
Abstract
Introduction:
Amyloid-beta (Aβ) pathology is the precipitating histopathological characteristic of Alzheimer's disease (AD). Although the formation of amyloid plaques in human brains is suggested to be a key factor in initiating AD pathogenesis, it is still not fully understood the upstream events that lead to Aβ plaque formation and its metabolism inside the brains.
Methods:
Matrix-assisted laser desorption ionization mass spectrometry imaging (MALDI-MSI) has been successfully introduced to study AD pathology in brain tissue both in AD mouse models and human samples. By using MALDI-MSI, a highly selective deposition of Aβ peptides in AD brains with a variety of cerebral amyloid angiopathy (CAA) involvement was observed.
Results:
MALDI-MSI visualized depositions of shorter peptides in AD brains; Aβ1–36 to Aβ1–39 were quite similarly distributed with Aβ1–40 as a vascular pattern, and deposition of Aβ1–42 and Aβ1–43 was visualized with a distinct senile plaque pattern distributed in parenchyma. Moreover, how MALDI-MSI covered in situ lipidomics of plaque pathology has been reviewed, which is of interest as aberrations in neuronal lipid biochemistry have been implicated in AD pathogenesis.
Discussion:
In this study, we introduce the methodological concepts and challenges of MALDI-MSI for the studies of AD pathogenesis. Diverse Aβ isoforms including various C- and N-terminal truncations in AD and CAA brain tissues will be visualized. Despite the close relationship between vascular and plaque Aβ deposition, the current strategy will define cross talk between neurodegenerative and cerebrovascular processes at the level of Aβ metabolism.
Impact statement
Matrix-assisted laser desorption ionization mass spectrometry-based chemical imaging has been successfully applied to comprehensively delineate spatial Aβ peptide- and neuronal lipid patterns in brains with Alzheimer's disease. This rather new approach overcomes major limitations inherent to commonly used biochemical methods and opens up for both static and dynamic biochemical interrogations of amyloid aggregation in situ.
Introduction
As the world population is living longer, the incidence of age-associated disorders, including dementia and neurological disorders, is increasing dramatically. Therefore, dementia constitutes an ever-increasing major public health burden and currently affecting >50 million people across the world. This number will dramatically rise and to triple by the year 2050 (Alzheimer's Association, 2020). Since Alzheimer highlighted the two key histopathological lesions, senile plaque (SP) and neurofibrillary tangles, more than 110 years ago (Alzheimer, 1907), advances in molecular biology, genetics, as well as image analysis and protein sciences helped us a lot in understanding the pathogenesis of the disease (Masters, 2010). However, no immediate disease modifying or curative therapeutic strategies are available yet (Selkoe and Hardy, 2016), despite some recent success of antiamyloid immunotherapies in early clinical trials such as lecanemab (van Dyck et al., 2023).
Amyloid plaques are precipitating histological features that are found in the cerebral parenchyma and deposited in vessels of Alzheimer's disease (AD) brains long before disease onset (Hardy and Allsop, 1991; Hardy and Higgins, 1992). These histopathological structures of 50–200 μm in size are easily visualized in tissue sections by using either silver impregnation methods or immunohistochemistry (IHC) (Braak and Braak, 1991; Gravina et al., 1995; Ikeda et al., 1989). Amyloid plaques have been identified to primarily consist of a small 4 kDa protein, beta-amyloid peptide (Aβ), secreted upon sequential enzymatic processing of the integral membrane protein, amyloid precursor protein (APP) (Selkoe et al., 1986).
The entire array of cerebral Aβ peptides is a family of hydrophobic, up to 43 amino acid containing peptides with differences in both their N- and C-terminal sequence motifs (Harigaya et al., 2000; Portelius et al., 2010), which in turn posed significant challenges to determine the accurate primary sequence of Aβ in plaques in the first place. This was mainly due to the limitation of previously established techniques for protein and peptide sequencing, mainly the Edman analysis, that complicated accurate sequence identification particularly at the N-terminal part of the peptide. The advent of biological mass spectrometry (MS) techniques has helped to significantly advance our understating of Aβ plaque chemistry.
Since the 1990s, MS has become the major method for protein sequence identification. In this study, the development of two soft ionization methods, including matrix-assisted laser desorption ionization (MALDI) (Karas and Hillenkamp, 1988) and electrospray ionization (ESI) (Fenn et al., 1989), was critical as this allowed for rapid and accurate protein sequence analysis. These developments fueled the development of an entire new research field, proteomics, which comprises large-scale protein sequence identification and quantification in biological samples (Tyers and Mann, 2003). Similarly, MS has further gained prominence in large-scale profiling of other biomolecules including metabolites and lipids.
Proteomic analyses targeting senile plaque (SP) and tangles in AD, Lewy body in Parkinson's disease (PD), as well as polyglutamine aggregates in polyQ diseases' brains have been extensively performed in combination with a robust biochemical purification step of the target structures and molecules. In the earlier studies, Allsop et al. (1983) were the first to isolate cores from neuritic plaques from postmortem AD brains that were insoluble in various denaturant agents. Subsequently, Masters et al. (1985) have succeeded in purifying and characterizing amyloid plaque cores (APCs) from the brains of elderly Down syndrome (DS) patients with AD. Since APCs are characterized by an unusual solubility profile, they compared and utilized the most effective solvents to achieve complete and rapid solubilization of APCs (Masters et al., 1985; Mukherjee et al., 2021).
From the early era of their discovery, diversity of Aβ proteoforms in AD brains has been extensively studied through immunocytochemical methods (Iwatsubo et al., 1996; Iwatsubo et al., 1994) as well as biochemical analysis as above mentioned. Especially, Aβ extracts from neuritic plaques and blood vessel walls in AD brain tissue showed Aβ peptide patterns with extensive N- and C-terminal modifications.
Pyroglutamation of 3-Glu (3pE-Aβ) is the predominant N-terminal modification of Aβ species. These were primary detected in the AD cerebral cortex extracts (Mori et al., 1992) and were found to be localized in diffuse plaques in AD and DS brain tissues much earlier than the full-length peptide (Saido et al., 1995). In addition, 3pE-Aβ showed faster aggregation kinetics, higher stability, and were found to be more neurotoxic than the untruncated Aβ isoforms (D'Arrigo et al., 2009; Wirths et al., 2010) and might be better targets for therapeutic interventions (Bayer and Wirths, 2014).
Whereas proteomic and lipidomic approaches for tissue extracts have been developed for the sensitive identification and quantitation of each protein/lipid so far, no distinct spatial information of those species in the corresponding part of the tissue was obtained. Especially, due to the cellular and histological complexity of central nervous system (CNS) tissues, the spatial information of CNS-related biomolecules is of major interest for the delineation of neuronal mechanisms.
Biochemical imaging tools are of essential relevance for delineating biomolecular localizations and processes at the subcellular level. To obtain spatial chemical information in vitro and in situ, the most commonly used bioimaging techniques include IHC, in situ hybridization, proximity ligation, and chemical staining together with fluorescent microscopy as well as spectroscopic methods such as infrared spectroscopy, Raman scattering (RS) microscopy, and coherent anti-Stokes RS.
Bioimaging strategy has been developed to achieve high sensitivity and high resolution at the subcellular and cellular levels for biological samples. As for specificity, this technology is particularly valid for imaging of endogenous lipid species, as there are no conventional biological imaging strategies for protein (IHC) or nucleotide (polymerase chain reaction, fluorescence in situ hybridization) detection available. In situ detection of proteins and lipids is also possible by a rather new imaging modality, mass spectrometry imaging (MSI), that makes use of the advantages of MS allowing chemically specific molecular imaging in tissues and cells.
Mass Spectrometry Imaging
Since its development in the mid-1990s, MSI has emerged as a powerful competitor to established techniques for chemical imaging of proteins, peptides, lipoids, and metabolites in cells and tissues (Andersson et al., 2008; Caprioli et al., 1997; Hanrieder et al., 2011; Hanrieder 2013a, Hanrieder et al., 2013b; Hanrieder 2015; McDonnell and Heeren, 2007; Schwartz and Caprioli, 2003; Seeley and Caprioli, 2008; Yang and Caprioli, 2011). MSI enables a label-free molecular imaging allowing an untargeted detection of a broad range of biomolecules. However, still it needs technological advancements to target relatively large molecules such as proteins and peptides, especially aggregation-prone characteristics.
MSI comprises different modalities that are characterized by the different probes used to desorb, transfer, and ionize different biomolecules from a tissue section or cell preparation into the mass spectrometer (Fig. 1). The most commonly used approach is MALDI, where a tissue section is covered with an organic, crystalline compound that has ultraviolet light absorbing properties, and molecular analytes are desorbed and ionized through irradiating the sample with a laser (Karas and Hillenkamp, 1988). Alongside MALDI, other prominent MSI approaches include time-of-flight (ToF) secondary ion mass spectrometry (SIMS) (Vickerman, 2011), where molecular species are sputtered into the gas phase using a highly focused primary ion beam. In another MSI modality, desorption ESI (Takátss et al., 2004), an electrospray is focused onto a tissue sample to achieve desorption and ionization.

Principle of MALDI-MSI.
Generally, in an MSI experiment, imaging is performed by acquiring mass spectra sequentially from x,y array that in essence defines the pixel resolution. Ion maps are generated by visualizing the ion intensity for a specific biomolecule within the pixel array that has been analyzed by MSI (Fig. 1).
The various MSI technologies are characterized by different advantages and limitations that typically balance spatial resolution and molecular information. MALDI-MSI provides a superior molecular mass range, that is, molecular coverage, with fair spatial resolution, good sensitivity, and high-molecular specificity. Most importantly, MALDI is particularly well suited for imaging of low-molecular weight (200–1000 Da) to small proteins (<30 kDa) with 1–10 μm lateral resolution (Kompauer et al., 2017; Spengler and Hubert, 2002).
This allows for direct correlation of proteins and peptide signals with histopathological features revealed through consecutive histological and immunostaining analyses performed on the same section (Chaurand et al., 2004; Kaya et al., 2017c; Michno et al., 2018; multiplexed imaging of both proteins and peptides as well as lipid species within the same tissue (Kaya et al., 2017a); and finally, tomographic imaging through 3D reconstruction of sequentially collected and acquired 2D tissue data enabling 3D spatial segmentation (Andersson et al., 2008; Jones et al., 2012) and correlation of MSI data with in vivo imaging techniques including magnetic resonance imaging and positron emission tomography (Abdelmoula et al., 2019; Henderson et al., 2018).
Herein we aim to provide a summary of MALDI-MSI for protein, peptide, and lipid imaging in brain tissue.
Further different advances as well as the challenges of various parameters defining optimal MALDI-MSI performance are all the aspects of sample preparation, matrix application, data acquisition and validation, as well as statistical analysis. An overview of MSI-based applications with focus on protein and peptide as well as lipid imaging in AD pathology is provided.
Experimental Considerations for MALDI-MSI of Plaque Pathology
Sample preparation
Appropriate sample preparation is a critical part within an MALDI-MSI experiment, to preserve the histomorphology of the samples and the biochemical state of the tissue to minimalize the risk of sample contamination and of postmortem changes such as molecular degradation. MALDI-MSI of both endogenous peptides and lipids is primarily performed on fresh frozen brain tissue, as embedding and fixation media introduce crosslinking artifacts and polymers that interfere with the MS ionization process and compromise the analyte signal. Tissue cryosections are collected and washed with physiological, volatile buffers such as ammonium bicarbonate for sample cleanup. In the case of peptide analysis, organic washes are performed for lipid removal and peptide fixation in case of amyloid peptide analysis.
A further critical step for Aβ imaging is to treat tissue sections with formic acid (FA) for dissolving amyloid aggregates in situ and to enhance the Aβ peptide signals. Vaporization of FA is required to preserve tissue integrity (Ikegawa et al., 2019).
A further source of consideration is matrix application. In MALDI, the matrix layer absorbs the energy from the irradiated laser and induces desorption and ionization of analytes. It is a well known fact that homogeneity of matrix crystal deposition is crucial for high sensitivity, reproducibility, and artifact-free imaging. There are automatic pneumatic sprayer systems being introduced for reproducible and robust matrix application yielding a highly preserved spatial resolution with a good reproducibility.
Finally, the choice of the proper matrix is crucial to ensure homogeneity of crystallization and efficient ionization of the analytes. A number of matrix compounds have been used for MALDI-MSI of various molecular species, including different lipids varying in polarity and hydrophobicity. The most commonly used matrices for peptide/protein analysis include α-cyano-4-hydroxycinnamic acid (CHCA), 2,5-dihydroxybenzoic acid (2,5-DHB), 2,6-dihydroxy acetophenone (2,6 DHA), and sinapinic acid. For Aβ imaging, currently 2,6 DHA was demonstrated to give the best signal quality, particularly in human tissue (Michno et al., 2019a). In contrast, for lipid analysis, the most commonly used matrices are 2,5-DHB, 1,5 di-amino-naphthalene (1,5-DAN), 9-aminoacridine, and CHCA.
Instrumentation and data acquisition
The key parameters for each MALDI imaging experiment are the target species (i.e., lipids, metabolites, peptides) and the spatial resolution. These are primarily dependent on the MALDI instrumentation as well as sample preparation and acquisition modii. The most common MALDI instruments are MALDI ToF setups that warrant sensitive and fast analysis of all kind of analytes over a very wide mass range at a high spatial and mass resolution. Particularly in the case of peptide analysis, MALDI-ToF instruments are superior as they provide the best sensitivity and scan speed along with the highest spatial resolution.
A limitation of ToF instruments can be the mass resolution, which while being very good in general (R = 40,000) are particularly relevant to distinguish and identify lipid isomers and metabolites in situ. This is overcome in high-resolution mass analyzers including Fourier transform ion cyclotron and orbitrap instruments. While those setups provide superior mass resolution for low-molecular-weight analysis, they have limitations with respect to sensitivity and mass range for intact peptide and protein imaging. In addition, those instruments have inferior acquisition speed restricting the acquisition area and/or spatial resolution of the experiment.
Data acquisition involves primarily definition of the acquisition region, spatial resolution, and mass range. In this study, it is important to note that spatial resolution and acquisition area need to be compromised as highly resolved analyses result in long acquisition time. This in turn is limited by the vacuum of the instrument, leading to matrix evaporation over time. The mass range is primarily important when considering the data size.
In addition, it is important to determine the number of single-shot events and laser energy, respectively, that can be obtained from a single pixel resulting in sufficient spectra quality. Using too many shots and or too high laser energy can lead to oversampling and compromises the analyte signal and spatial resolution negatively (Kaya et al., 2017c). This is of particular relevance for MSI of peptides and proteins, where higher laser fluences are needed as those species are less efficiently extracted, ionized, and desorbed.
Furthermore, a large emphasis of the MSI acquisition run needs to be put on acquisition time. This parameter is dependent on spatial resolution, that is, number of pixels, laser frequency, as well as the number of laser shots corresponding to the number of MALDI experiments performed on each pixel. Ultimately, large tissue areas at high resolution will take several hours. This retards a reproducibility of data as the volatile matrix might sublimate away, which will lead to reduced data quality for MSI experiments performed during that phase when the matrix is already partially or totally evaporated.
Molecular identification
A cornerstone and cavity in MSI are the fact that molecular annotation is predominantly performed through intact mass annotation using the accurate mass of the detected peaks along literature or database references as well as with isotope pattern prediction. This requires orthogonal validation typically through histological and immunohistochemical staining as well as tissue identification through tandem mass spectrometry (MS/MS). Below we summarize the most common approaches for analyte validation currently used in MSI.
Immunohistochemistry
Most commonly, IHC is performed after an MSI experiment, preferably of the same tissue sections. However, IHC has limitations with respect to antibody specificity. Furthermore, it is critical to perform IHC toward the same truncated protein isoform, something that can be difficult as there might be no specific antibodies available. The main caveats for IHC with respect to specificity and sensitivity are particularly challenging for the visualization and quantification of endogenous peptides, including beta-amyloid species. Different amyloid species differ in only a few amino acids C-terminally. Consequently, there is a lot of cross-reactivity for this kind of target sequences, significantly hampering the application of IHC for in situ quantification and validation.
MS-based protein and peptide identification
Given the challenges with orthogonal, antibody-based methods, in situ or ex situ MS/MS is the preferred choice for accurate protein and peptide identification. A significant challenge in MSI of proteins is the fact that those data are acquired in a linear mode at low mass resolution rendering accurate mass annotation highly uncertain and speculative. Therefore, MS/MS data are essential for unequivocal identification. Preferably those MS/MS data should be acquired in situ or in extracts of diligently dissected tissue pieces (Andersson et al., 2008; Groseclose et al., 2007; Hanrieder et al., 2013a). Naturally, ex situ analysis restricts the spatial interpretation of MSI data as the protein MS/MS data are not necessarily acquired from the same region.
Therefore, the most desired approach would comprise direct, in situ MS/MS-based identification of peptides and proteins, also termed top-down characterization. In situ MS/MS poses some challenges due to sensitivity issues, sample complexity, and the limited number of MS/MS experiments that can be acquired from a pixel (Kiss et al., 2014; Stauber et al., 2010; Taban et al., 2007).
The most established strategy for identification of unknown, endogenous peptides in tissues is based on tissue homogenization, protein and peptide prefractionation, and finally, liquid-chromatography mass spectrometry (LC–MS/MS)-based sequence identification (Skold et al., 2002; Svensson et al., 2007; Svensson et al., 2003; Yin et al., 2011).
Lipid identification
Lipids are a structurally diverse class of molecules with important biological functions. Challenges are associated with the lack of sufficient molecular fingerprints due to limited information compared with proteins made from 20 amino acids. However, similar to protein and peptide imaging, initial mass peak annotation is based on accurate mass and reference values reported in the literature or on online repositories. A strong advantage in MSI of low-molecular-weight compounds, including lipids, is the resolving power of high-end mass analyzers, in that mass range particular Fourier transform instruments and also reflector ToF analyzers. This in turn facilitates compound identification based on sub ppm mass accuracy as well as the elemental composition deduced by the resolved true isotope pattern. Increased confidence in identification is achieved through follow-up MS/MS or MSn fragmentation analysis and structural characterization.
In contrast to protein identification, this step is, however, more challenging as lipid compounds and consequently their fragmentation patterns are far more diverse. This renders the generation of predictive databases such as those available for protein sequences very challenging. Notable attempts to generate MS/MS spectral libraries have been made to address this issue, but are of course limited due to the vast number of diverse lipid classes and species (Fahy et al., 2009; Sud et al., 2007).
A further significant challenge in lipid characterization, and lipid imaging in particular, is the identification of lipid isomers both structural isomers and also stereoisomers. This is one of the most significant challenges as isomeric lipid species can have entirely different biological functions. Very elegant approaches by in vitro and even in situ derivatization have been proposed (Bednařík et al., 2018; Su et al., 2019; Thomas et al., 2007; Wäldchen et al., 2019). Most notably are the approaches using gas-phase ozonolysis, both in offline lipidomics and most recently also online, within the MSI experiment (Paine et al., 2018). These advances indicate that lipid characterization in vitro and in situ is becoming increasingly robust and routine.
MALDI Imaging of Plaque Pathology in Mouse Models of AD
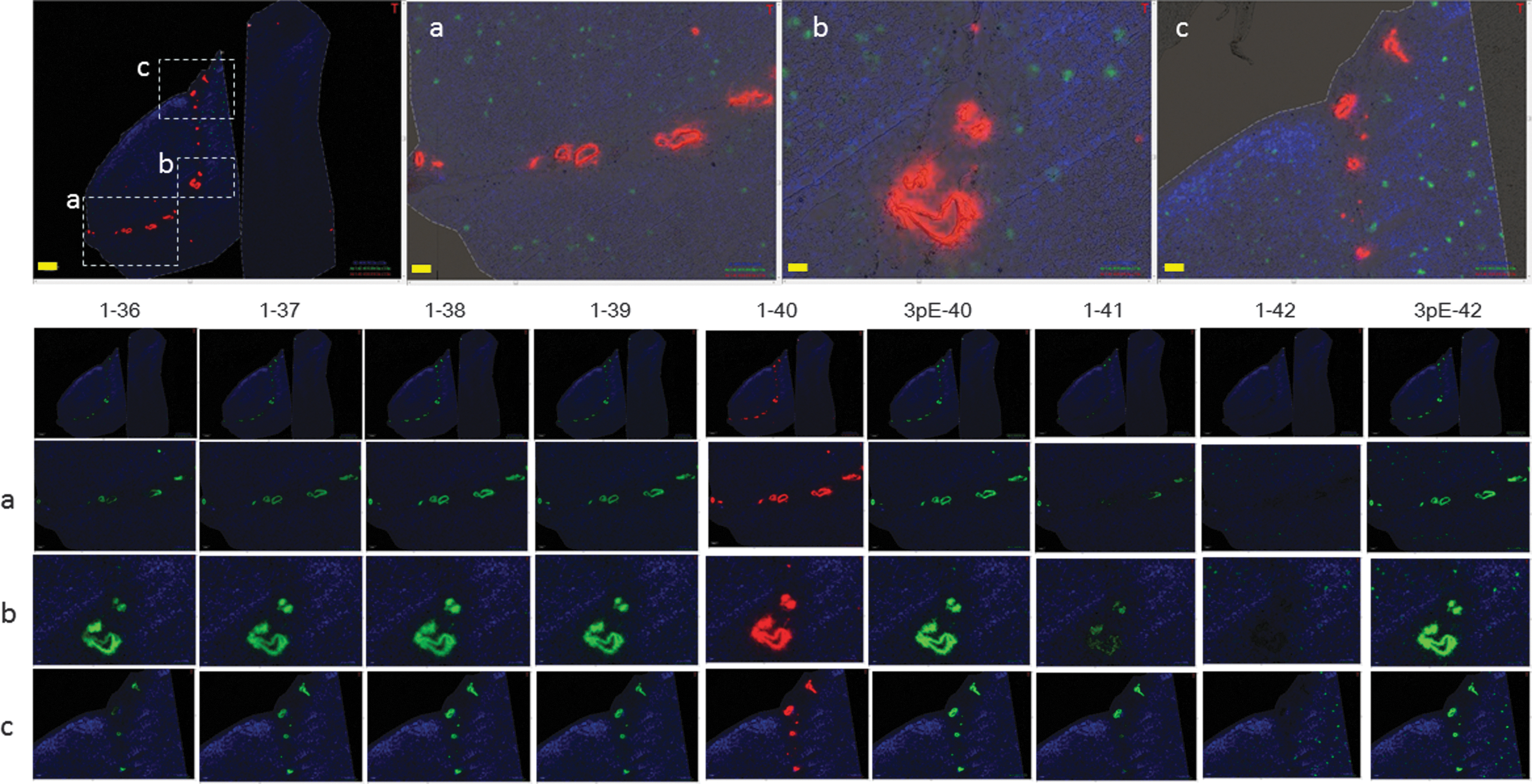
Given its potential for in situ peptide imaging, MSI has consequently been successfully used to delineate single plaque-specific accumulations of various Aβ species in both human tissue and mouse models of AD (Carlred et al., 2016; Kakuda et al., 2017; Kaya et al., 2017a; Kaya et al., 2017b; Michno et al., 2020; Michno et al., 2019a; Michno et al., 2019b; Michno et al., 2019c; Michno et al., 2018; Rohner et al., 2005; Seeley and Caprioli, 2008; Stoeckli et al., 2006). As an early example, Stoeckli et al. (2006) and Stoeckli et al. (2002) have succeeded in visualizing Aβ deposits in a transgenic animal model of AD, APP23. Carlred et al. (2016) used MALDI-MSI to map the Aβ peptide deposition pattern within single plaques in tgArcSwe mice, showing a predominant Aβ1–40 arc and Aβ1–38 arc deposition compared with other mouse models that carry mutations affecting gamma secretase activity, which is well in line with other IHC studies (Lord et al., 2006; Philipson et al., 2009).
Similarly, a predominant Aβ1–40 deposition was found in plaque pathology in tgSwe mice, including both diffuse and compact deposits (Lord et al., 2011; Michno et al., 2019a; Michno et al., 2018). In this study, over time plaque maturation was found to correlate with increased Aβ1–40 deposition, but not Aβ1–42. This suggests a critical role of Aβ1–42 in early plaque formation, while deposition of the less hydrophobic Aβ1–40 is characteristic in plaque maturation into cored deposits. This is relevant as cored plaque formation has been linked to AD pathogenesis, while diffuse plaques are also present in patients who show amyloid pathology, but remained cognitively normal (Michno et al., 2019a).
Probing Plaque Pathology in Human Tissue
Heterogeneity of Aβ proteoforms in parenchymal deposits
Previously, our group reported a protocol for MALDI-MSI targeting both parenchymal and vascular deposits of Aβ of human autopsied brains that was developed (Ikegawa et al., 2019; Kakuda et al., 2017). As anatomic distribution of Aβ plaques is variable and it differs among brain regions in a given person, it is important to select the specimen of the brains in accordance with experimental purposes. In our case, we have analyzed the occipital lobe of the cortex as the initial assessment area of Aβ burden for both parenchymal and vascular deposits in AD and cerebral amyloid angiopathy (CAA) cases. MALDI-MSI analysis identified distributions of full-length Aβ1–42 and Aβ1–40 in deposits in brain tissue (Fig. 2).
MALDI imaging MS in human AD brain tissue.
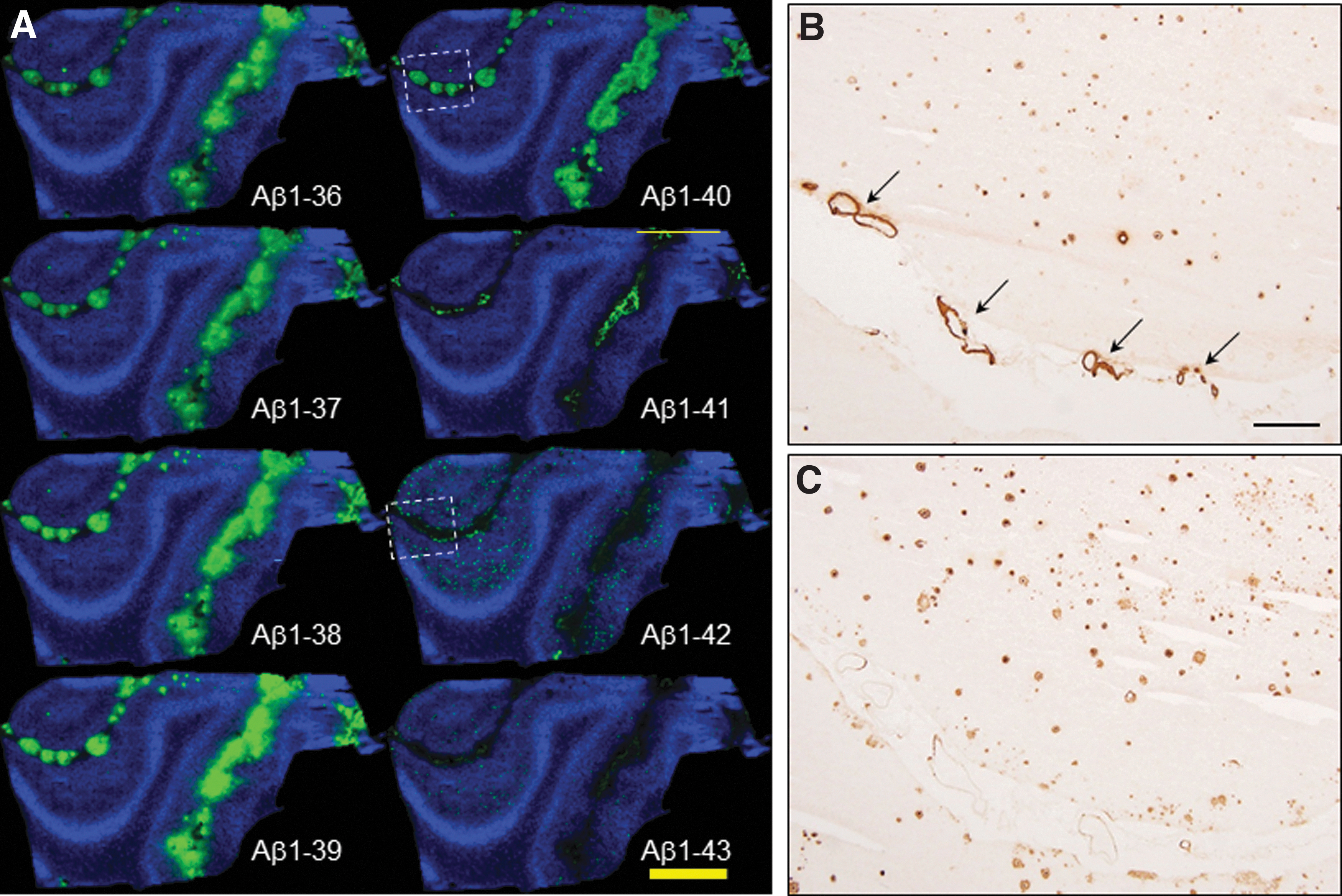
MALDI-MSI revealed predominant cerebral Aβ1–40 distribution in the leptomeningeal vessels of the subarachnoid space and arterioles (Fig. 2). On the contrary, localization of Aβ1–42 is drastically limited into SPs in the subpial molecular layer and pyramidal cell layer in the cerebral parenchyma (Fig. 2). In particular, MALDI-MSI showed subpial Aβ1–42 deposition along the subarachnoid space despite a 100 μm pitch resolution. MSI spectral data of leptomeningeal blood vessels show different Aβ patterns for vascular features in the subarachnoid space, the arterioles, and cerebral parenchyma (Fig. 3).
MALDI-MSI of AD/CAA brain tissue at 20 μm resolution showing C-terminal and N-terminal truncated Aβ isoform profiles in amyloid deposits in AD with severe vascular pathology. Aβ1–36 to Aβ1–41 are found to be preferentially deposited in leptomeningeal blood vessels. Aβ1–42 and Aβ1–43 are deposited in the cerebral parenchyma within senile plaques. Dotted squares of a, b, and c are selected and are enlarged in the lower panel. Modified from Ikegawa et al. (2019).
In this study, Aβ1–40, along with Aβ1–42, was detected in vessels and in the cerebral parenchyma, with the Aβ1–40 in CAA to exceed the Aβ1–40 levels in parenchymal senile plaques, something that has been further validated by IHC, as outlined in Figure 2. Accumulation of Aβ1–38 and Aβ1–41 in the leptomeningeal blood vessels occurred in the cognitive normal brains at first and followed accumulation of Aβ1–36 to Aβ1–40 in the leptomeningeal blood vessels in AD (Kakuda et al., 2020; Kakuda et al., 2017).
In addition to full-length Aβ peptides starting at Asp-1, several N-terminally truncated Aβ species have been identified, including truncated peptides with Aβ2–x to Aβ11–x, including pyroglutamated species at Aβ3–x and Aβ11–x (Bayer and Wirths, 2014; Kummer and Heneka, 2014; Mukherjee et al., 2021; Portelius et al., 2010). In the previous studies of conventional antibody-mediated approach, N-truncated Aβ peptides were shown to enhance Aβ aggregation (Pike et al., 1995). Notably, the accurate profiles of N-truncated Aβ species have first been delineated by Aβ immunoprecipitation combined with MS (Table 1). Still it is challenging to understand the spatial and temporal dynamics of the formation of truncated Aβ species in precipitating amyloid pathology in vivo, which largely relates to the limitations of conventional molecular biology techniques. Consequently, it is still not fully understood what the functional and pathological relevance of these peptides is in AD pathogenesis.
Heterogeneity of Human Amyloid-Beta Isoforms Identified with Immunoprecipitation-Mass Spectrometry and Matrix-Assisted Laser Desorption Ionization-Mass Spectrometry Imaging
Both N- and C-terminal truncation and modification form of Aβ from brain, CSF and plasma were characterized by different research groups. The total number of Aβ proteoforms identified is specified including the corresponding references.
Aβ, amyloid-beta; CSF, cerebrospinal fluid; IMS, imaging mass spectrometry; IP, immunoprecipitation; MALDI, matrix-assisted laser desorption ionization; MS, mass spectrometry; PTM, posttranslational modification.
In this study, MALDI-MSI can play a pivotal role for studying amyloid formation dynamics as the technique offers the necessary molecular specificity, sensitivity, and resolution, superior to IHC. This in turn allows to clarify previously conflicting data on Aβ truncation in AD pathogenesis. For example, Masters and Selkoe (2012) and Masters et al. (1985) identified Aβ peptides beginning with phenylalanine at position 4 in both AD and DS cases. While Aβ4–x peptides are among the most abundant Aβ species, little is known about their spatial distribution in AD brains. Wirths et al. (2017) have generated antibodies selective for Aβ4–x to study these N-truncated Aβ species in both transgenic AD mice and in human AD brain tissue to conclude its significance.
In our MALDI-MSI data, distribution of Aβ4–42 is directly proved to be quite specific to the subpial area of occipital cortex, while full-length Aβ1–42 deposited mainly in the parenchyma as SP (Fig. 4D) (Kakuda et al., 2017).
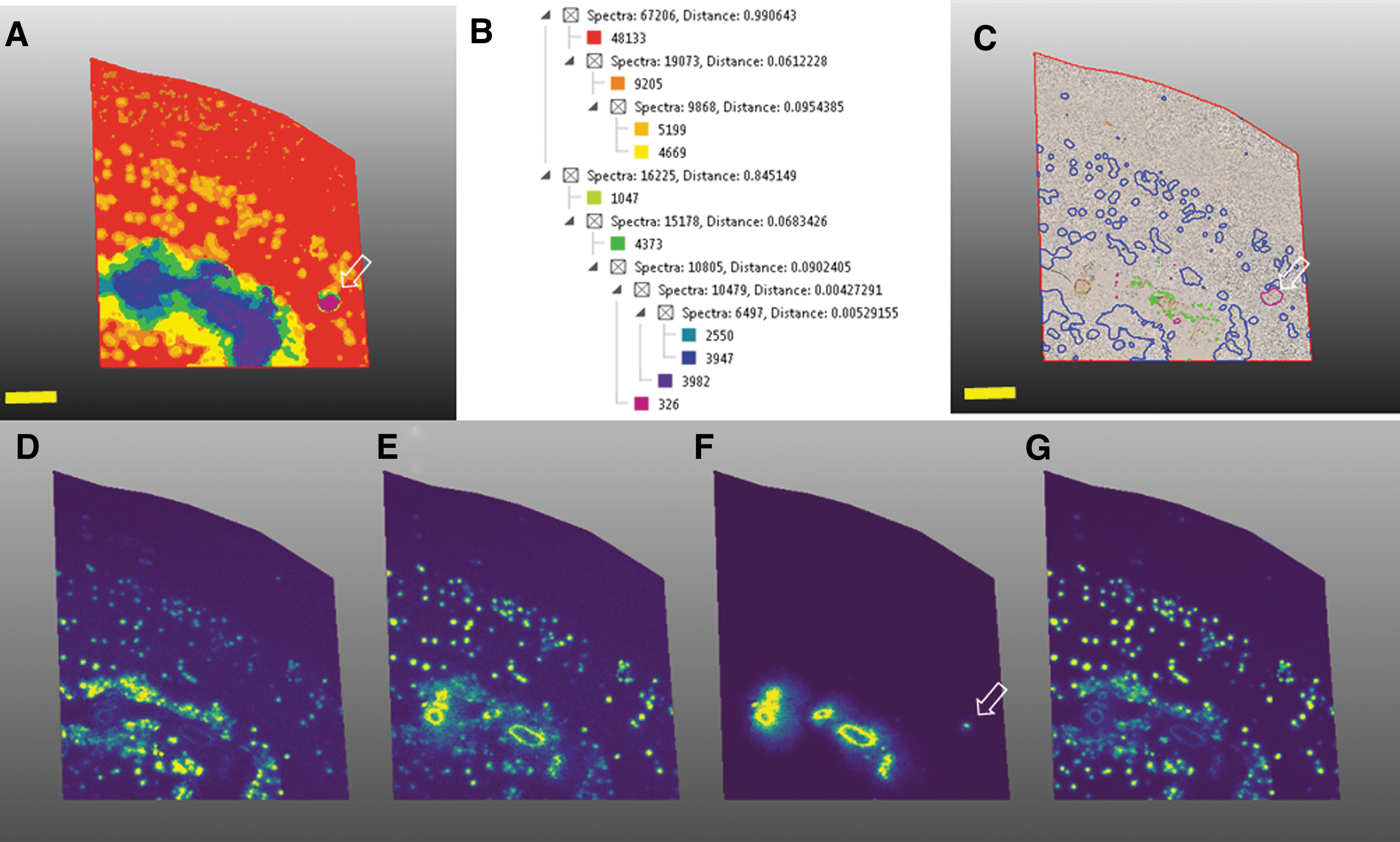
Multivariate image segmentation using HCA (bisecting k-means) reveals characteristic plaque pathology including senile plaques, large structures in the subarachnoid vasculature, and small arterioles in the parenchyma.
N3pE-Aβ42 peptides are the most abundant Aβ species in SP, more so than Aβ1–42. The antibody against the N-terminus of N3pE-Aβ is not able to distinguish between N3pE-Aβ40 and N3pE-Aβ42. In contrast, MALDI-MSI showed that N3pE-Aβ42 deposition shows an independent pattern than the one observed for Aβ1–42. Therefore, both N-terminally truncated Aβx–42 (with x = 2–11, including Aβ 3pE-42 and Aβ 11pE-42) and Aβ1–42 make aggregation core in SP. Possibly, more soluble N-truncated Aβ species are entrapped more distally in their drainage pathway (Fig. 3), while more fibrillinogenic Aβ isoforms are more likely to aggregate in the extracellular spaces in brain tissue and will therefore not reach extracellular fluids through the corresponding drainage pathways.
Recently, Michno et al. (2019a) published an application of MALDI-MSI into the study of intraplaque heterogeneity of a protein landscape. In this study, using a hyperspectral chemical imaging strategy, they reported that plaque maturation of previously diffuse deposits into cored, senile plaques correlated with increased deposition of Aβ1–40 in those plaques. MALDI-MSI showed that Aβ1–40 deposition occurred characteristically at the core of those cored/mature SPs. In contrast, Aβ1–42 showed localization to diffuse plaques. In addition, in sporadic AD, diffuse plaques showed elevated content of pyroglutamated Aβx–42, which, however, was not observed in diffuse plaques in cognitively unaffected amyloid-positive patients. This indicates that pyroglutamation is an AD pathology-specific event that is suggested to constitute a functionalization of Aβx–42 in otherwise nontoxic diffuse plaques, increasing Aβ hydrophobicity and in turn promoting deposition of less hydrophobic but more abundant species, mainly Aβ1–40 (Michno et al., 2019a).
Heterogeneity of Aβ proteoforms in vascular deposits
MALDI-MSI can visualize the whole distribution of the diverse Aβ proteoforms with multiple modifications, so far mainly N-terminal modifications. As for C-terminal Aβ truncations in leptomeningeal vessels and arteriole walls, MALDI-MSI delineated preferential localization of Aβ1–36 to Aβ1–41. In contrast, both Aβ1–42 and Aβ1–43 were formed to localize to cored plaques in the cerebral parenchyma of the occipital cortex (Table 1 and Figs. 2–4).
C-terminal truncation is a consequence of γ-secretase activity and results in formation of a variety of Aβ x–y species with y: 36–43 (Kang et al., 1987). MALDI-MSI was demonstrated to delineate deposition of all of those Aβ isoforms in the human brain (Kakuda et al., 2017). The heterogeneity in C-terminal truncation has been suggested to be a secondary event following initial γ-cleavage of APP between amino acid residues Leu49 and Val50 at the membrane/cytoplasmic interface (Weidemann et al., 2002). Ihara et al. proposed an Aβ processing model, where the β-C-terminal fragment is sequentially processed by γ-secretases (Funamoto et al., 2020; Kakuda et al., 2006; Matsumura et al., 2014; Morishima et al., 2000; Qi-Takahara et al., 2005). This model suggests that Aβ1–38 is a product of initially generated Aβ1–45 through Aβ1–42.
In contrast, Aβ1–41 is suggested to be generated from Aβ1–45 by sequential γ-secretase processing (Funamto et al., 2020; Matsumura et al., 2014; Qi-Takahara et al., 2005; Suzuki et al., 1994; Takami, 2009). The amounts of deposited Aβ1–38 correlated with those of Aβ1–40 and the ratios of deposited Aβ1–38/Aβ1–42 and Aβ1–40/Aβ1–43 were proportional and discriminated the Braak SP stages accurately. This indicates that the generation of Aβ1–38 and Aβ1–40 decreased and the generation of Aβ1–42 and Aβ1–43 increased with advancing SP stages (Kakuda et al., 2020).
Among those studies, Aβ1–41 has been suggested to be solely a minor component in human AD tissue. Notably, Aβ1–41 in human brains was shown for the first time through MALDI-MSI and further validated with IHC. It has been hypothesized that subtle C-terminal sequence modification by as little as one amino acid residue has significant effects on the peptide's aggregation propensity (Kakuda et al., 2017). This is due to observations where vascular deposition of Aβ1–41 was found predominantly in the arterial smooth muscle, whereas Aβ1–40 was found to localize to the adventitia region of blood vessels. In this study, Aβ41 aggregation appeared restricted within the intramural, periarterial drainage pathways. In contrast, Aβ40 showed translocation across the smooth muscle basement membranes to the pial–glial basement membranes, as a consequence of less aggregation and higher solubility, respectively.
The common role of Aβ deposition in CAA and AD is recently reviewed as a cross talk between cerebrovascular and neurodegenerative pathologies (Greenberg et al., 2020). The intersections between these two diseases indicate a possibility of improving vascular function in the development of novel therapies. Both AD and CAA pathologies share common features with respect to secretion levels, circulation, and clearance of Aβ. In our Aβ proteoform mapping of human brains with MALDI-MSI, avenues for developing novel therapy as well as diagnostic indices are to be accelerated.
The process of excretion of Aβs through the perivascular drainage pathway from the cerebral parenchyma to the lymph node has been extensively studied by Weller et al. (2009a), Weller et al. (2009b), Weller et al. (2008), Weller et al. (1998), Hawkes et al. (2011), and Morris et al. (2016). Another advantage of adopting MALDI-MSI for in situ proteomics of the human brains is to generate a segmentation map at 20 μm resolution, which in turns supports the theory for brain-specific, perivascular drainage pathways, significantly contributing to both pathogenesis of CAA and AD (Fig. 4) (Ikegawa et al., 2019).
Furthermore, these results allow identification of other proteins that colocalized with SP and subarachnoid vasculature (Fig. 4). Those species might in turn serve as potential biomarkers for heterogenous amyloid pathology. It is of note that a single mass distribution from Aβ4–42, 3pE-42, and 1–42 clearly represents a dominant localizing area: subpial area in the cortex, vascular and parenchymal area, and parenchymal area, respectively. By contrast, Aβ1–40 predominantly distributed in the leptomeningeal vasculature as well as parenchymal small artery, as shown by the arrow in Figure 4A, C, and F.
Following Amyloid Aggregation Over Time
A significant limitation in studying amyloid formation dynamics is the lack of temporal information as all the imaging modalities used for probing amyloid plaque pathology are static. MS techniques have gained great impact in biomolecular and clinical research to probe protein dynamics in complex biological matrices comprehensively (Aebersold and Mann, 2003). Indeed, MS-based proteomics in combination with metabolic labeling with stable isotopes allowed to quantify protein turnover dynamics, respectively, in tissue extracts and body fluids (McClatchy et al., 2007; Ong et al., 2002; Westman-Brinkmalm et al., 2011). For these time course proteomic experiments, stable isotopes (e.g., 15N, 13C, 18O, or 2D) are provided systemically by infusion or diet and incorporated into newly synthesized proteins (i.e., metabolic labeling). This approach has been successfully developed for following protein dynamics in vivo in AD patients.
In this study, patients received intravenous infusions of 13C6-Leu, which was followed by repetitive cerebrospinal fluid (CSF) drainage over a continuous spinal tab. This pioneering work allowed to quantify Aβ and tau turnover and clearance dynamics and revealed novel insights into mechanisms associated with plaque pathology such as impaired clearance of pathogenic proteoforms (Bateman et al., 2006; Paterson et al., 2019). Furthermore, these methods enabled us to follow amyloid pathology-associated protein regulations in AD mouse models (Hark and Savas, 2021; Hark et al., 2021; Savas et al., 2017). A limitation, as with all ex situ studies, is the lack of spatial information on local isotope incorporation and protein turnover, respectively.
Combining stable isotope labeling and nanoSIMS-based imaging MS has been shown previously to probe the turnover kinetics of proteins and nucleotides in both cells and tissues (Steinhauser et al., 2012). Notably, the Bateman group describe a method to determine the production and clearance rates of proteins within the human CNS with stable isotope labelling (Bateman et al., 2006) and have demonstrated the use of nanoSIMS and stable isotope labeling to study amyloid deposition in postmortem brain from AD patients (Wildburger et al., 2018).
A significant limitation of nanoSIMS is the restricted molecular information. While the technique provides outstanding spatial resolution (50 nm) and sensitivity to reveal spatial isotope enrichment, no data can be obtained for the actual labeled peptides and proteins. This limitation can be addressed through MALDI-MSI as the technique allows in situ visualization of intact amyloid peptides as outlined above. Our group has recently made use of these developments by combining metabolic labeling with multimodal MSI for imaging stable isotope labeling kinetics (iSILK) to follow amyloid aggregation in evolving plaque pathology in novel, APP knockin mice (APPNL-G-F) (Michno et al., 2021).
To establish the early events of Aβ aggregation and heterogenous plaque formation, we performed a series of elaborate complementary PULSE/CHASE setups where we supplied 15N enriched protein diet from before and during plaque onset (PNW 10) followed by a washout (chase) period of up to 10 weeks. Comprehensive MALDI and nanoSIMS-based MS imaging demonstrate that the method of feeding the stable isotopes works and leads to detectable levels of label within the plaques for meaningful analysis. The results show that plaques in APPNL-G-F mice form via early deposition of Aβ1–42 into compact core (5 μm in size), which is followed by plaque growth by homogenous deposition throughout the plaque. Moreover, it was clearly evidenced that plaques form first in the cortical areas before forming deposits in the hippocampus and other subcortical areas.
Finally, the iSILK study has shown that in APPNL-G-F mice, primary events in amyloid pathology involve secretion and deposition of Aβ1–42 followed by a later secretion deposition of Aβ1–38 (Michno et al., 2021).
MALDI-MSI of Plaque Pathology-Associated Lipid Species
Lipid analysis is of great relevance in neuroscience research as it has significant roles as both signaling molecules and structural interactions with membrane proteins. Especially, it has been well established that lipids interact with self-aggregating proteins, such as amyloidogenic proteins, and even promote aggregation of these proteins in a pathological context (Jucker and Walker, 2013). Aggregation of amyloidogenic proteins into insoluble deposits is a seminal histopathological characteristic in many neurodegenerative diseases including AD, frontotemporal lobe dementia, Huntington's disease, PD, amyotrophic lateral sclerosis, and Niemann–Pick disease (Blennow et al., 2006; Jucker and Walker, 2013). Generally speaking, lipids with the lack of sufficient molecular fingerprints were due to limited “information” compared with proteins made from 20 amino acids; it is challenging to obtain pathological implications directly from tissue lipid distributions. However, MALDI imaging offers here the unique opportunity to characterize the protein pathology-associated lipid biochemistry, which in turn provides targets for mechanistic investigations.
In this context, MALDI imaging has most prominently been used for delineating amyloid plaque pathology-associated lipid dysregulations in AD. Specifically, while MALDI imaging identified plaque-specific localizations of gangliosides, ceramide, lysophosphatidic acids, and inositol, sulfatides in turn were depleted at plaques (Michno et al., 2019c). Moreover, when interfaced with advanced fluorescent microscopy using structure-sensitive luminescent probes, MALDI-MSI identified lipid patterns that could be correlated with Aβ plaque polymorphism (Michno et al., 2019b; Michno et al., 2018). Of note, ganglioside GM1 was found to correlate with the core structure of senile plaques, while ceramides and phosphoinositols (PI) located to the diffuse periphery (Michno et al., 2019b; Michno et al., 2019c) (Fig. 5).
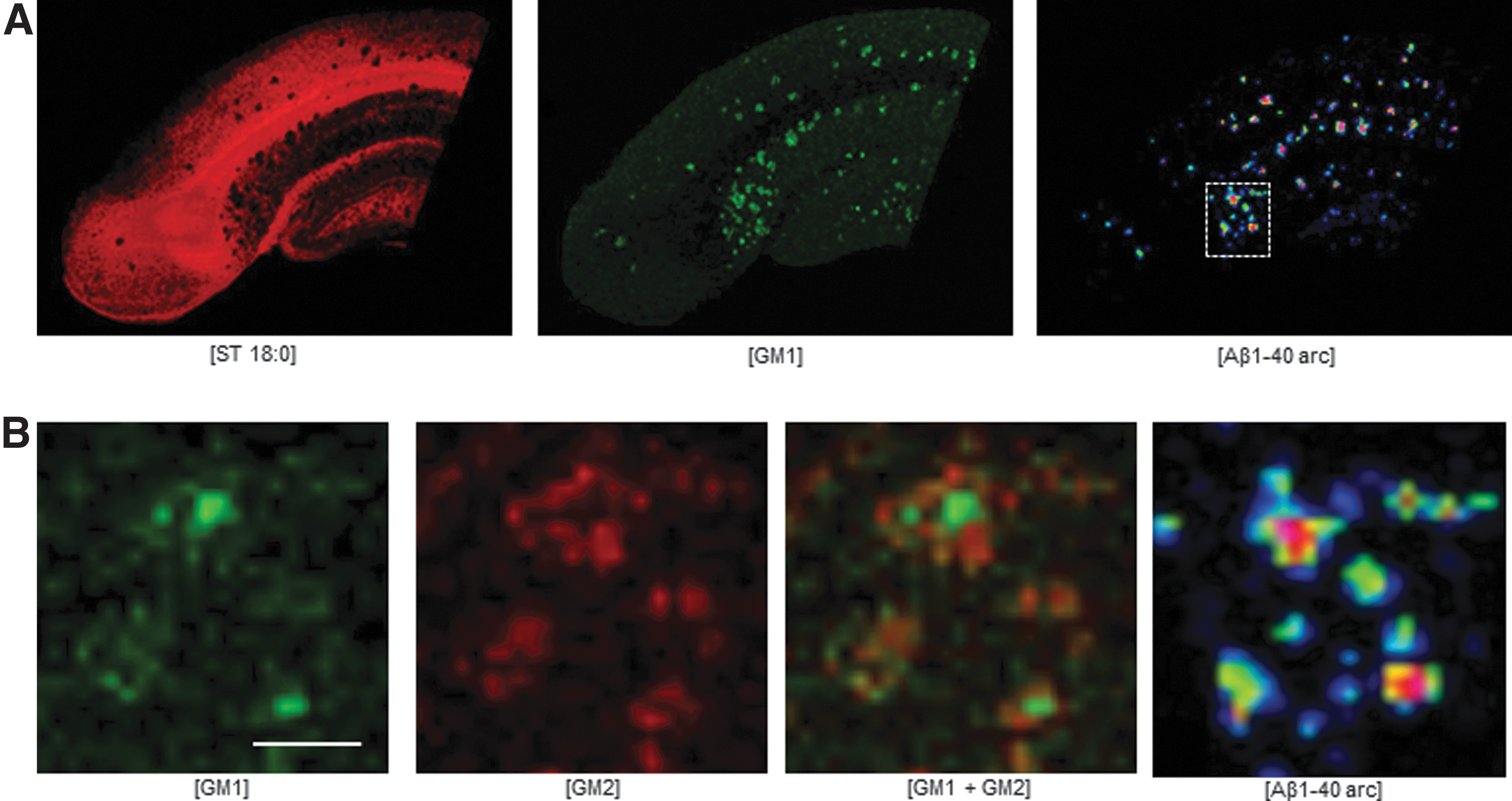
Correlative, multimodal lipid and peptide imaging within the same section identified plaque pathology-associated changes in ganglioside metabolism.
A further development toward multimodal imaging approaches includes bimodal (Michno et al., 2019b; Michno et al., 2018) and trimodal (Kaya et al., 2018; Kaya et al., 2017b) correlative lipid and peptide imaging within the same tissue. This approach is facilitated by the low laser desorption energies, fluences, and shot numbers necessary to obtain satisfactory lipid spectra using 1,5-DAN matrix dry coating protocols or sublimation (Kaya et al., 2017b; Kaya et al., 2017c).
With the current strategy, in situ lipid changes were shown to be associated with protein mechanisms as identified with imaging mass spectrometry, spectroscopy, and conventional microscopy techniques (Michno et al., 2019a; Michno et al., 2018). Of note, distinct lipids such as ganglioside GM1 were found to correlate with the 40 amino acid species of Aβ (Aβ1–40) within spectroscopy assigned core structures. In contrast, other lipids, including ceramides and PI species were observed in a negative mode and lysophosphocholin was detected in a positive mode, correlate with diffuse parts and Aβ1–42, respectively.
Future Perspectives
A variety of modern proteomic studies have proposed several excellent strategies for deciphering protein identification as well as protein–protein interactions. This includes extracellular matrix proteins as well as inflammation markers, acute-phase proteins, cytokines, immunoglobulins, complement cascade proteins, and components of the inflammasome were found in the vicinity of amyloid pathology (Masters and Selkoe, 2012), which can play as drivers of amyloid accumulation. This way, copurification of senile plaques has revealed proteins of yet not understood pathogenic relevance, such as a fragment (60–95 aa) of α-synuclein protein. Intraneuronal accumulation of α-synuclein is indeed observed in some cortical nerve cells, which explains how these species are detected in copurified cortical tissue homogenates.
Moreover, most of the individual neuropathology of AD and/or CAA must have overlapping features with so-called proteinopathies, one of the standard future applications of MALDI-MSI for AD and CAA brains must be an integration of in-depth proteomics obtained by MALDI-MSI with tissue digestion and shotgun analysis (Toyama et al., unpublished data).
In general, MALDI-MSI of proteins potentially faces a number of obstacles such as a limit of mass range of analysis. If formalin-fixed, paraffin-embedded samples are to be analyzed, enzymatic digestion must be an essential step in protein identification (Spengler, 2014). A direct approach to protein analysis involves detecting and identifying them on tissues in MS imaging analyses, but this requires that the protein digestion is performed on the tissue section with restricted lateral diffusion to maintain spatial resolution.
Recently, with current strategy, Park et al. (2022) have identified neuronal intranuclear inclusions (NIIs) in brains of neuronal intranuclear inclusion disease (NIID) by combining LC–MS/MS shotgun proteomics and MALDI-MSI. They hypothesized that the NIIs of diseased human brains may have proteins with compositionally biased regions, and really identified a serine-rich protein called hornerin. Since the analyzed NII-rich fraction was also the serine-rich, hornerin, a major component of the NIIs was suggested and a specific distribution of hornerin in NIID not in control or AD brains was also investigated by the on tissue digestion protocol of MALDI-MSI.
Another aspect of future application of MALDI-MSI can be a generating 3D-MSI model of AD pathology. Very recently, Enzlein et al. (2020) have succeeded in obtaining 3D images of Aβs with MALDI-MSI on APPNL-G-F mouse at 67 weeks of age. They proposed a seed mechanism, where longer and less abundant peptides may seed the aggregation of more abundant ones. Implementing MALDI-MSI for standardized 3D lipidomics and proteomics is a promising approach to understand the intracellular interplay across histopathological features to grasp a more comprehensive view on the pathogenesis of various neurodegenerative diseases (Enzlein et al., 2020). Furthermore, the characterization of Aβ proteoforms in human brains could lead to the targeted analysis of CSF or plasma (Masters et al., 2018; Nakamura, 2018) in nondemented subjects to identify which proteoforms may be predictive markers for progression to clinical dementia by connecting a fluid biomarker study with an MALDI-MSI study.
Footnotes
Acknowledgments
We are deeply indebted to Prof. Yasuo Ihara (Doshisha University) and Prof. Shigeo Murayama (The Brain Bank for Aging Research, Tokyo Metropolitan Geriatric Hospital and Institute of Gerontology) for the gift of current opportunity for launching the MALDI-MSI study of AD and CAA brains. We also acknowledge Prof. Colin L. Masters (The Florey Institute, The University of Melbourne) and Prof. Nobuyuki Nukina (Doshisha University) for their great concerns to MALDI-MSI and encouragements. Profs. Shoichi Ishiura and Satoru Funamoto (Doshisha University) should be acknowledged for many useful scientific discussions.
Authors' Contributions
M.I. and J.H. drafted the structure of the review. M.I., N.K, T.M., Y.T., T.N., K.M., and J.H. edited the figures and wrote the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported, in part, by the Grant-in-Aid for Scientific Research on Innovative Areas (Brain Protein Aging and Dementia Control 26117004, MI). This research was partially supported by the Strategic Research Program for Brain Sciences from Japan Agency for Medical Research and Development, AMED (MI). Jörg Hanrieder acknowledges The Swedish Research Council (No. 2018-02181), Alzheimer Research U.K., Alzheimerfonden, Demensfonden, Ahlén Stiftelsen (Jörg Hanrieder), Stiftelsen Gamla Tjänarinnor, Gunoch Bertil Stohnes Stiftelse, and Magnus Bergvalls Stiftelse for financial support.