
Abstract
Background:
To date, numerous studies have documented various alterations in resting brain activity in Alzheimer's disease (AD) and other neuropsychiatric diseases. In particular, disease-related alterations of functional connectivity (FC) in the resting state networks (RSN) have been documented. Altered FC in RSN is useful not only for interpreting the phenotype of diseases but also for diagnosing the diseases. More recently, several studies proposed the dynamics of resting-brain activity as a useful marker for detecting altered RSNs related to AD and other diseases.
Objectives:
In this article, we review recent studies exploring alterations of static and dynamic functional connectivity in AD and other neuropsychiatric diseases. We then discuss how to utilize and interpret dynamics of FC for studying resting brain activity in diseases.
Results:
In contrast to previous studies, which focused on FC calculated using an entire fMRI scan (static FC), newer studies focused on the temporal dynamics of FC within the scan (dynamic FC) to provide more sensitive measures to characterize RSNs. However, despite the increasing popularity of dynamic FC, several statistical investigations of dynamic FC cautioned that the results obtained in commonly used analyses for dynamic FC require careful interpretation.
Conclusions:
Although static and dynamic FC are likely to be a useful tool to detect altered RSN in patients affected by AD and other neuropsychiatric disorders, interpretation of altered dynamic FC in patients require special care.
Impact statement
We review recent studies of static and dynamic functional connectivity (dFC) in Alzheimer's disease and discuss interpretation of dFC.
Alteration of Static Functional Connectivity in Alzheimer's Disease
Alzheimer's disease (AD) is the most common neurodegenerative disease, which causes numerous cognitive dysfunctions including recent memory loss. Functional neuroimaging studies using positron emission tomography (PET) and single photon emission computed tomography (SPECT) in AD patients have shown hypoperfusion in the temporal lobe, medial (posterior cingulate cortex [PCC] and precuneus) and lateral parts (superior parietal lobule and inferior parietal lobule) of the parietal lobe, which are considered to belong to the default mode network (DMN; Fig. 1) (Burns et al, 1989; Neary et al, 1987).
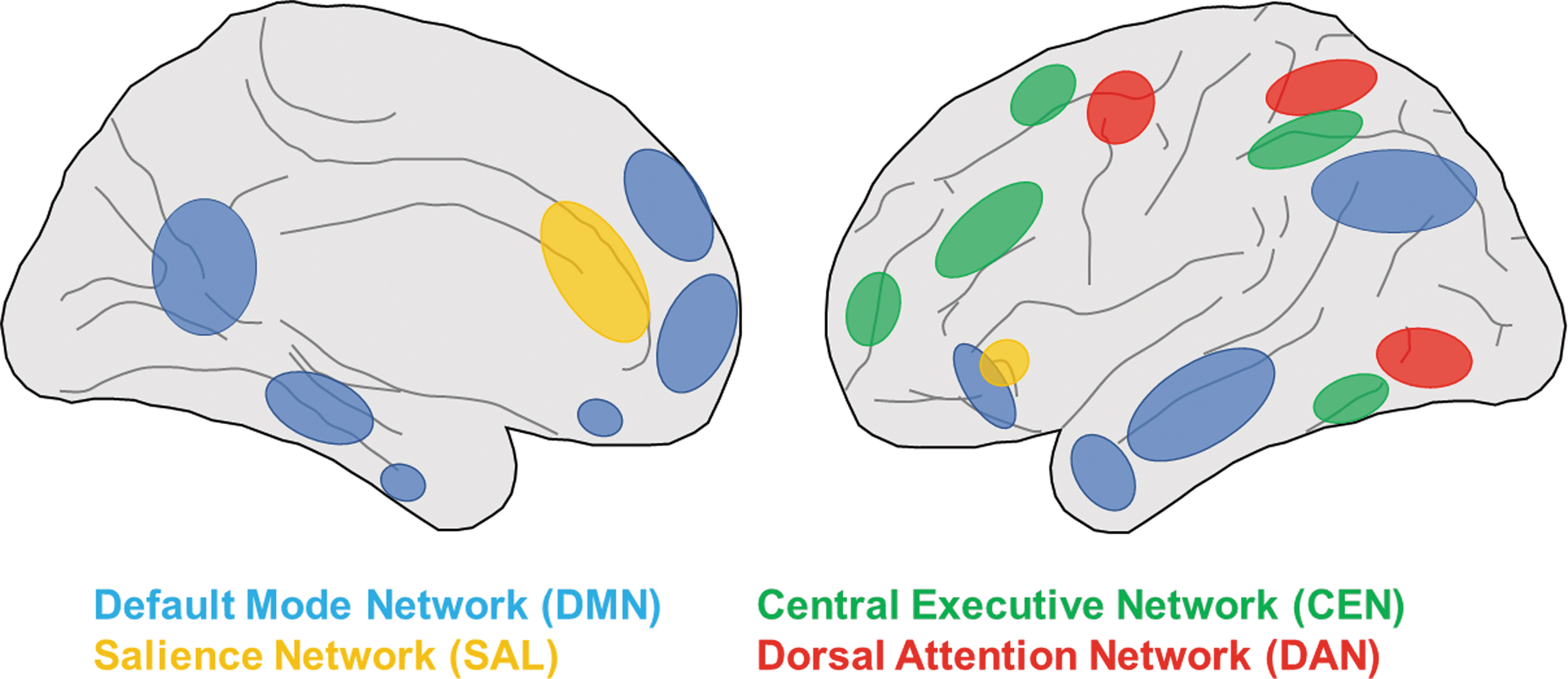
RSNs altered in AD. Cortical nodes of DMN (blue), SAL (yellow), CEN (green) and DAN (red) are shown. Left and right show lateral and medial view of the human brain, respectively. AD, Alzheimer's disease; CEN, central executive network; DAN, dorsal attention network; DMN, default mode network; RSNs, resting state networks; SAL, salience network.
DMN also showed task-related hypoactivation during cognitive performance in functional magnetic resonance imaging (fMRI) studies of AD patients (Buckner et al, 2008). A study using PiB (PET ligand of β-amyloid) reported β-amyloid deposition in the region of DMN with impaired activity (Sperling et al, 2009), indicating neural dysfunction in the core regions of DMN associated with β-amyloid deposition. Moreover, fMRI activity in the precuneus/PCC exhibited a statistically significant reduction of deactivation in mild cognitive impairment patients compared with cognitively normal aged individuals (Sperling et al, 2009). Thus, these studies collectively indicate altered DMN activity in AD patients.
Several studies found impairment of global brain network activity, including DMN, and its correlation with disease severity in AD patients (Table 1). Using fMRI, Greicius et al (2004) reported decreased resting state activity during cognitive tasks in the PCC and hippocampus of AD patients compared with normal subjects, indicating that functional connectivity (FC) between the two regions of DMN is impaired in AD. Consistently, graph theoretic analysis of resting state fMRI (rs-fMRI) data in AD patients revealed a loss of small-world properties in the brain networks, suggesting the dysfunction of resting state networks (RSNs) in AD patients (Supekar et al, 2008). Moreover, Shultz and colleagues reported abnormality within the DMN increased as the score of Clinical Dementia Ratings, which measures AD severity, advanced (Brier et al, 2012). The study also reported dysfunction of FC between RSNs, such as DMN salience network (SAL) and DMN dorsal attention network (DAN), in AD patients.
List of Studies Reporting Altered Functional Connectivity/Dynamic Functional Connectivity in Alzheimer's Disease and Other Neuropsychiatric Disorders
AD, Alzheimer's disease; ADHD, attention deficit hyperactivity disorder; ASD, autism spectrum disorder; CEN, central executive network; DAN, dorsal attention network; DMN, default mode network; FC, functional connectivity; FPN, frontoparietal network; M-CIN, midcingulo-insular network; PD, Parkinson's disease; RSNs, resting state networks; SAL, salience network; SMN, somatomotor network.
Recent studies further revealed the correlation between impaired FC and various cognitive dysfunctions in AD patients, such as impairments in memory, orientation, and executive functions (Buckner and DiNicola, 2019). We recently reported a relationship in AD between impaired resting state FC and disorientation in time (Yamashita et al, 2019b; Disorientation in time refers to a condition in which a person loses track of the exact day, date, or year). The difference of resting state FC in groups of AD patients with good versus poor orientation in time (OT) revealed a significantly stronger FC between PCC and the right middle temporal gyrus (MTG) proximal to the temporoparietal junction, suggesting that disconnection between PCC and the right ventral attention network including MTG may cause OT disturbance in AD patients. In a similar study, He et al (2014) reported a potential link between the central executive network (CEN) and reduced executive function in AD.
In addition to impairments of FC within each RSN (e.g., DMN, DAN), FC between RSNs is also impaired in AD patients. Wang et al (2019) reported that DMN and DAN showed decreased anticorrelation in AD patients suggesting impaired inter-RSN FC. Recently, we examined RSNs in AD patients treated with acetylcholinesterase inhibitors (AChEIs) (Yamashita et al, 2022). AChEIs have been used as a pharmacological treatment for mild-to-moderate moderate AD, and short-term improvements in cognitive functions and associated changes in FC of restricted brain areas have been reported (Griffanti et al, 2016). By comparing the resting state FC of AChEI-responsive and nonresponsive AD patients, we found that the reduction because of AChEI of the abnormal increase in FC between DAN and the visual attention network was paralleled with the amelioration of OT. This suggested close association between inter-RSN FC and higher cognitive functions affected by AD (Yamashita et al, 2022).
Alteration of Static FC in Neuropsychiatric Diseases
In addition to AD, recent rs-fMRI studies revealed a link between impairment of RSNs and other neuropsychiatric disorders. In Parkinson's disease (PD), seed-based analysis of rs-fMRI data revealed increased FC between the cerebellum and somatomotor network (SMN) in parallel with decreased FC within the SAL compared with the controls (Tuovinen et al, 2018). Furthermore, severity of motor symptoms of PD as scaled by Unified Parkinson's disease rating scale three (UPDRS-III) was associated with hypoconnectivity from SMN (Tuovinen et al, 2018). A decrease of intra-FC in DMN was also reported in PD patients with cognitive impairment (Gorges et al, 2015).
In epilepsy, although a SPECT study showed increased perfusion of the seizure focus, patients with a generalized seizure or complex partial seizure showed decrease of activation in DMN areas during the ictal state (Blumenfeld et al, 2004). Based on these observations from the group of patients with surgically proven mesial temporal lobe epilepsy, the network inhibition hypothesis postulates that seizure produces a polyspike discharge that propagates from the temporal lobe to subcortical structures as well as bilateral frontoparietal association cortex. This polyspike discharge suppresses activity in these areas, including DMN, and leads to a loss of consciousness (Blumenfeld, 2012).
Associations between various psychiatric diseases and RSNs dysfunctions are also reported. Greicius et al (2007) reported increased FC of DMN in depression patients compared with healthy subjects using independent component analysis (ICA) and indicated that the altered activity in DMN was associated with the ruminative nature of depression. Concurrent with the hyperconnectivity within DMN, impaired connectivity between DMN and CEN was also observed. The decrease in inter-RSN FC may explain the difficulties of depressed patients to disengage the processing of self-focused, often negatively biased thoughts. Of interest, in contrast to DMN and CEN, increased internetwork connectivity between DMN and SAL was reported (Manoliu et al, 2013). The increased FC between DMN and SAL may be linked to depression events as well as behavioral responses to internal and external stimuli.
Although still controversial, previous studies have reported a potential link between schizophrenia and altered RSNs. Some studies indicated that impaired SAL that induces insufficient suppression of DMN and a decrease in auto-excitation of CEN may cause positive psychosis (Looijestijn et al, 2015). Similarly, Tu et al (2013) used brainwise analysis of rs-fMRI data to show that patients with schizophrenia exhibited larger FC between the frontoparietal network (FPN) and DMN and other RSNs. This hyperconnectivity in schizophrenia may be one of the mechanisms of the cognitive and perceptual symptoms.
Finally, the link between DMN dysfunction in developmental disorders was elucidated by recent studies. A study using visual-evoked potentials, event-related potentials, and DTI reported that abnormal visual perception associated with autism spectrum disorders (ASDs) was related to abnormality in anatomical connectivity between parallel visual pathways and attention networks (Yamasaki et al, 2017). Furthermore, an fMRI study that reported coupling between activated DMN and deactivated FPN proposed the DMN interference hypothesis in patients with attention deficit and hyperactivity disorders (Sonuga-Barke and Castellanos, 2007).
Alteration of Dynamic Functional Connectivity in AD and Neuropsychiatric Diseases
Recently, nonstationary aspects of resting brain activity have been increasingly used to characterize RSNs with higher sensitivity compared with static FC (Calhoun et al, 2014; Hutchison et al, 2013; Matsui et al, 2019; Mitra et al, 2018; Preti et al, 2016; Shine et al, 2016). In this dynamic view of resting brain activity, it is assumed that the resting brain switches among multiple states each of which is characterized by distinct patterns of coordinated activity among brain regions. The switching among different states produces a moment-to-moment fluctuation of the spatial patterns of RSNs that is often referred to as dynamic functional connectivity (dFC).
At present, the two most widely used methods to analyze dFC are sliding window correlation analysis and coactivation pattern (CAP) analysis (Allen et al, 2014; Karahanoğlu and Van De Ville, 2015; Liu and Duyn, 2013; Liu et al, 2013; Fig. 2). The idea of the sliding window correlation analysis is to calculate FC within a short sliding window (∼40 sec) to track the temporal evolution of RSNs (Allen et al, 2014). CAP analysis uses point process analysis of an activity time course in a selected brain area to detect momentary RSN activity at single volume level (Liu and Duyn, 2013). In both methods, the extracted momentary patterns of RSN activity, even those extracted in the same scanning run, are diverse and can be clustered into distinct modules. Because of such distinct activity patterns, these modules extracted by the sliding window correlation analysis and CAP analysis are often regarded as specific states that comprise dFC (Liu et al, 2018; Preti et al, 2016).
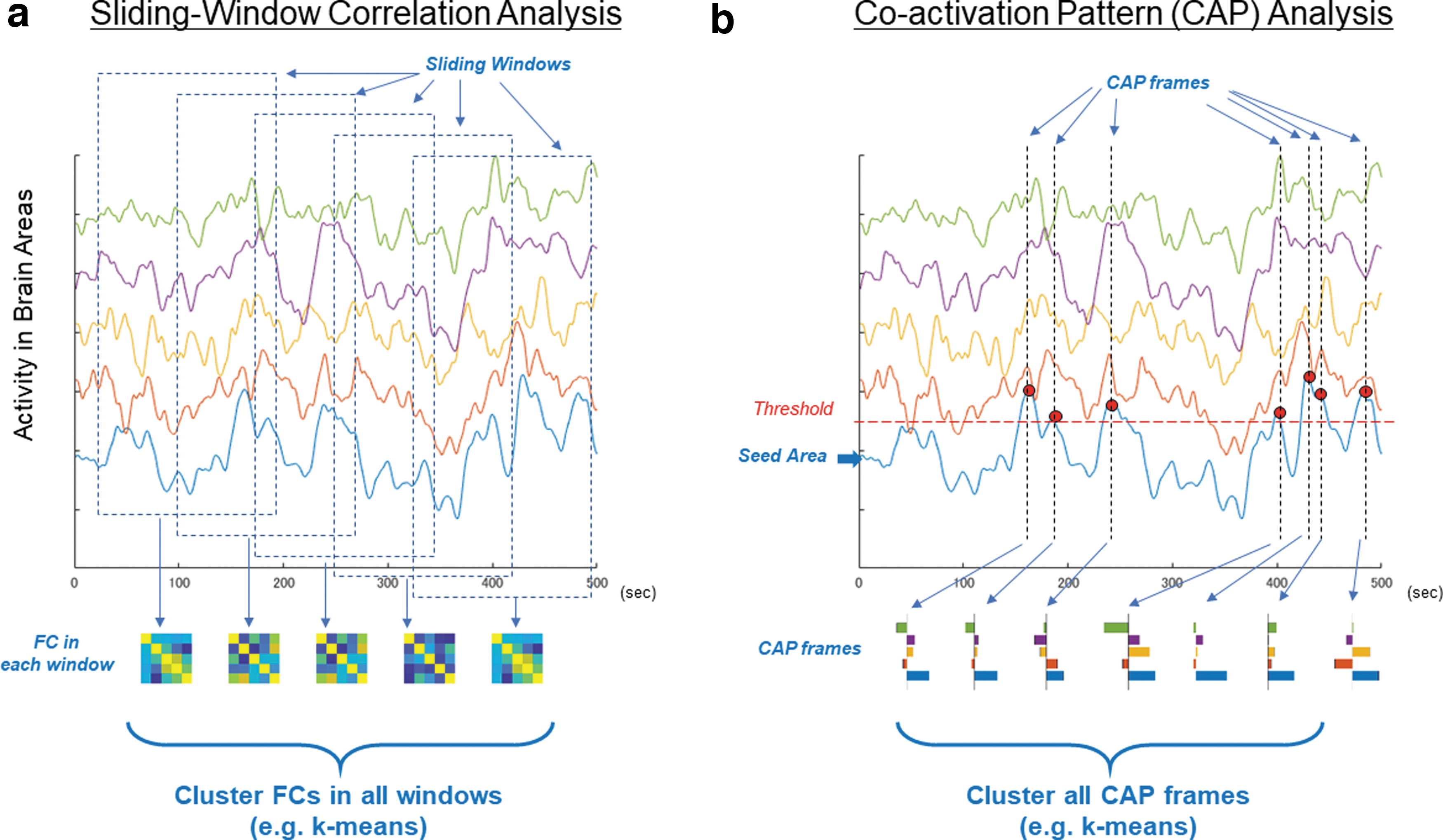
Methods for analyzing dFC.
Several studies demonstrated that dFC can be used to distinguish between patients and healthy controls. A dFC study using the sliding window procedure reported that AD patients spent less dwell time in brain states (i.e., time spent in the brain states) with strong posterior DMN contributions and more dwell time in brain states with strong anterior DMN contributions relative to controls (Jones et al, 2012). Altered dFC in AD patients suggests that oscillatory patterns are progressively altered over the AD staging, eventually converging to a shrinkage of the dynamic repertoire (Córdova-Palomera et al, 2017). Similarly, dFC analysis in PD patients reported the presence of two characteristic states: a more frequent and strongly segregated “State I” and a less frequent, more integrated “State II” (Kim et al, 2017). PD patients exhibited a significant decrease of dwell time in State I along with a proportional increase of dwell time in State II compared with healthy subjects. Of importance, the increase of dwell time in State II was correlated with the severity of motor symptoms, indicating that the loss of functional segregation between brain networks might represent the pathogenesis of PD. These studies suggest that dFC is indeed useful to characterize the resting brain activity in patients affected by AD and other neuropsychiatric disorders.
Other recent studies demonstrated that CAP analysis can discover altered dFC in patients affected by other neuropsychiatric disorders. Marshall et al (2020) used CAP analysis to investigate the salience/midcingulo-insular network (M-CIN) in children with ASD. They found that coactivations of M-CIN and FPN or DMN were less frequent in children with ASD compared with typically developing children. Similarly, less frequent DMN coactivations were reported in AD patients (Ma et al, 2020). In contrast, in schizophrenic patients, coactivations of DMN and SAL were more frequent than in healthy controls (Yang et al, 2021). Thus, these studies suggest that dFC is a promising approach for characterizing resting activity of brains affected by AD and neuropsychiatric disorders.
dFC Requires Careful Interpretation and Further Investigations
Of importance, however, the characterizations of dFC with the sliding window correlation analysis or CAP analysis need careful interpretations (Handwerker et al, 2012; Hindriks et al, 2016; Liégeois et al, 2017; Lurie et al, 2020; Matsui et al, 2022; Zalesky and Breakspear, 2015). It has been shown by the previous studies that the distinct states of resting brain activity discovered by sliding window correlation or CAP analyses merely reflect statistical sampling error, which were reproduced by using simulated data produced by null models that are stationary by construction (Fig. 3a; Cifer et al, 2017; Laumann et al, 2016; Matsui et al, 2022; Novelli and Razi, 2022). Recently, we showed that the spatial patterns of CAPs were closely replicated by stationary Gaussian noise that retained only the covariance structure (i.e., static FC; Matsui et al, 2022). We further showed that, by using only a first-order autoregressive model, the transition patterns among CAPs could also be replicated from a stationary null model. These findings were consistent with a previous study focusing the sliding window correlation method (Liégeois et al, 2017). These studies suggest that aspects of dFC, such as the spatial patterns of CAPs and transition patterns among them, can almost be fully explained by a stationary model that takes into account only static FC and first-order autoregressive dynamics (Fig. 3b). Thus, to be on the safe side, one should avoid interpretation of results obtained by the sliding window correlation analysis and CAP analysis that assumes nonstationarity of dFC (i.e., switching among distinct network states).
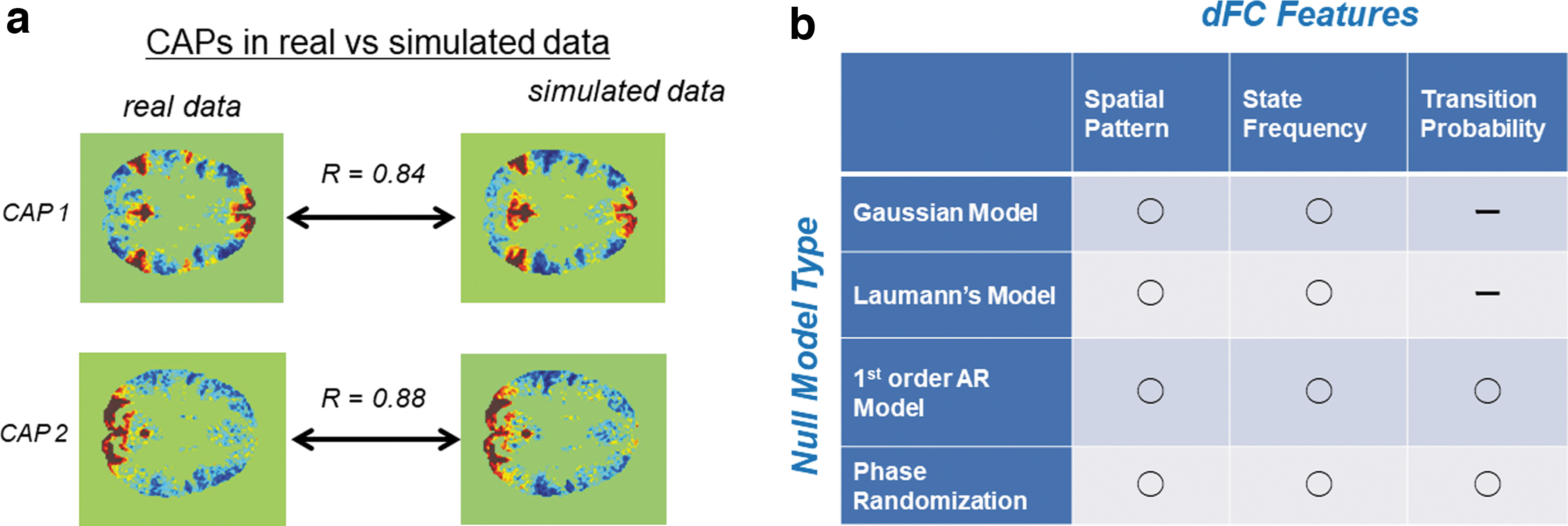
Null models preserve many features of dFC.
We would like to note that these studies did not deny the potential clinical usefulness of the sliding window correlation analysis and CAP analysis as described previously. Although these analysis methods do not provide the proof of nonstationary resting brain activity, it does not automatically mean that the analysis methods do not have usefulness or clinical relevance (Liégeois et al, 2021). In other words, dFC as characterized at present by the sliding window correlation and CAP analyses needs to be interpreted with great care. In some cases, transition patterns among CAP modules may be useful to distinguishing between patients with some neuropsychiatric disorders and healthy controls. Even in such a case, one should still be cautious to avoid interpretations that assume distinct network dynamics in patients and healthy control.
There exist several additional technical issues related to reliability of dFC analysis of AD patients. First, it is well known that head movement during rs-fMRI scanning is harmful to estimation of FC (Power et al, 2012). AD patients may produce larger head movements during scanning that are harmful to rs-fMRI analysis. The problem of movement-related contamination is likely exacerbated in dFC analysis that require high spatiotemporal resolution scanning techniques. Thus, in dFC analysis of AD patients, careful application of cleaning methods for movement-related contamination, such as censoring of volumes with large movements (Power et al, 2014), is essential. Another concern is that the brain-wide association studies (BWAS), in general, require large samples (Marek et al, 2022). Because dFC analysis examines temporal characteristics of the relatively high-frequency band (∼0.02 Hz), BWAS of AD using dFC requires large samples acquired at high spatiotemporal resolution, which is currently unavailable. Nevertheless, there are several reasons to be optimistic that the problem would be solved in the near future. First, high magnetic field MRI scanners equipped with rapid acquisition (Feinberg et al, 2010) is increasingly becoming available and accessible to researchers and clinicians. Second, there is a trend for building large-scale datasets that covers not only healthy participants but also patients with AD or other neuropsychiatric disorders (e.g., UK Biobank). We believe that efforts to push forward these trends would solve the problem of sample size. These trends would be further accelerated by developing techniques that can combine rs-fMRI data collected in diverse locations and scanners into a single large-scale dataset (Yamashita et al, 2019a).
More studies are awaited to bridge the gap between the apparent stationarity of rs-fMRI data and our intuition of nonstationary brain dynamics. One promising avenue is to construct realistic computational models of the resting brain that replicate the features of real fMRI data as quantified by methods such as CAP analysis and sliding window correlation analysis (Wang and Krystal, 2014). Such models would be indispensable for understanding the difference between the resting brain dynamics of healthy individuals and those affected by mental disorders. Another possible approach is to test alternative analysis techniques such as the energy landscape analysis (Ezaki et al, 2017) and temporal ICA (Smith et al, 2012), which are designed to analyze the fine-grained dynamics of resting brain activity. These analysis methods can be tested using the null-model framework as has been performed for the CAP analysis and the sliding window correlation analysis. The use of advanced machine learning techniques may also be useful (Fan et al, 2020; Matsui et al, 2021). Finally, in model animals such as mice, it is now possible to monitor neural activity and hemodynamic activity simultaneously and directly compare the two (Barson et al, 2020; Ma et al, 2016; Matsui et al, 2016, 2018). These new experimental techniques can not only investigate the neuronal activity dynamics in the resting brain of healthy animals but also be applied to mouse models of AD and other neuropsychiatric disorders (Busche et al, 2015). It is hoped that these multidisciplinary approaches will provide a better picture of resting brain dynamics, which would allow the proper interpretation of the results of various dFC analyses.
Footnotes
Authors' Contributions
T.M. and K.i.Y. conceived the study and wrote the article.
Author Disclosure Statement
Authors declare no conflict of interest.
Funding Information
This study was supported by JSPS Kakenhi (JP21H0516513 to T.M., JP20K07888 to K.i.Y.); a grant from JST-PRESTO to T.M.; a grant from Narishige Neuroscience Research Foundation to T.M.