
Abstract
Background:
Youths with thought problems (TP) are at risk to develop psychosis and obsessive–compulsive disorder (OCD). Yet, the pathophysiological mechanisms underpinning TP are still unclear. Functional magnetic resonance imaging (fMRI) studies have shown that striatal and limbic alterations are associated with psychosis-like and obsessive-like symptoms in individuals at clinical risk for psychosis, schizophrenia, and OCD. More specifically, nucleus accumbens (NAcc) and amygdala are mainly involved in these associations. The current study aims to investigate the neural correlates of TP in youth populations using a dimensional approach and explore potential cognitive functions and neurotransmitters associated with it.
Methods:
Seed-to-voxels functional connectivity analyses using NAcc and amygdala as regions-of-interest were conducted with resting-state fMRI data obtained from 1360 young individuals, and potential confounders related to TP such as anxiety and cognitive functions were included as covariates in multiple regression analyses. Replicability was tested in using an adult cohort. In addition, functional decoding and neurochemical correlation analyses were performed to identify the associated cognitive functions and neurotransmitters.
Results:
The altered functional connectivities between the right NAcc and posterior parahippocampal gyrus, between the right amygdala and lateral prefrontal cortex, and between the left amygdala and the secondary visual area were the best predictors of TP in multiple regression model. These functional connections are mainly involved in social cognition and reward processing.
Conclusions:
The results show that alterations in the functional connectivity of the NAcc and the amygdala in neural pathways involved in social cognition and reward processing are associated with severity of TP in youths.
Impact Statement
Taking thought problems (TP) as a dimension, the multivariate linear regression model showed that the altered functional connectivity of the nucleus accumbens and amygdala is correlated to the severity of TP in youths. The replicability of identified connectivity was tested using an adult cohort. Functional characterization analyses revealed that these functional connectivities are involved in cognition (cognitive control), face detection (social cognition), and motivation/reward. Findings of the altered functional connectivity, related brain regions, and cognitive functions associated with TP can be a stepping stone to understanding the underlying pathophysiological mechanisms.
Introduction
In the past decade, a growing body of research investigated the interindividual differences in severity of mental disorders (First and Wakefield, 2013; Stein et al., 2021; Sumner et al., 2022; Wakefield, 2016; Wendt et al., 2022; Zimmerman et al., 2018), in order to overcome limitations of categorical-based classification systems (American Psychiatric Association, 2013). Many studies applied dimensional approach of constructs for assessments of mental disorders (Deyo and Langdon, 2021; Hart and Lewine, 2017; Mackinley et al., 2021; Stein et al., 2022; Sumner et al., 2022; Wendt et al., 2022). For instance, the Hierarchical Taxonomy of Psychopathology (HiTOP) model was proposed to emphasize the multi-dimensionalities of mental disorders while considering the comorbidity issue, including thought disorder as one of the six major spectra (Kotov et al., 2022). Early signs of thought problems (TPs) are defined as repetitive and negative thoughts about self, others, or situations (Farley et al., 2022; Klimes-Dougan et al., 2013), and typically include illusions/(pseudo-) hallucinations, obsessive-compulsive symptoms, and strange thoughts/behaviors (Abdellaoui et al., 2008). These symptoms are generically evaluated by an empirically derived set of items from the TP subscale of Child Behavior Checklist (CBCL-TP) (Achenbach and Rescorla, 2001) using a dimensional point of view (Ivarsson et al., 2008). TP symptoms characterize various mental disorders (e.g., schizophrenia, bipolar disease, and obsessive-compulsive disorder [OCD]), yet TP alone is not a nosology term in the Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition (DSM-V) (American Psychiatric Association, 2013). Despite the clinical significance of assessing these early manifestations, the pathophysiological mechanism underpinning TP remains not well understood (Abdellaoui et al., 2008; Klimes-Dougan et al., 2013; Salcedo et al., 2018).
Consistently with how they are defined (illusions, obsessive thoughts, etc.), TPs have been regularly associated with psychosis (Salcedo et al., 2018; Simeonova et al., 2014) and OCD (Anagnostopoulos et al., 2016; Ivarsson et al., 2008). In addition, psychotic-like symptoms are relatively common in patients with OCD (14%) (Eisen and Rasmussen, 1993; Okamura et al., 2022). On neurobiological grounds, it has been proposed that psychotic symptoms may arise from striatal dopaminergic dysregulation and the aberrant attribution of motivational/emotional salience to irrelevant stimuli (Griffin and Fletcher, 2017; Kapur, 2003; Winton-Brown et al., 2014). When the salience is incurred by stimulations, phasic dopamine is released subsequently, and the controls of prefrontal cortex toward the limbic system are weakened when there are multiple stimulations (Kapur, 2003; Winton-Brown et al., 2014). The dysregulation of dopaminergic pathway affecting brain regions, including prefrontal cortex and amygdala, could disrupt the priorities of saliences and, in turn, cause the aberrant salience (Howes and Nour, 2016; Winton-Brown et al., 2014). Aligned with this hypothesis, functional magnetic resonance imaging (fMRI) studies have shown that patients with schizophrenia display hyperactivations in the striatum (Ceaser and Barch, 2016), amygdala (Bourque et al., 2017; Dugré et al., 2019; Lukow et al., 2021; Pinkham et al., 2015), hippocampus (Dugré et al., 2019; Kätzel et al., 2020), and insula (Dugré et al., 2019; Walter et al., 2016) in response to neutral/ambiguous stimuli. Moreover, altered functional connectivity of amygdala has been implicated with schizophrenia development (Anticevic et al., 2014; Zhang et al., 2020). Dysfunctions of the amygdala, hippocampus, and striatum have been observed in individuals at clinical high risk for psychosis with altered salience processing (Bourque et al., 2017; Modinos et al., 2020). Higher aberrant salience observed in adolescents with TP is also potentially involved in higher susceptibility to psychosis (Lisi et al., 2021). Previous evidence suggests that evaluation of TP provides promising accuracy in identifying individuals with clinically significant psychosis-spectrum disorders among help-seeking, at-risk youths (Thompson et al., 2022). Taken together, these results show that striatal and limbic alterations are associated with psychotic symptoms in patients with schizophrenia and individuals at clinical risk for psychosis.
Patients with OCD often experience negative emotions such as anxiety in response to obsessive thoughts that may motivate them to engage in compulsive behaviors (Thorsen et al., 2018). Several fMRI studies have examined how these patients respond to stimuli that provoke negative emotions. Such studies found that during emotion processing, patients with OCD have increased activations of the amygdala and putamen, as well as anterior cingulate and frontal regions (Thorsen et al., 2018). Apart from these limbic alterations, there is now growing evidence showing that the brain reward system is involved in the pathophysiology of OCD. The brain reward system comprises the ventral tegmental area, the ventral striatum (nucleus accumbens [NAcc]), and the medial orbitofrontal cortex (mOFC) (Kim and Sul, 2023; Lewis et al., 2021). In theory, the brain reward system is thought to play a key role in OCD through abnormal reinforcement of repetitive behaviors adopted to prevent anticipated catastrophic outcomes (Alves-Pinto et al., 2019; Ferreira et al., 2017). Coherent with this model, the results of a meta-analysis of 47 resting-state functional connectivity studies have shown that the connectivity between the NAcc and fronto-limbic regions (e.g., insula and mOFC) is reduced in OCD (Liu et al., 2022). Adding to the validity of this model, several trials have shown that deep brain stimulation of the NAcc (NAcc-DBS) improves OCD symptoms (Schüller et al., 2022; Shi et al., 2023) in patients with severe and treatment-resistant OCD. The mentioned knowledge is mostly obtained from adult cohorts. To the best of our knowledge, the neural correlates of TP have rarely been directly assessed at the population level in adolescents. Two studies used Adolescent Brain and Cognitive Development (ABCD) dataset to investigate this question. Using the resting-state functional connectivity of this large dataset, Wainberg et al. (2022) showed that several neural networks, including the salience network, and also the cingulo-opercular, cingulo-parietal, default mode, dorsal attention, frontoparietal, and retrosplenial–temporal networks, are linearly associated with the TP dimension in children (Wainberg et al., 2022), whereas another analysis showed that cortico-striatal functional connections are important mediators of TP in children with insufficient sleep (Yang et al., 2022). Overall, these findings indicate that TP may be characterized by deficits in reward and emotion processing, with amygdala and NAcc as potential targets.
In view of the current state of knowledge, the present study aims to investigate the neural correlates of TP in child and adolescent populations using a dimensional approach. In this study, resting-state fMRI data obtained from 1360 children and adolescents were analyzed, and potential confounders related to TP such as anxiety and cognitive functions were included as covariates in multiple regression analyses. Considering that the striatum (NAcc) and amygdala have been implicated in the pathophysiology of both psychotic and obsessive–compulsive symptoms (which are the two main components of TP), we opted to perform seed-to-voxels analyses using the NAcc and amygdala as regions-of-interest. Furthermore, the replicability of the connectivity was tested in adult cohorts. To ease the interpretation of findings, functional decoding of identified altered functional connectivity based on these regions of interest was conducted through meta-analytic coactivation modeling. Exploratory analyses were also performed on the receptor/transporter density maps associated with the functional connectivity findings.
Materials and Methods
Participants: healthy brain network
Data from 2200 participants were obtained from the Healthy Brain Network (HBN), an ongoing initiative in the New York area (USA) that aims to investigate heterogeneity and impairment in developmental psychopathology (5–21 years old) (Alexander et al., 2017). The HBN adopted a community-referred recruitment model in which advertisements were provided to community members, educators, and parents. Exclusion criteria were impairments that prevented full participation in the study (e.g., serious neurological disorders, hearing or visual impairments), neurodegenerative disorder, acute encephalopathy, acute intoxication, and serious psychiatric disorders (recent diagnosis of schizophrenia and/or manic episode). Supplemental information is provided elsewhere (Alexander et al., 2017).
From the 2200 participants included in the Data Release 7.0, functional neuroimaging data were obtained from 1583 participants. Written assent was obtained from participants younger than 18 years old, and written consent was obtained from their legal guardians. Written informed consent was obtained from participants of age ≥18 prior to enrolling in the study. The original HBN study was approved by the Chesapeake Institutional Review Board (now Advarra Inc., see https://www.advarra.com/). The current study was approved by the Local Ethics Committee of Center intégré universitaire de santé et de services sociaux (CIUSSS) de l'Est-de-l'Île-de-Montréal.
Acquisition parameters
MRI acquisition took place at three different sites: mobile 1.5 T Siemens Avanto in Staten Island, 3 T Siemens Tim Trio at Rutgers University Brain Imaging Center (RUBIC), and 3 T Siemens Prisma at the CitiGroup Cornell Brain Imaging Center (CBIC) [acquisition protocols and parameters can be found in Supplementary Table S1 in Alexander et al. (2017), as well as at http://fcon_1000.projects.nitrc.org/indi/cmi_healthy_brain_network/]. Data at the CBIC were obtained using the same data acquisition protocol implemented at RUBIC. The acquisition of the two resting-state scans lasted 5 min each, during which participants viewed a fixation cross located at the center of the computer screen. Data for the Siemens Avanto were acquired in a single run lasting 10 min (details in Supplementary Table S1).
Main assessments
TP were evaluated using the Child Behavior Checklist (CBCL) (Achenbach and Rescorla, 2001), which comprised 15 items spanning psychotic-like experiences and obsessive–compulsive problems. Parents rated each item using a 3-point scale (0 = not true to 2 = very true) (α = 0.73).
We also examined whether functional connectivity results may be mediated by clinical and neuropsychological features, such as severity of anxiety [i.e., Parent-report Screen for Child Anxiety Related Disorders (BIRMAHER et al., 1999)] and conduct problems [i.e., CBCL (Achenbach and Rescorla, 2001)]. Executive functions in children were assessed using the NIH Toolbox Cognition Battery, including Dimensional Change Card Sorting and the Flanker tests (Zelazo et al., 2013). For both measures, age-adjusted scale scores were used.
fMRI data preprocessing
Functional images were realigned, corrected for motion artifacts with the Artifact Detection Tool (Power et al., 2014) (ART; setting a threshold of 0.9 mm subject to ART’s composite motion and a global signal threshold of Z = 5) with the implemented in CONN Toolbox (Whitfield-Gabrieli and Nieto-Castanon, 2012), bandpass filtered (0.01 Hz < f < 0.10 Hz), and coregistered to the corresponding anatomical image. The anatomical images were segmented (into gray matter, white matter, and cerebrospinal fluid) and normalized to the Montreal Neurological Institute (MNI) stereotaxic space. Functional images were then normalized based on structural data, spatially smoothed with a 6 mm full-width-at-half-maximum (FWHM) 3D isotropic Gaussian kernel and resampled to 2 mm3 voxels. For the preprocessing, the anatomical component–based noise correction method (aCompCor strategy) (Behzadi et al., 2007) was used to remove confounding effects from the blood-oxygen-level-dependent (BOLD) time series, such as the physiological noise originating from the white matter and cerebrospinal fluid. This method was found to increase the validity and sensitivity of analyses (Chai et al., 2012). As described recently, preprocessed images were manually checked for each of the 1583 participants (Dugré and Potvin, 2023). We found preprocessing issues due to the poor quality of images in 108 participants, which resulted in the software being unable to adequately detect and segment volumes into tissue classes (i.e., gray matter, white matter, and cerebrospinal fluid). In addition, 59 adolescents exhibited high movements (exceeding 3 mm), leaving a remaining sample of 1416 participants. Finally, given that the CBCL measures children and adolescent psychopathologies (<18 years), 56 adult subjects were excluded, leaving a final sample size of 1360 adolescents.
Statistical analyses
Seed-based connectivity analyses
The amygdala and the NAcc were selected as seeds of interest (i.e., FSL Harvard-Oxford Atlas distributed in the CONN toolbox). Physiological noise, realignment parameters, and movement artifacts were regressed as confounding effects from the BOLD time series at each voxel. In the first-level analysis, Pearson’s correlation coefficients were calculated between the residual BOLD time course from each seed and the time course of all other voxels, for each subject. Coefficients were converted to normally distributed z-scores using the Fisher Z-Transformation. Second-level analyses were conducted to identify functional connected voxels that were significantly correlated with severity of thought problems. Linear regression was conducted across the whole sample, while adjusting for age, sex, site, percentage of valid scans, and mean motion. A threshold of p < 0.001 uncorrected with a cluster extent threshold of 20 voxels was applied as a good trade-off (Woo et al., 2014) between a too liberal threshold (Lieberman and Cunningham, 2009) (p = 0.005) that may produce spurious results (Eklund et al., 2016) and a stringent cluster-level FWE, which inherently impact the cluster found in smaller brain regions. We nonetheless report clusters that remained statistically significant after applying cluster family-wise error (cFWE) < 0.05.
Subanalyses
Subanalyses were subsequently conducted using partial correlation, to examine the reliability of the relationship between functional brain connectivity and TP, while adjusting for age, sex, site, valid scans, and framewise displacement. First, we removed items of the CBCL-TP that were clinically specific (Lengua et al., 2001) to Psychotic-Like Experience (“Hears things not there”, “Sees things not there”, “Strange behavior”, and “Strange ideas”) (total of 12 items, α = 0.64) and to Obsessive–Compulsive problems (“Can’t get mind off thoughts”, “Repeats certain acts”, and “Stores up thing”) (total of 11 items, α = 0.66), to test whether results may have been driven by a particular clinical dimension. Differences in brain–behavior correlation between the subscales of the two scales were tested using Fisher r-to-z transformation (p < 0.05, two-tailed). Second, we added clinical (i.e., anxiety and conduct problems) and neuropsychological features (i.e., card sorting and flanker tests), as potential confounders in the partial correlation analyses.
Feature importance across developmental stages
After having identified significantly associated brain connectivity, we further investigated their importance in predicting TP. Feature importance was performed by conducting a multivariate linear regression, which included the resulting brain connectivity measures (independent variables) in association with the severity of TP. Importance of a particular feature is determined through a decrease in model R 2 score when a given feature is randomly shuffled (i.e., 5000 times) compared with the baseline model. Cross-validation was determined with Group KFold, using developmental stages (Early Childhood [6 to <8 years], Later Childhood [8 to <10 years], Early Adolescence [10 to <13 years], Middle Adolescence [13 to <16 years], and Late Adolescence [16 to <18 years]) as folds. Moreover, we removed the variance associated with confounds within each fold of the training data (i.e., age, sex, site, valid scans, framewise displacement) rather than on the whole data, as suggested recently (More et al., 2021; Snoek et al., 2019). We extracted changes in R 2 scores for each test sets to examine feature importance across developmental stages. Overall performance of the model was evaluated by a Pearson correlation between predicted and true values.
Replicability in adult sample
To assess the replicability of our results, we used the MPI-Leipzig Mind-Brain-Body (MPILMBB) dataset (Mendes et al., 2019). The MPILMBB includes 228 participants, including young- to middle-aged adults (between 20 and 40 years) and older adults (between 55 and 80 years), who performed a resting-state session for 15 min 30 sec (MRI acquisition sequences are detailed elsewhere,
Functional decoding
Functional decoding was conducted to better characterize the functional brain connectivity underpinning TP, based on the analysis of external data on task-based functions. The neurochemical correlates of functional connectivity alterations were also examined. To identify the neurocognitive functions characterizing a functional connectivity between two ROIs, we conducted a multi-ROI meta-analytic coactivation modeling (MACM). Briefly, we meta-analyzed studies from the BrainMap repository that reported at least one peak coordinate in both ROIs (i.e., seed and target). The BrainMap environment includes a repository of neuroimaging studies which contain brain coordinates and metadata (e.g., sample size, behavioral categories) for >21,083 experiments. An activation likelihood estimation (ALE) algorithm was then used to calculate spatial convergence across studies (Eickhoff et al., 2012) implemented in NiMaRE (Salo et al., 2022). The resulting ALE meta-analytic map (z-map) would thus reflect a task-based fMRI map in which both ROIs are coactivated.
After having identified the coactivation map of a given functional connectivity associated with TP, we calculated its spatial similarity with 13 data-driven task-based (tb)fMRI maps (see https://neurovault.org/collections/13769/, Dugré et Potvin, Preprint) and 19 receptor/transporter density maps (Hansen et al., 2022), which include serotonin (i.e., 5-HT1A, 5-HT1B, 5-HT2A, 5-HT4, 5-HT6, 5-HTT), dopamine (i.e., D1, D2, DAT), norepinephrine (i.e., NET), histamine (i.e., H3), acetylcholine (i.e., α4β2, M1, VAChT), cannabinoid (i.e., CB1), opioid (i.e., MOR), glutamate (i.e., NMDA, mGluR5), and GABA (i.e., GABAA/BZ). Spatial associations with the tbfMRI maps and the PET density maps were conducted using JuSpace (version 1.4) (Dukart et al., 2021). Briefly, the mean values of 728 brain regions which included 690 parcels of the Yale Brain Atlas (McGrath et al., 2022) and additional 38 subcortical and cerebellar regions from the AAL3 atlas (Rolls et al., 2020) were extracted for the two images. Partial correlation (Spearman’s rank correlation) adjusting for spatial autocorrelation (i.e., local gray matter probabilities) was then performed between the two sets. Exact permutation-based p-values (with 10,000 permutations) were computed and corrected using false discovery rate (FDR).
Results
Description of the HBN sample
The final sample comprised 1360 adolescents with a mean age of 10.79 (SD = 3.16). More than half of the sample was boys (62.1%). About half of the sample was White (47.9%), followed by bi/multiracial (17.5%), Black (16.5%), Hispanic (11.3%), and others. Percentage of valid scans after scrubbing was 86.6%, and mean motion was 0.35 (SD = 0.46). Raw CBCL-TP score was 3.51 (3.49) with an average T-Score of 59 (SD = 8.59).
Seed-based connectivity analyses
In children and adolescents, TP were negatively associated with functional brain connectivity of the left amygdala (seed) with the retrosplenial cortex, the posterior part of the superior temporal gyrus, and the secondary visual area (V2) (Fig. 1, Table 1). Moreover, TPs were positively associated with functional connectivity of the right amygdala (seed) to the right lateral PFC, but negatively with the right amygdala to the left caudate and to the left perigenual anterior cingulate cortex (Fig. 1, Table 1).
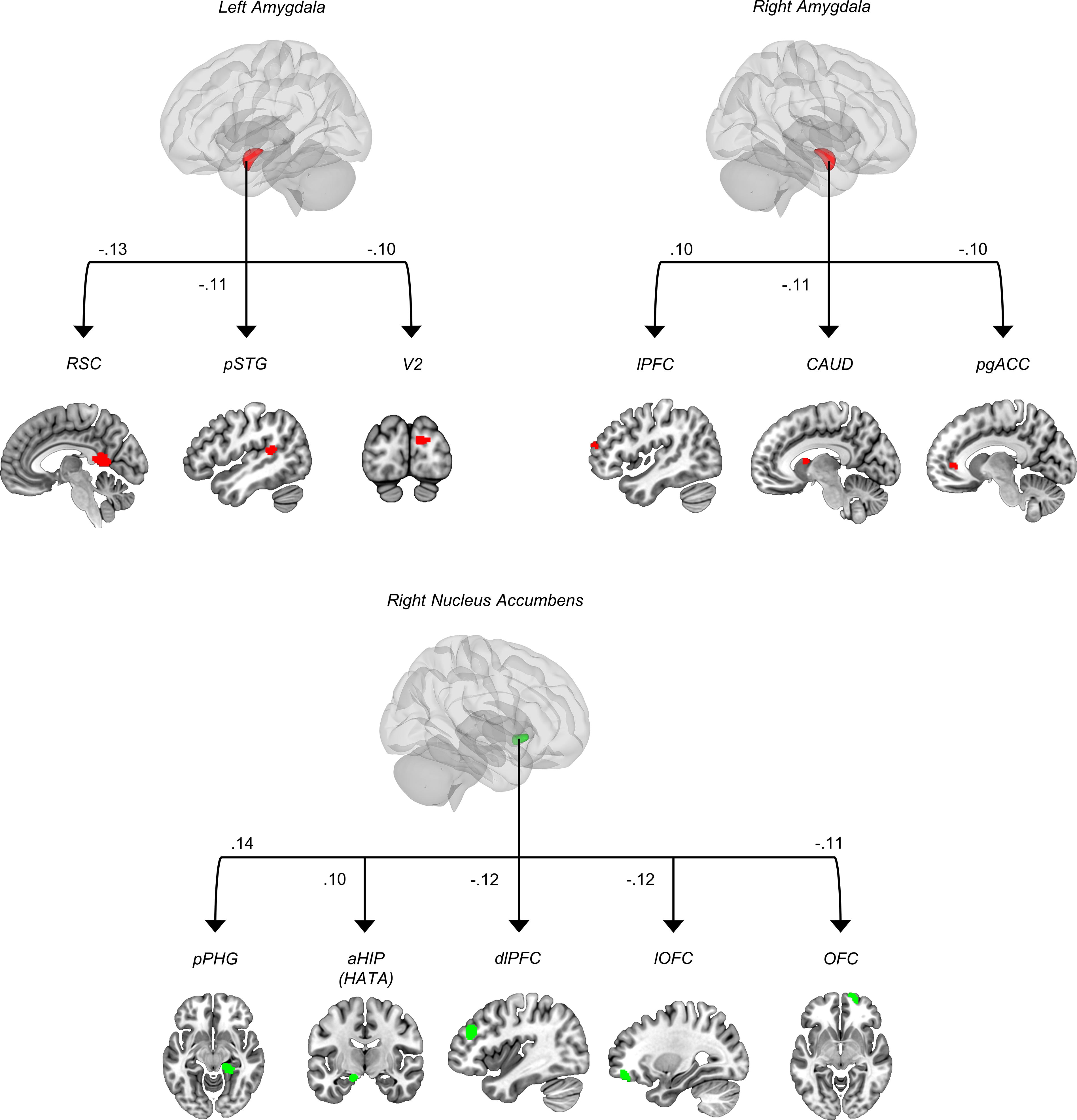
Seed-to-voxel functional connectivity associated with thought problems. RSC, retrosplenial cortex; pSTG, posterior superior temporal gyrus; V2, visual area 2; lPFC, lateral prefrontal cortex; CAUD, caudate nucleus; pgACC, perigenual anterior cingulate cortex; pPHG, posterior parahippocampal gyrus; aHIP, anterior hippocampus; HATA, hippocampal amygdala transition area; dlPFC, dorsolateral prefrontal cortex; lOFC, lateral orbitofrontal cortex; OFC, orbitofrontal cortex.
Seed-to-Voxel Functional Connectivity Results Associated with Thought Problems
p < 0.001 uncorrected; 20 voxels. Clusters surviving a cFWE < 0.05 threshold are marked by an asterisk (*).
STG, superior temporal gyrus; PFC, prefrontal cortex; ACC, anterior cingulate cortex; PHG, parahippocampal gyrus; HIP, hippocampus; HATA, hippocampal amygdala transition area; FP, frontopolar cortex; OFC, orbitofrontal cortex.
Using the NAcc as seeds of interest revealed that TPs were positively associated with functional connectivity of the right NAcc (seed) and the right posterior part of the parahippocampal gyrus and the left Hippocampal–Amygdala Transition Area (HATA)/anterior Hippocampus (Fig. 1, Table 1). TP were negatively correlated with connectivity from the right NAcc (seed) to the right dorsolateral prefrontal cortex, the right lateral orbitofrontal cortex, and frontopolar cortex/orbitofrontal cortex. No significant results were found for the left NAcc (Fig. 1, Table 1).
Subanalyses
We aimed to examine whether functional connectivity associated with the total TP could be driven by a particular clinical subconstruct (i.e., psychotic-like experiences and obsessive–compulsive problems). Comparing correlation coefficients with Fisher’s transformation revealed no significant differences across the 11 pairs of brain regions, suggesting that results are driven by a general TP construct. In addition, there is no significant difference on the relationship between brain connectivity and TP measurement across different age groups (age ≤ 10 years vs. age > 10 years) and scanner types (1.5 T vs 3 T). More specifically, the Z-test between participants with age ≤ 10 years and with age > 10 years of the brain connectivity between Left AMY-RSC, Right NAcc-dlPFC, Right NAcc-lOFC, and Right NAcc-pPHG were all statistically nonsignificant (details of Z-test results are listed in Supplementary Table S3). In addition, the relationship between brain connectivity and TP measurement did not differ between sex except the Right NAcc-FP, which showed greater negative correlation with TP in boys compared to girls (Z = 3.32, p < 0.001; details in Supplementary Table S4). Subanalyses suggested that some functional connectivity associations may be weaker in subjects scanned with a 1.5 T scanner (details in Supplementary Table S3).
Testing the effects of potential clinical confounders revealed a significant relationship between functional brain connectivity and anxiety and conduct problems, but not executive functions. After applying FDR correction for multiple comparisons, anxiety was significantly associated to (A) Left AMY-RSC, (B) Left AMY-pSTG, (C) Left AMY-V2, (D) Right AMY-lPFC, and (E) Right NAcc-lOFC. In turn, conduct problems were related to (A) Left AMY-pSTG, (B) Right NAcc-pPHG, and (C) Right NAcc-lOFC (Supplementary Table S2). Adding the severity of anxiety and conduct problems as confounders weakened some relationships with TP but remained statistically significant (Supplementary Table S2).
Feature importance across developmental stages
As shown in Figure 2, permutation feature importance revealed that across the developmental stages, functional connectivity between the right NAcc-pPHG (R 2 changes of 0.029), between the right amygdala–lPFC (R 2 changes of 0.022), and between the left amygdala–V2 (R 2 changes of 0.017) were the most important features in predicting TP, whereas the left amygdala–pSTG (R 2 changes of 0.004) and the right NAcc-lOFC (R 2 changes of 0.003) were the least.
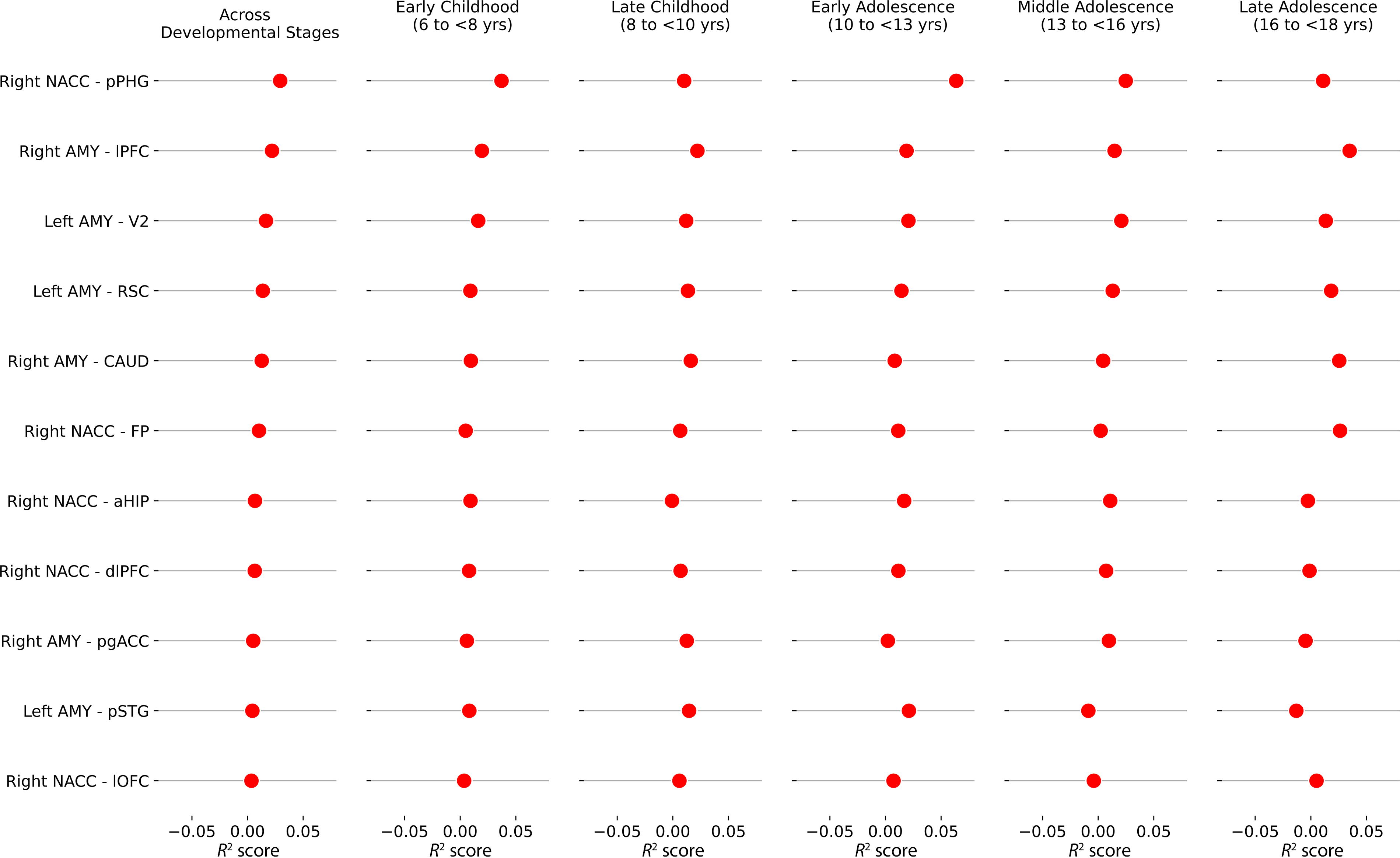
Importance of functional connectivity across developmental stages. NACC, nucleus accumbens; AMY, amygdala; RSC, retrosplenial cortex; pSTG, posterior superior temporal gyrus; V2, secondary visual area; lPFC, lateral prefrontal cortex; CAUD, caudate nucleus; pgACC, perigenual anterior cingulate cortex; pPHG, posterior parahippocampal gyrus; aHIP, anterior hippocampus; HATA, hippocampal amygdala transition area; dlPFC, dorsolateral prefrontal cortex; lOFC, lateral orbitofrontal cortex; OFC, orbitofrontal cortex.
Across developmental stages with age groups of 6 to <8 years, 8 to <10 years, 10 to <13 years, 13 to <16 years, and 16 to <18yrs, functional connectivity between the right amygdala–lPFC (0.022 ± 0.0077), left amygdala–V2 (0.0166 ± 0.004), and left amygdala–RSC (0.0137 ± 0.0033) showed lowest coefficient of variation of R 2 scores indicating stable relationship across developmental stages, whereas the functional connectivity between the left amygdala–pSTG (0.0044 ± 0.0148) highly varies from childhood to adolescence (peak in early adolescence and significantly reduced in mid- and late adolescence, details listed in the Supplementary Table S5).
Replication in adult samples
Partial correlation analyses were then carried out to examine specific relationships. Adjusting for the effect of age, sex, valid scans, and framewise displacement, only the left amygdala–V2 (partial r = −0.256, p = 0.017) and the left amygdala–pSTG (partial r = −0.22, p = 0.04) were associated with severity of TP in adults.
Functional decoding
Functional decoding analyses (Fig. 3) were performed for connectivity between (1) the right NAcc and the pPHG, (2) the right AMY and lateral PFC, and (3) the left AMY-V2, as these were found to be the most strongly correlated features of TP across developmental stages.

Functional characterization of seed-to-voxel results significantly associated with thought problems.
Functional decoding analyses for the right NAcc–pPHG connectivity revealed that coactivation pattern (including the left NAcc, anterior midcingulate cortex, bilateral amygdala, and fusiform gyrus) mainly correlated with face detection (z’ = 0.48, pFDR < 0.05), Motivation (z’ = 0.47, pFDR < 0.05), as well as with dopamine transporter (z’ = 0.61, pFDR < 0.05) and serotonin transporter (z’ = 0.61, pFDR < 0.05) density maps.
Coactivation network of the right amygdala and the lateral PFC (i.e., right lateral PFC, left amygdala, thalamus, anterior midcingulate cortex, dorsolateral PFC, anterior insula) was mostly associated with multiple demand (z’ = 0.61, pFDR < 0.05) and cognitive control (z’ = 0.58, pFDR < 0.05), as well as with VAChT (z’ = 0.39, pFDR < 0.05), dopamine transporter (z’ = 0.34, pFDR < 0.05), and D2 (z’ = 0.33, pFDR < 0.05).
Finally, the coactivation pattern of the left amygdala and the V2 (i.e., right amygdala/anterior hippocampus, dorsomedial PFC, fusiform gyrus, ventrolateral PFC) was significantly associated with face detection (z’ = 0.39, pFDR < 0.05) and both dopamine transporter (z’ = 0.34, pFDR < 0.05) and serotonin transporter (z’ = 0.31, pFDR < 0.05) density maps.
Discussion
The present study investigated the functional connectivity associated with TP in children and adolescents (age under 18 years) using the amygdala and NAcc as seed regions. TPs were positively correlated with the functional connectivity between the right amygdala and right NAcc, whereas TPs were negatively associated with the functional connectivity of left and right amygdala and right NAcc. The best predictors of TP in this study were the functional connectivity between the right NAcc and pPHG, between the right amygdala and lPFC, and between the left amygdala and V2. Two (the right amygdala–lPFC, the left amygdala–V2) out of those three functional connectivities presented high consistency (low variation of R 2 coefficients) across the five developmental stages. Moreover, one of those three functional connectivities (the left amygdala–V2) was found to be positively correlated with the severity of TP in adults.
Amygdala is a core structure of limbic system and is well known for its involvement in processing fear (Becker et al., 2012), emotional memories (Richardson et al., 2004), addiction (Ko et al., 2023; Koob, 2009), and social interaction (Becker et al., 2012; Gupta et al., 2011). Intense negative emotions such as irritability or fear can be induced by electric stimulation of the amygdala (da Cunha-Bang et al., 2019; Domínguez-Borràs et al., 2020). Lesions of the amygdala could reduce response of fearful stimulations in humans (Domínguez-Borràs et al., 2020) and potentially contribute to flattening of affect (Bliss-Moreau et al., 2011; Katayama et al., 2009). As for the lateral PFC, a large fMRI literature has shown that it plays a key role in superior executive functions, such as cognitive flexibility and planning skills (Axelsson et al., 2021; Fujino et al., 2017; Jones and Graff-Radford, 2021). Coherently with these findings, functional decoding analyses showed that the functional connectivity between amygdala and lateral PFC was associated with multiple demand and cognitive control. Alteration of functional connectivity between amygdala and lateral PFC has been reported to be associated with childhood adversity (Cheng et al., 2021), bipolar I disorder (Townsend et al., 2013), and it is a candidate endophenotype of schizophrenia (Quarto et al., 2018). The neural connection between the amygdala and lateral PFC is bidirectional with participation of both excitatory and inhibitory neurons and involves hippocampal afferent signals to both amygdala and prefrontal cortex, as well as efferent signal sent from lateral PFC to amygdala and vice versa (Sigurdsson and Duvarci, 2016; Yang and Wang, 2017). In rodents, dopamine, GABA, glutamate, and acetylcholine were neurotransmitters reported to be involved in these neuronal circuits (Sigurdsson and Duvarci, 2016; Tsolias and Medalla, 2022; Wang et al., 2020; Yang and Wang, 2017). Here, we found that the coactivation maps of the functional connectivity between the amygdala and lateral PFC were correlated with the density maps of VAChT, DAT, and D2 receptors, consistently with the previous preclinical literature (Dixon, 2015; Duncan, 2010).
For the functional connectivity between amygdala and V2, the preclinical literature has shown that both regions are involved in fear conditioning and has highlighted that there is an indirect neuronal pathway from V2 to amygdala, whereby visual information is first processed by V1, then transmitted to V2, and then from V2 to the amygdala (Shi and Davis, 2001). In our study, functional decoding analyses showed that the coactivation maps of the amygdala-V2 functional connectivity were mostly associated with face detection, which is a social cognitive function involved in the early processing of social threat detection (Schellhaas et al., 2022). As such, these results are consistent with the face recognition deficits that have been observed in schizophrenia (Maher et al., 2019; Maher et al., 2016; McBain et al., 2010). Interestingly, one of the symptoms that individuals with TP may experience is consistent, repetitive, and negative thoughts (Farley et al., 2022). These can be triggered by negative emotions (i.e., anxiety, depression, or stress), including fear (Minor et al., 2015). As for the analyses on transporter/receptor density maps, they revealed that the coactivation map of the amygdala–V2 functional connectivity was associated with the DAT and serotonin transporter (SERT). The preclinical literature has clearly shown that both dopamine and serotonin can modulate basic negative emotions through the amygdala (Dannlowski et al., 2013; McDonald, 2023). Moreover, preclinical cytoarchitectonic studies (Rapan et al., 2022) and experimental studies on serotonergic psychedelic drugs (e.g., psilocybin) have shown that serotonin receptors (e.g., 5-HT1A) are found in significant concentrations in the occipital lobe (including V2) and are involved in visual perception (Castelhano et al., 2021; Rocha et al., 2019). Other than the fact that dopamine receptors do not play a key role in occipital-related functions, our results are generally consistent with the available evidence.
The NAcc is mainly involved in reward processing (Hikida et al., 2016), motivation (Floresco, 2015), and reinforcement learning (Gale et al., 2014), whereas the pPHG is mostly involved in memory encoding and retrieval, spatial navigation, and scene recognition (Bastin et al., 2013; Burgmans et al., 2011). Alteration of NAcc activity is associated with amotivation and anhedonia (Li et al., 2018). The parahippocampal gyrus mediates cortico-hippocampal communication and plays an important role in the regulation of memory, including emotional memories, which are often formed in the context of social interactions (Burgmans et al., 2011; Mikels and Reuter-Lorenz, 2019; Rolls, 2019). The NAcc receives input signals (afferent) and projects output signals (efferent) from and to the hippocampal formation for the integration of reward-related contextual information and memory processes (Makris et al., 2008). Coherently with the known separate roles of the NAcc and parahippocampus, the functional decoding analyses showed that the coactivation maps of the functional connectivity between both regions were mainly associated with face detection (e.g., social cognition) and motivation (reward). Moreover, the coactivation maps of the NAcc-pPHG functional connectivity were correlated to the dopamine and serotonin transporters. These results are consistent with the preclinical literature indicating that both NAcc and parahippocampal gyrus receive dopaminergic and serotonin inputs (Boulenguez et al., 1996; Iritani et al., 2006; Kärkkäinen et al., 2015; Raslau et al., 2015).
In our study, resting-state functional imaging data were collected in >1300 youths (under 18 years). The identified functional connectivity associated with severity of TP remained statistically significant when taking potential confounders such as severity of anxiety, conduct problems, and children’s executive functions (i.e., card sorting and flanker tests) into account. In terms of replicability, the functional connectivity between the left amygdala–V2 was also found to be associated with TP in adults. Moreover, the presented study is one of the first resting-state functional connectivity studies on the TP dimension in young individuals. Yet, there are a few limitations that need to be acknowledged. First, there was a large range of age (6–18 years) across the collected dataset. This could potentially incur the heterogeneity to the data, especially for the cohorts in developmental and adolescent phases. However, in two out of three, the most predictive functional connectivity identified in our study presented the lowest variation across the developmental stages. Second, the CBCL-TP subscale measuring the TP dimension combines OCD-like and psychotic-like symptoms (Lengua et al., 2001). In the dimensional view of mental disorders such as the HiTOP model, OCD-like and psychotic-like symptoms are defined in different spectra (i.e., internalizing and thought disorder spectra) with different dimensions associated with them (Kotov et al., 2022). However, it is noteworthy that stratification of dimension for mental disorders might not be the same between adults and young individuals (age <18 years). On the one hand, there is a continuum between OCD-like and psychotic-like symptoms (Abdellaoui et al., 2008). On the other hand, the developmental periods of childhood and early adolescence may be too early to differentiate between these symptoms. This inconsistency will need to be addressed in future research. In a large sample of youth, this study adopted a dimensional approach to the study of the functional alterations associated with early manifestations of TP. As such, this current study is the first of this kind. Our main results showed that the severity of TP was correlated with the functional connectivity between the right amygdala and lateral PFC, between the left amygdala and occipital cortex, and between the right NAcc and pPHG. Functional characterization analyses revealed that these functional connectivities are involved in cognition (cognitive control), face detection (social cognition), and motivation/reward. Future studies on the neural mechanisms involved in the development of TP will need to adopt longitudinal designs, with follow-ups until late adolescence and early adulthood. In these investigations, attention will need to be paid to the potential commonalities and differences between OCD-like and psychotic-like symptoms.
Footnotes
Acknowledgments
S.P. is holder of the Eli Lilly Canada Chair on schizophrenia research; J.R.D. is holder of a postdoctoral fellowship from the Canadian Institutes of Health Research (MFE-181885).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study received no external funding.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.