
Abstract
Objectives:
Attention-deficit hyperactivity disorder (ADHD) in adulthood shows high co-occurrence rates with cocaine use disorder (CoUD). The self-medication hypothesis (SMH) provides a theoretical explanation for this comorbidity. This study investigates the neurobiological mechanisms that could support SMH in adult patients with attention-deficit hyperactivity disorder with cocaine use disorder (ADHD–CoUD).
Materials and Methods:
We included 19 ADHD–CoUD patients (84.2% male; age: 32.11 years [7.18]) and 16 CoUD patients (68.7% male; age: 36.63 years [8.12]). All subjects underwent a fluorine-18-fluorodeoxyglucose positron emission tomography (18F-FDG PET) brain scan. We tested brain metabolism differences between ADHD–CoUD and CoUD patients using voxel-based and regions of interest (ROIs)-based analyses. The correlation between dependence/abstinence duration and regional brain metabolism was also assessed in the two groups. Lastly, we investigated the integrity of brain metabolic connectivity of mesocorticolimbic and nigrostriatal dopaminergic systems, and large-scale brain networks involved in ADHD and addictions.
Results:
The voxel-wise and ROIs-based approaches showed that ADHD–CoUD patients had a lower metabolism in the thalamus and increased metabolism in the amygdala and parahippocampus, bilaterally, than CoUD subjects and healthy controls (HCs). Metabolism in the thalamus negatively correlated with years of dependence in ADHD–CoUD patients. Moreover, connectivity analyses revealed that ADHD–CoUD patients had a more preserved metabolic connectivity than CoUD patients in the dopaminergic networks and large-scale networks involved in self-regulation mechanisms of attention and behaviors (i.e., anterior default mode network [ADMN], executive network [ECN], and anterior salience network [aSAN]).
Conclusions:
We demonstrated distinct neuropathological substrates underlying substance-use behaviors in ADHD–CoUD and CoUD patients. Furthermore, we provided neurobiological evidence in support of SMH, demonstrating that ADHD–CoUD patients might experience short-term advantages of cocaine assumption (i.e., compensation of dopaminergic deficiency and related cognitive-behavioral deficits).
Impact statement
This study provides neurobiological evidence for self-medication hypothesis (SMH) in adults with attention-deficit hyperactivity disorder with cocaine use disorder (ADHD–CoUD). The current results suggest differential treatment approaches, namely pharmacological approaches for cocaine use disorder (CoUD) individuals with and without attention-deficit hyperactivity disorder (ADHD), to improve functional adjustment and reduce the risk of relapse in addictive behaviors. Specifically, stimulant pharmacological treatments (e.g., methylphenidate- and amphetamine-based stimulants) with a long-acting formulation should be considered the first line of intervention for adults with ADHD–CoUD. Furthermore, these pharmacological treatments could be combined with evidence-based behavioral interventions for emotional dysregulation in patients with substance use disorders and ADHD.
Introduction
Attention-deficit hyperactivity disorder (ADHD) is one of the most common neurodevelopmental disorders, affecting ∼5% of children and early adolescents worldwide (Polanczyk et al., 2014). ADHD persists throughout adulthood in up to 57% of patients (Fayyad et al., 2017). About 51.7% of adult patients with ADHD had at least one psychiatric comorbid disorder; substance use disorders (SUDs) are the most co-occurring conditions (Simon et al., 2009). Consistently, 20% of adult treatment-seeking individuals with SUDs meet the criteria for ADHD (van de Glind et al., 2014). Furthermore, a considerable percentage of treatment-seeking adult individuals with ADHD (∼10%) develop cocaine use disorder (CoUD) (Oliva et al., 2021).
One of the most interesting explanations of the robust association between ADHD and CoUD is the self-medication hypothesis (SMH). The SMH gives a valuable perspective to understanding the emotional and psychological dimensions of reliance on drugs (Khantzian, 1997), suggesting that personality organization, individual characteristics, and inner psychological states can drive specific drugs assumption as the best practical solution to relieve such suffering and distress. Individuals with ADHD bump into cocaine abuse because of its modulatory effects on hyperactivity, inattention, and behavioral dysregulation (Mariani et al., 2014).
On the contrary, individuals who use cocaine without a history of ADHD choose this substance to relieve feelings of boredom, emptiness, and fatigue, or to elate sensations and energy (Suh et al., 2008). However, no empirical studies have demonstrated this hypothesis, and its neurobiological foundations need more clarification. Further research may reveal mechanisms involved in developing and maintaining different forms of addiction.
Cocaine is a psychostimulant; it binds to the dopamine transporter (DAT) and blocks its physiological function (dopamine reuptake), increasing dopamine availability in the synaptic cleft. It has been demonstrated that cocaine abuse is responsible for a dopamine signaling adaptation in mesocorticolimbic and nigrostriatal dopaminergic pathways (Volkow et al., 2001). Disturbances in these pathways can cause impaired cognitive abilities, motivational deficits, and alteration of voluntary movement control in ADHD (Gold et al., 2014).
Decreased attention, restlessness, and impaired learning in patients with ADHD seem to be caused by the pathological increased synaptic reuptake of dopamine, resulting in reduced levels of extracellular synaptic dopamine (Levy, 1991). The crucial role of dopaminergic systems in the pathophysiology of ADHD and the impact of the modulation of cocaine effects on brain functioning might represent the underlying neurobiological substrate in supporting SMH principles in these patients.
Fluorine-18-fluorodeoxyglucose positron emission tomography (18F-FDG PET) is a functional imaging technique that helps understand the brain alterations underlying psychopathological states. 18F-FDG PET represents a proxy for neuronal activity and an index of synaptic function and density, and it is used to measure resting-state cerebral metabolic rates of glucose (Perani et al., 2020). The application of multivariate methods to 18F-FDG PET data can reliably contribute to the in vivo access of the biochemical and functional architecture of the brain, such as brain metabolism, metabolic network connectivity of large-scale networks, and neurotransmission systems (Sala et al., 2023).
This study investigates possible neurobiological mechanisms supporting the SMH in adult CoUD subjects with (i.e., attention-deficit hyperactivity disorder with cocaine use disorder [ADHD–CoUD]) and without a history of ADHD (i.e., CoUD). To do so, we investigated the differences between ADHD–CoUD and CoUD patients in (1) short- and long-term effects of cocaine on brain functioning as measured with 18F-FDG PET data, (2) metabolic connectivity alterations of mesocorticolimbic and nigrostriatal dopaminergic systems, and (3) the integrity of major large-scale brain networks involved in ADHD and cocaine addiction.
We hypothesized the presence of brain compensation mechanisms related to cocaine assumption in ADHD–CoUD patients; specifically, we expected these mechanisms to involve the dopaminergic and large-scale brain networks underlying core psychopathological domains of ADHD (i.e., self-regulation of attention and behavior).
Materials and Methods
Participants
Expert psychiatrists enrolled the patients at the Psychiatric Disorders Centre of Ospedale di Prato (NOP) S. Stefano, Prato, Italy, between 2016 and 2018. Empirical evidence supported the reliability of unstructured clinical interviews based on DSM diagnostic criteria for several mental disorders, including substance-related disorders (Miller et al., 2001). Expert psychiatrists conducted a traditional diagnostic assessment (Sharp et al., 2013) based on DSM-5 (American Psychiatric Association, 2013) criteria to assess CoUD. Thirty-five patients were clinically diagnosed as CoUD and included in the study.
Structured evaluations of adult ADHD compared with unstructured clinical assessments are recommended (Schneider et al., 2019). At the time of recruitment, DIVA 2.0 was the available semistructured diagnostic interview for ADHD in adults based on DSM-IV-TR criteria (American Psychiatric Association, 2000). All 35 patients underwent DIVA 2.0 (Kooij and Francken, 2010), administered by a trained psychiatrist to assess the presence of adult ADHD.
Nineteen patients fulfilled the diagnosis of adult ADHD. Eight patients with ADHD (42%) suffered from the inattentive subtype, four participants (21%) met the criteria for the hyperactive-impulsive subtype, and seven patients (37%) showed the ADHD combined type. Abstinence maintenance was self-reported by patients and demonstrated through a urine toxicological screening. The participants did not present other neurological conditions, including pervasive developmental disorders and epilepsy. All patients (ADHD–CoUD and CoUD) also underwent an 18F-FDG PET brain scan.
Thirty gender-matched healthy controls (HCs) were included from the internal database of the In Vivo Human Molecular and Structural Neuroimaging Unit, IRCCS San Raffaele Scientific Institute, Milan, Italy. They did not have a history of neurological or psychiatric diseases or other chronic illnesses and were not taking psychoactive medication.
The local ethical committees approved this research study according to the Helsinki Declaration. All participants or their caregivers approved and signed a informed consent form.
Regional brain metabolism analysis
Section S1 in the Supplementary Data contains details of images acquisition and preprocessing. We tested brain metabolism differences between ADHD–CoUD and CoUD patients using both voxel-based and regions of interest (ROIs)-based methods. We performed a voxel-wise group comparison through a Statistical Parametric Mapping (SPM) two-sample t-test, entering age as a variable of no interest. The p-value was set at p uncorrected <0.005 with cluster extent (Ke)p ≥ 100 voxels. We also analyed the differences in the means uptake of metabolism among the three groups (ADHD–CoUD, CoUD, and HCs) in ROIs with crucial relevance for cognitive and behavioral functioning in ADHD and CoUD conditions (Ivanov et al., 2010; Koob and Volkow, 2010; Mehta et al., 2019).
Based on literature, we selected the following ROIs: frontopolar cortex (BA10), anterior cingulate, medial frontal orbital cortex, hippocampus, amygdala, dorsal caudate nucleus, dorsal putamen, ventral striatum, globus pallidus, thalamus, cerebellum hemispheres, and vermis. We extracted the mean values of metabolism from each ROI for all subjects. ROIs were mainly derived from the Automated Anatomical Labelling (AAL) atlas (Tzourio-Mazoyer et al., 2002). However, due to some AAL ROIs' relative coarseness, we complemented the selected AAL ROIs with more refined ROIs, especially cerebellar regions and frontopolar cortex (BA10), derived from dedicated atlases (Harvard Oxford subcortical Atlases available in FSL and ANATOMY toolbox atlas).
Given the thalamus's functional-anatomical complexity in terms of connections with the brain cortex, we used ROIs derived from the Oxford thalamic connectivity atlas that segmented the thalamic gray matter based on white-matter connectivity traits to six cortical areas (Behrens et al., 2003). Specifically, this atlas identifies six gray-matter regions named in accordance with their projection to the cortex: thalamus–motor, thalamus–parietal, thalamus–prefrontal, thalamus–premotor, thalamus–temporal, and thalamus–visual.
We computed a linear regression analysis with each ROI's metabolism as a dependent variable and age as an independent variable. The unstandardized residuals were retained to represent age-adjusted metabolism values, excluding the aging effect in the group comparisons. Then, we performed the analysis of variance with Bonferroni correction for normally distributed variables and Kruskal–Wallis with Bonferroni correction for nonparametric variables to study the metabolic difference among CoUD with and without ADHD and HC groups in the selected ROIs. The statistical threshold used was p < 0.05.
Lastly, we performed bivariate spearman correlation analyses to test the correlation between dependence/abstinence duration and age-adjusted metabolism values of each ROI. The statistical threshold was p < 0.05 (Benjamin–Hochberg corrected for multiple comparisons). Statistical analyses were performed using SPSS 26.0 (SPSS Inc., Released 2019; SPSS for Windows, Version 26.0; SPSS, Inc., Chicago, IL) and RStudio for Windows.
Brain metabolic connectivity analyses
Neurotransmitters networks analyses
According to well-validated node selection and ROIs definition procedures (Caminiti et al., 2017; Carli et al., 2020), we have addressed neural networks related to dopaminergic neurotransmission systems—nigro-striato-cortical and mesolimbic pathways. Specifically, the two dopaminergic systems were assembled by considering the topographical organization of ascending dopaminergic projections based on biochemical, histochemical, and anatomopathological findings (Ciliax et al., 1999; Fallon, 1988).
ROIs used for the connectivity analyses were derived from the AAL atlas (
Partial correlation analysis
We created a subject-by-Node/ROI matrix for each group (i.e., ADHD–CoUD, CoUD, and HCs) to assess the connectivity within the nigro-striato-cortical and the mesolimbic dopaminergic pathways. The matrices contained the regional metabolic mean value extracted from each neurotransmitter network's specific ROIs for each subject.
We computed partial correlations using MATLAB for Windows (Mathworks, Inc., Sherborn, MA). Partial correlation is a statistical approach that allows measuring metabolic connectivity between two nodes while factoring out other regions' contributions (Sala and Perani, 2019). This multivariate method relies on good reproducibility and general applicability within experimental settings typical of PET neuroimaging studies (Veronese et al., 2019). The resulting partial correlation matrices were set at a statistical threshold of p < 0.01, uncorrected for multiple comparisons.
Age was entered as a nuisance covariate in the analyses. We applied Fisher's transformation to each coefficient to test whether the strength of the partial correlation coefficients (between nodes–dopaminergic networks) differed between groups (ADHD–CoUD and HCs; CoUD and HCs). Then, Z-test was used to test significant changes in partial correlation coefficients. We set the statistical threshold at p < 0.01, corrected for Bonferroni multiple comparisons. The resulting z-score matrices were then used to calculate the percentage of altered metabolic connections of each network and compare them between groups (ADHD–CoUD and CoUD). See Section S2 in the Supplementary Data for details of connectivity measures.
Metabolic connectivity in large-scale brain networks: interregional correlation analysis
We performed a brain metabolic connectivity analysis to investigate metabolic changes in large-scale brain networks of ADHD–CoUD and CoUD patients compared with HCs. We considered the large-scale brain networks whose seeds showed maximal involvement in ADHD and cocaine addiction, namely anterior and posterior default mode networks (ADMN and PDMN), executive network (ECN), attentive network (ATTN), sensory-motor network (SMN), basal ganglia network (BGN), limbic network (LIN), and anterior salience network (aSAN).
Based on the core principle that brain regions whose metabolism is correlated at rest are functionally interconnected (Horwitz et al., 1984), we applied a voxel-wise interregional correlation analysis (IRCA). This method was previously validated for 18F-FDG PET data and allowed to derive large-scale brain metabolic network connectivity starting from proper seed regions (Lee et al., 2008). Seed ROI was defined from the functional atlas of large-scale brain networks (Shirer et al., 2012) (Supplementary Table S3 in the Supplementary Data).
The mean 18F-FDG seed uptake, extracted separately for each clinical group, was set as a variable of interest in multiple regression models, testing for voxel-level correlations with the whole brain metabolic activity in different groups (ADHD–CoUD, CoUD, and HCs). Age was used as a nuisance variable. The statistical threshold was set at p-uncorrected <0.005, family-wise error-corrected at the cluster level, with Kep ≥100 voxels. Differences in network topography and spatial extension were measured according to the proper similarity index (i.e., Dice similarity coefficient) and the number of correlated voxels (Ncv) (Savio et al., 2017), respectively.
Results
Demographic and clinical features
The ADHD–CoUD and CoUD patients did not differ in demographic and clinical variables (Table 1). They presented a comparable percentage of males and females, age, years of CoUD, and days of abstinence before the scan.
Demographic and Clinical Features of Patients
HCs differed from both CoUD and ADHD–CoUD patients.
ADHD, attention-deficit hyperactivity disorder; ADHD–CoUD, attention-deficit hyperactivity disorder with cocaine use disorder; CoUD, cocaine use disorder; F, female; HCs, healthy controls; M, male.
Comparisons between groups in regional metabolism
Voxel-based analyses
ADHD–CoUD patients had reduced metabolism in the bilateral thalamus and increased metabolism in the bilateral amygdala and parahippocampus, compared with CoUD subjects and HCs (Fig. 1A). CoUD patients showed a lower and more diffuse metabolism than HCs in frontal regions: superior orbitofrontal gyrus, superior and middle frontal gyrus, bilaterally (Supplementary Fig. S1 in the Supplementary Data). ADHD–CoUD also showed lower metabolism than HCs in the frontal cortex but was limited to the right middle frontal gyrus (Supplementary Fig. S1 in the Supplementary Data).

Brain metabolism regional differences: voxel-wise and ROIs comparisons.
ROIs-based analyses
ADHD–CoUD patients had a lower metabolism in the thalamus than CoUD subjects and HCs. ADHD–CoUD patients also showed a higher metabolism than HCs in the amygdala. Both groups (ADHD–CoUD and CoUD) showed a lower metabolism than HCs in the frontopolar cortex; only CoUD patients had lower metabolism than HCs in the medial orbitofrontal cortex (Fig. 1B and Supplementary Table S4 in the Supplementary Data).
Correlation results
ADHD–CoUD patients showed a significant negative correlation between thalamus metabolism and years of addiction. ADHD–CoUD patients also showed a negative correlation between metabolism in the vermis and days of abstinence. In other words, ADHD–CoUD participants who reported fewer days of abstinence had a higher metabolism in the vermis (ρ = −0.58; p = 0.038). No significant correlation was found in CoUD patients (Fig. 2).
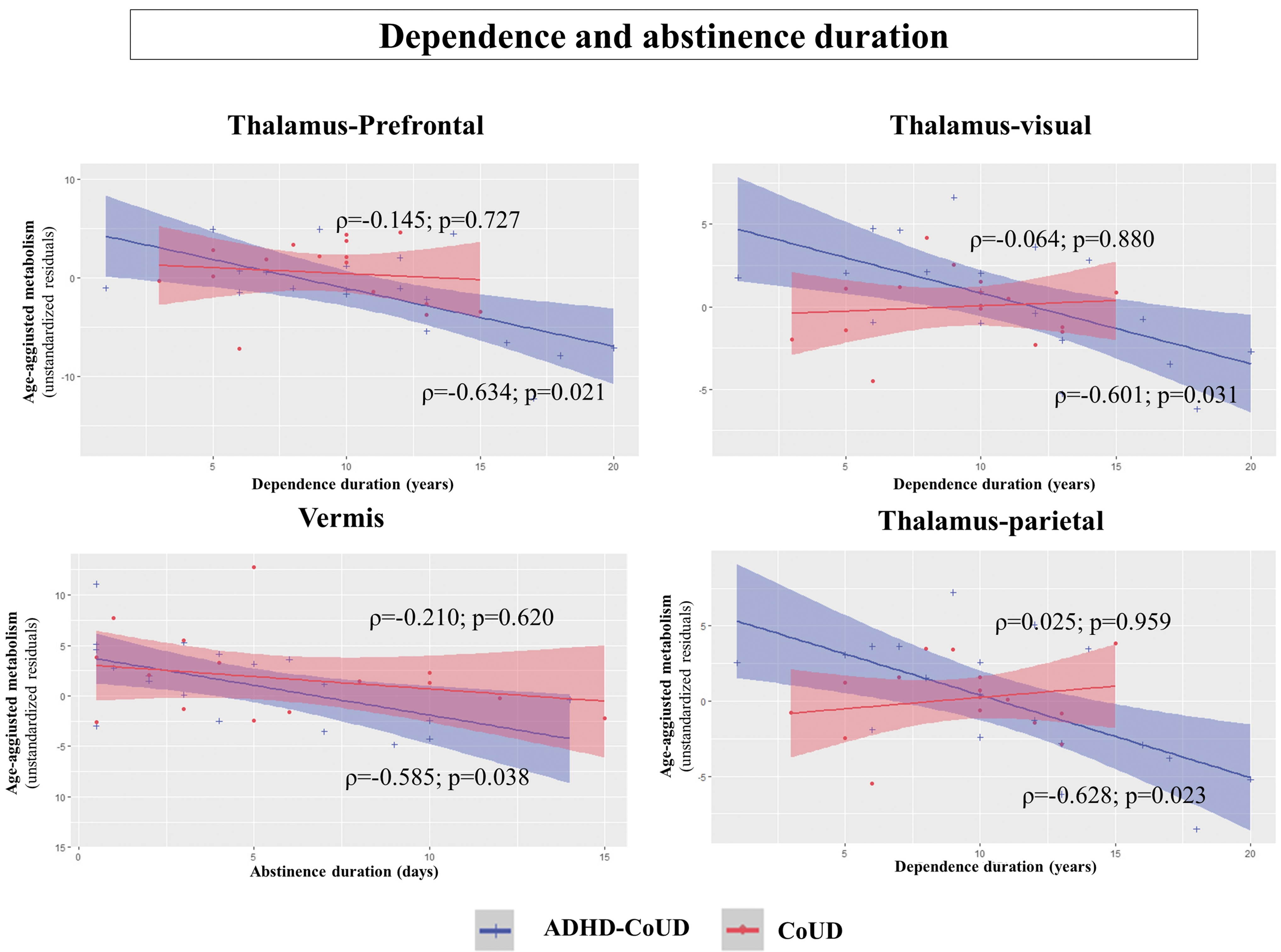
Correlation between dependence/abstinence duration and regional metabolism of each ROI. Significant correlations among age-corrected metabolism values and dependence duration (left) and abstinence duration (right) in ADHD–CoUD (blue) and CoUD (red) clinical groups (Benjamini-Hochberg correction for multiple comparisons). Among the 17 ROIs analyzed, only 4 showed a significant correlation with these 2 clinical variables: dependence and abstinence duration. The subregions of the thalamus were derived from the Oxford thalamic connectivity atlas that segmented the thalamic gray matter based on white-matter connectivity traits to six cortical areas: prefrontal, premotor, parietal, motor, temporal, and visual cortices (Behrens et al., 2003). The six gray-matter regions of the thalamus derived their name according to their projection to the specific cortices: thalamus–motor, thalamus–parietal, thalamus–prefrontal, thalamus–premotor, thalamus–temporal, and thalamus–visual. Three out of six thalamic ROIs significantly correlated with years of dependency duration: thalamus–prefrontal, thalamus–visual, and thalamus–parietal.
Metabolic connectivity
Nigro-striato-cortical network
CoUD patients demonstrated widespread alteration of metabolic connectivity within the nigro-striato-cortical dopaminergic network, showing 39% altered connections compared with HCs. On the contrary, the connectivity reconfiguration of the same network was substantially unaffected in the ADHD–CoUD group—3% of changes compared with HCs. The direct comparison between the clinical groups showed that metabolic connectivity alterations of the nigro-striato-cortical network were more severe in the CoUD than in the ADHD–CoUD subjects (χ2 = 35.5385; p < 0.00001). See Figure 3A for nigro-striato-cortical connectivity results.

Nigro-striato-cortical and mesolimbic cortical dopaminergic networks. ADHD–CoUD and CoUD dopaminergic networks are graphically represented with connectivity matrices and 3D brain templates. The matrices represent the significant differences obtained when comparing partial correlation coefficients between ADHD–CoUD patients (blue) and CoUD patients (red) with HCs in the dopaminergic nigro-striato-cortical network
Mesocorticolimbic network
The analyses found widespread connectivity alterations within the mesolimbic dopaminergic network among CoUD patients compared with HCs—53% of altered connections. On the contrary, the connectivity reconfiguration of the same network was limited in the ADHD–CoUD group compared with HCs—9% of altered connections. The direct comparison between clinical groups showed that metabolic connectivity alterations of the mesolimbic network were more severe in the CoUD subjects than in the ADHD–CoUD subjects (χ2 = 41.2698; p < 0.00001). See Figure 3B for mesolimbic connectivity results.
Metabolic connectivity in large-scale brain networks: IRCA results
ADMN and ECN
ADMN and ECN showed excellent overlap between ADHD–CoUD patients and HCs (ADMN = 92% and ECN = 93%). CoUD patients showed a lower degree of overlap with HCs (ADMN = 66% and ECN = 65%), and greater decrease of network extension (ADMNNcv = 31,446 and ECNNcv = 31,286) than ADHD–CoUD patients (ADNNcv = 39,694 and ECNNcv = 40,373).
Posterior default mode network
PDMN was the most affected network in both groups (PDMNADHD-CoUD = 26%; PDMNcocaine = 37%), although with divergent changes. ADHD–CoUD patients showed a reduced network extension (Ncv = 7210) than HCs (Ncv = 16,561), and the CoUD group presented a more extended network (Ncv = 23,280) than HCs (Ncv = 16,561).
ATTN and BGN
ADHD–CoUD and CoUD patients showed a similar percentage of overlap with HCs in ATTN (ATTNADHD–CoUD = 44%; ATTNCoUD = 49%) and BGN (BGNADHD–CoUD = 51%; BGNCoUD = 46%).
SMN
ADHD–CoUD patients showed a higher percentage of overlap with HCs (39%) and higher network extension (Ncv = 18,763) than CoUD patients (26% of overlap with HCs and Ncv = 5872), suggesting a more preserved connectivity.
LIN
Despite both clinical groups showing a similar percentage of overlap with HCs (LINADHD–CoUD = 39% and LINCoUD = 44%), ADHD–CoUD patients presented a more extended network (Ncv = 9305) than CoUD patients (Ncv = 3073).
aSAN
ADHD–CoUD patients presented a more preserved aSAN than CoUD patients, considering both the degree of overlap with HCs (ADHD–CoUD = 71%; CoUD = 39%) and network extension (ADHD–CoUD = 35,641; CoUD = 18,972).
See Figure 4 and Supplementary Tables S5 and S6 in the Supplementary Data for IRCA results in graphical representations and indices.
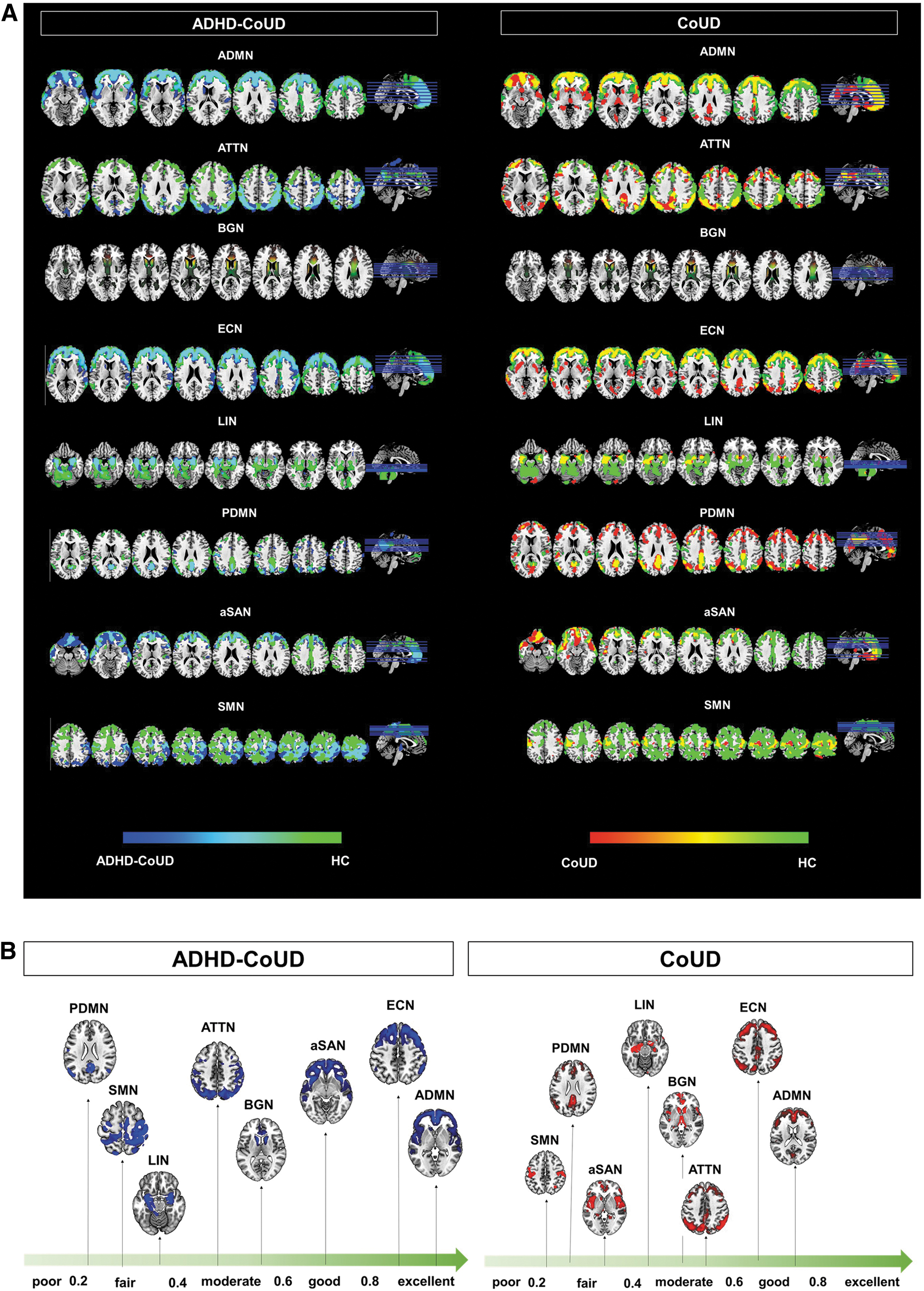
Large-scale brain network metabolic connectivity analyses.
Discussion
ADHD in adults showed high co-occurrence rates with CoUD. According to the SMH, ADHD patients might use drugs to suppress distressing symptoms. Cocaine seems to calm and counteract hyperactivity, emotional lability, and inattention in ADHD individuals (Mariani et al., 2014). Despite the SMH might help explaining the comorbidity of substance abuse in ADHD, its neurobiological correlates are not clarified yet.
This study explored the neurobiological mechanisms that could support SMH in adult patients with ADHD–CoUD by applying univariate and multivariate connectivity approaches to 18F-FDG PET data. We expected ADHD–CoUD patients to have altered brain mechanisms related to cocaine assumption within brain regions and networks involved in its core psychopathological domains (i.e., self-regulation of attention and behavior).
We found both detrimental and compensative effects of cocaine abuse in patients with ADHD–CoUD. Concerning detrimental cocaine effects, ADHD–CoUD patients showed a thalamic hypofunction associated with years of cocaine abuse. We also found possible compensative effects of cocaine use in ADHD patients: (1) more preserved dopamine networks (i.e., nigro-striato-cortical and mesocorticolimbic) than CoUD patients and (2) more preserved brain connectivity than CoUD patients in those large-scale networks involved in self-regulation mechanisms of attention and behaviors (i.e., ADMN, ECN, and aSAN) (Goulden et al., 2014; Sun et al., 2012).
These results provide insight into the neurobiological mechanisms, and support the SMH. The details of each main finding are discussed in the following paragraphs.
Both voxel-wise and ROI-based approaches revealed a specific thalamic hypofunction in ADHD–CoUD patients. Thalamus dysfunction is considered a key event in ADHD pathogenesis, leading to hyperactivity and inattention symptoms (Bailey and Joyce, 2015). We also found a significant correlation between number of years of cocaine abuse and lower metabolism in several thalamic divisions (i.e., prefrontal, visual, premotor, motor, and parietal) in the ADHD–CoUD group. A recent study demonstrated that cocaine abuse results in surface atrophy of the thalamus (Xu et al., 2023). Thus, the association between thalamus hypometabolism and total years of cocaine abuse might indicate a long-term detrimental effect of cocaine use on the thalamus in these patients.
However, we did not find the same relationship in the CoUD group. The explanation might lay in the higher vulnerability of this brain structure in ADHD–CoUD patients than in CoUD patients due to pre-existing thalamic dysfunctions (Bailey and Joyce, 2015). The combined effect of ADHD-related pathological mechanisms and long-term cocaine-induced neuroadaptations in glutamate transmission (Rouiller et al., 2003)—which plays a relevant role in modulating the activity of these regions (Sherman, 2011)—might cause the specific vulnerability observed in our cohort.
In ADHD–CoUD patients, the voxel-wise approach also revealed a higher metabolism than in CoUD patients and HCs in limbic regions (i.e., amygdala and parahippocampal gyrus bilaterally). The ROIs-based analyses confirmed these results. ADHD–CoUD patients showed higher metabolism than HCs and CoUD patients in the amygdala without reaching the statistical threshold with the latter group (Supplementary Table S4 in the Supplementary Data). These findings are coherent with previous data on amygdala dysfunctional activity in ADHD adults compared with HCs (Tajima-Pozo et al., 2018).
Amygdala dysfunction is associated with emotional dysregulation, significantly contributing to ADHD psychopathology during the lifespan (Corbisiero et al., 2013). The difference in amygdala activation between ADHD–CoUD and CoUD patients might reflect various neurobiological bases of emotional dysregulation in individuals with ADHD and SUDs. Impaired emotion regulation in SUDs is mainly associated with a hypoactivation of the rostral anterior cingulate cortex/ventromedial prefrontal cortex (Wilcox et al., 2016).
Accordingly, we found a more extended hypofunction in frontal cortices in CoUD patients than in ADHD–CoUD patients (Supplementary Fig. S1 and Supplementary Table S4 in the Supplementary Data). Notably, the frontal cortex receives massive dopaminergic projections, representing a central hub in the dopaminergic networks (Ciliax et al., 1999; Fallon, 1988). Consistently, metabolic connectivity analyses revealed a severe alteration in CoUD patients of nigro-striato-cortical, and mesocorticolimbic dopaminergic networks.
On the contrary, we found a preserved metabolic connectivity of the same dopaminergic networks in ADHD–CoUD patients (Fig. 3). These data might support the hypothesis that cocaine intake replaces the dopaminergic deficiency underlying ADHD clinical manifestations (Gold et al., 2014; Levy, 1991). Empirical studies showed that stimulant pharmacological treatments for ADHD increase the functional connectivity of frontoparietal networks associated with improving attentional performances (Wong and Stevens, 2012).
Consistently, the prolonged cocaine intake in ADHD patients might mimic the effects of stimulant pharmacotherapy. Of note, cocaine modulation of the dopaminergic system might represent the neurobiological substrate for the ADHD patients' perception of attentional improvement after cocaine use (Mariani et al., 2014).
ADHD–CoUD patients with a limited period of abstinence showed a higher metabolism in the vermis. The vermis has relevant implications for the turnover of dopamine and noradrenaline in the caudate and nucleus accumbens through its projections to these structures (Snider and Maiti, 1976). Furthermore, the cerebellar vermis showed morphological and functional alterations among individuals with ADHD, which were associated with motor symptoms (Paloyelis et al., 2007). Accordingly, vermis functioning has been selected as a key outcome for stimulant pharmacological treatments (e.g., methylphenidate), which exert robust effects on catecholamine systems in ADHD (Volkow et al., 2005).
Specifically, an 18F-FDG PET study demonstrated that a single dose of methylphenidate significantly increased cerebellar metabolism in ADHD individuals (Volkow et al., 1997). Thus, considering the stimulant pharmacological effects of cocaine on the dopaminergic system and its rapid pharmacokinetic (Cone, 1995), the improvement of vermis functionality with cocaine use in ADHD patients may suggest a possible compensatory effect on motor symptoms, according to clinical evidence and the SMH principles (Mariani et al., 2014).
Additional support for SMH came from IRCA results. Notably, patients with ADHD–CoUD showed more preserved connectivity than individuals with CoUD in ADMN, ECN, and aSAN. The better integrity of these networks in ADHD–CoUD patients suggests that cocaine may compensate for attention deficits in ADHD patients due to the crucial role of ADMN, ECN, and aSAN in attentive processes (Sun et al., 2012). Of note, previous studies showed the therapeutic effects of stimulant medications on the functional organization of these networks and their positive impacts on attentive symptoms in ADHD patients (Peterson et al., 2009).
All these neurobiological findings support the SMH of cocaine use among ADHD individuals based on a still spared neurobiological dopaminergic substrate, allowing the self-medication attempt to counteract the attentional symptoms linked to this condition.
Large-scale brain network analyses revealed a disruption of PDMN connectivity in ADHD–CoUD patients. This result could support the role of emotion dysregulation as a core feature of individuals with ADHD. Indeed, PDMN plays a crucial role in sustaining emotional evaluation and responsiveness (Vogt, 2014). Moreover, the disruption of PDMN might reflect a more pronounced vulnerability of patients with ADHD than with CoUD to cocaine-induced neuroadaptations linked to corticotropin-releasing factor activity that significantly impact the functional organization of this network (Koob and Le Moal, 2008). These results may also fit with a hypothalamic–pituitary–adrenal axis alteration, which is considered a stable biomarker of ADHD during the disease's development (Scassellati et al., 2012).
Overall, our results offer supporting evidence that ADHD–CoUD and CoUD patients are characterized by distinct neurobiological substrates, which might underpin different core psychopathological features of these conditions. Specifically, ADHD patients are characterized by pervasive deficits in self-regulatory mechanisms of motor activity, attention, and emotions (Barkley, 2022) linked to alterations of dopaminergic networks (Gold et al., 2014; Levy, 1991). Subjects with CoUD without a history of ADHD present core dysfunctions in reward-processing systems and related decision-making neural mechanisms (Luijten et al., 2017) that are associated with the dimension of impulsivity, especially considering the motor disinhibition domain (Kozak et al., 2019; Stevens et al., 2014).
Despite this evidence, we acknowledge some limitations. Although short-term abstinence maintenance allowed us to observe differential effects of cocaine within the clinical groups directly, future studies should replicate the current findings, including subjects with medium- and long-term abstinence (e.g., 1, 3, and 6 months) to confirm SMH principles among individuals with ADHD–CoUD. Moreover, the absence of a clinical control group composed of adult ADHD patients without SUDs and pharmacological treatment naive represented an additional limitation to our analyses.
Future research should also administer a more comprehensive assessment (e.g., neuropsychological and self-report) of core ADHD psychopathological domains (i.e., self-regulation of attention, behaviors, and emotions) to demonstrate how preserved and disrupted brain networks effectively might explain neurocognitive performances and emotional manifestations associated with this condition.
Nevertheless, this is the first study to provide neurobiological evidence for SMH in adults with ADHD–CoUD. The current results suggest differential treatment approaches, especially pharmacological approaches for CoUD individuals with and without ADHD, to improve functional adjustment, and reduce the risk of relapse in addictive behaviors. Specifically, stimulant pharmacological treatments (e.g., methylphenidate- and amphetamine-based stimulants) (Faraone and Glatt, 2009), especially with a long-acting formulation, should be considered the first line of intervention for adults with ADHD–CoUD to reduce symptomatology linked to self-regulation deficits of attention and behaviors and, in turn, reducing the probability of relapse in addictive behaviors (i.e., cocaine use and medication abuse) (Geffen and Forster, 2018).
Furthermore, these pharmacological treatments could be combined with evidence-based behavioral interventions for emotional dysregulation in patients with SUDs and ADHD (Cavicchioli et al., 2019; Hirvikoski et al., 2011), considering their implications for relapse prevention (Cavicchioli et al., 2020).
Footnotes
Authors' Contributions
G.C. contributed to study concept and design, analysis and interpretation of data, and drafting/revising the article; M.C. was involved in interpretation of data and drafting/revising the article; S.S., A.L.M., and C.M. carried out acquisition of PET data, study concept and design, interpretation of data, and revising the article; M.B. and A.M. were in charge of acquisition of clinical data, interpretation of clinical data, and revising the article; L.P. took charge of analysis of data and revising the article; and D.P. contributed to study concept and design, interpretation of data, and revising the article.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
The authors declare no financial funding.
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.