, in addition to the microstructural changes in the CC subregion II and III, the amputees with RLP also showed AD reduction in the CC subregion IV, which structurally connects the bilateral S1. Persistent pain following loss of sensory input has been suggested to be associated with deterioration in the structure and function of the sensory cortex (Makin et al, 2013b; Zheng et al, 2021). Our previous study also correlated RLP with metabolite concentration and BOLD activity in the sensory cortex (Guo et al, 2019b). Taking account of pain-related reorganization in the sensory cortex, we believe the white matter that connects the bilateral sensory cortex could also be affected by pain.
Second, in addition to AD reduction, FA was also found decreased in the amputees with RLP. FA is a highly sensitive indicator of microstructure integrity and it is not very specific to the type of change (Alexander et al, 2011). FA of CC was reported to be influenced by several different microstructural properties such as myelination and axon density (Friedrich et al, 2020). Furthermore, the amputees with RLP had a lower AD in CC subregion III and IV than the amputees without RLP, which further demonstrated the impact of RLP on AD. Therefore, our results imply more degeneration of interhemispheric white matter connections in the amputees with RLP.
Generally, FC estimated using correlation has a higher test–retest reliability, fingerprinting accuracy, and system identifiability compared with partial correlation, coherence, and information theory-based measure. Using this method, there were no significant changes in interhemispheric FC after upper-limb amputation. Our results of FC are inconsistent with a previous study that reported reduced interhemispheric FC in sensorimotor network after upper-limb amputation (Makin et al, 2013b), which could be due to the PLP. Most of the amputees (16/18) in Makin et al (2013b) reported PLP, while only 3 amputees (3/22) in this study had PLP. Makin et al (2013b) have revealed that the magnitude of PLP is positively correlated with reduced interhemispheric FC. Although literature has reported the consistency of structural and FC (Greicius et al, 2009; Skudlarski et al, 2008), there were also studies pointing out that FC was not entirely dependent on structural connectivity.
A study combing fMRI and DTI showed that the structural connected regions showed a high level of FC, but the reverse was not necessarily true, as FC may also be mediated indirectly through the third gray matter region (Koch et al, 2002). Clinical case studies reported that FC could be preserved after disruption of white matter structures (Johnston et al, 2008; Uddin et al, 2008). For example, a patient with complete commissurotomy showed comparable interhemispheric FC with healthy controls (Uddin et al, 2008); another case with complete section of CC preserved interhemispheric FC in the sensorimotor cortex (Johnston et al, 2008). All the preserved interhemispheric FC was postulated to be coordinated by other white matter connections. Similarly, we speculated that upper-limb amputees preserved the interhemispheric FC by compensation in other regions, such as the anterior and posterior parts of the CC.
Our results suggest that if PLP was absent, the interhemispheric FC could be preserved even if the structural connections have been disrupted after upper-limb amputation.
In addition to the changes in interhemispheric connectivity, upper-limb amputation may also lead to intrahemispheric changes. Although whole-brain DTI analysis in lower-limb amputees observed prominent changes in CC (Simões et al, 2012), another study on upper-limb immobilization still found a significant change of corticospinal tract in the affected hemisphere (Langer et al, 2012). However, in our sample of upper-limb amputees, no significant changes of corticospinal tract were observed. These results suggested that unilateral upper-limb amputation mainly induced degeneration of interhemispheric motor pathway rather than intrahemispheric ones, as a result of long-term underuse of the amputated limb and overuse of the intact limb.
In summary, the current neuroimaging study revealed a degeneration of interhemispheric structural connectivity after upper-limb amputation, unveiling the underlying structural basis for bilateral functional reorganization after amputation. The amputation side and RLP might be two confounding factors for structural degeneration. However, the small sample size may limit the generalization of the present results, especially those obtained from subgroup comparisons (9 RLP amputees and 13 non-RLP amputees; 15 dominant-side amputees and 7 nondominant-side amputees). Further studies with larger samples are needed to demonstrate the effects of these factors.
Footnotes
Authors' Contributions
K.Z.: Methodology; software (lead); data analysis (lead); and writing—original draft (lead). C.W.: Methodology (lead) and software (supporting). Y.L.: Conceptualization (lead) and data curation (lead). J.X.: Investigation (lead) and data curation. C.P.: Data curation (supporting) and conceptualization. X.G.: Writing—original draft (supporting) and writing—review and editing (equal). S.T.: Validation (lead) and writing—review and editing (equal).
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Basic Research Program of China (973 Program) (No. 2011CB013304) and the National Natural Science Foundation of China (No. 61771313).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.