
Abstract
Purpose:
Rhyming is a phonological skill that typically emerges in the preschool-age range. Prosody/rhythm processing involves right-lateralized temporal cortex, yet the neural basis of rhyming ability in young children is unclear. The study objective was to use functional magnetic resonance imaging (fMRI) to quantify neural correlates of rhyming abilities in preschool-age children.
Method:
Healthy pre-kindergarten child–parent dyads were recruited for a study visit including MRI and the Preschool and Primary Inventory of Phonological Awareness (PIPA) rhyme subtest. MRI included an fMRI task where the child listened to a rhymed and unrhymed story without visual stimuli. fMRI data were processed using the CONN functional connectivity (FC) toolbox, with FC computed between 132 regions of interest (ROI) across the brain. Associations between PIPA score and FC during the rhymed versus unrhymed story were compared accounting for age, sex, and maternal education.
Results:
In total, 45 children completed MRI (age 54 ± 8 months, 37–63; 19M 26F). Median maternal education was college graduate. FC between ROIs in posterior default mode (imagery) and right fronto-parietal (executive function) networks was more strongly positively associated with PIPA score during the rhymed compared with the unrhymed story [F(2,39) = 10.95, p-FDR = 0.043], as was FC between ROIs in right-sided language (prosody) and dorsal attention networks [F(2,39) = 9.85, p-FDR = 0.044].
Conclusions:
Preschool-age children with better rhyming abilities had stronger FC between ROIs supporting attention and prosody and also between ROIs supporting executive function and imagery, suggesting rhyme as a catalyst for attention, visualization, and comprehension. These represent novel neural biomarkers of nascent phonological skills.
Impact Statement
This is the first study using magnetic resonance imaging (MRI) and connectivity-based analyses to describe relationships between functional brain networks and nascent rhyming ability in preschool-age children listening to stories. The research question is significant on a mechanistic level, as rhyming is among the first phonological skills to develop in support of subsequent language and reading abilities and also in terms of early interventions designed to enhance these abilities. The findings are consistent with MRI research involving the neural signature of prosodic processing in older children and adults and also with behavioral–educational evidence regarding the appeal of rhymed stories at this age.
Introduction
Rhyming is an early phonological (language) skill involving correspondence of ending sounds between words (Moats, 2009). Mastery evolves from perception of such word sounds (e.g., during story listening), through enjoyment of them (e.g., rhyming songs), and, ultimately, production of rhymes de novo (Anthony et al., 2003). Rhyming abilities also often involve attention to prosody, which is defined as patterns of rhythm in sounds, such as music, songs, or stories (Wolters et al., 2022). Indeed, the rhythmic cadence of rhymed stories and songs is a substantial part of what helps make them so fun, memorable, and popular during early childhood, exemplified by classical nursery rhymes and the “ABC Song.”
Rhyming is among the earliest phonological skills to emerge, typically between 3 and 5 years old, influenced by genetic and environmental factors (Xie, 2019). It has been called a “constrained” cognitive ability, in that emergence is typically in a finite, binary window, as opposed to “unconstrained” skills such as vocabulary, which develops incrementally with no upper bound (Paris, 2005). Overall, phonological abilities have been estimated to be 45% heritable and tend to persist across the lifespan (Grigorenko, 2004) though are modifiable through interventions (Torgesen et al., 1999), ideally during early childhood when neural plasticity is high (Gilmore et al., 2018; Knudsen, 2004). Indeed, low to absent rhyming ability can be an early sign of language and subsequent reading difficulties, particularly those with a phonological basis (e.g., dyslexia), and is thus an appealing potential target for early screening and intervention (Olson et al., 1989).
Magnetic resonance imaging (MRI) is a powerful tool that can provide insights into the neurobiological basis of language and other cognitive abilities and how these are impacted by experiences and interventions, complimenting behavioral evidence (Black et al., 2015; Vandermosten et al., 2016). Functional MRI (fMRI) tasks utilized in language and literacy research include active response (e.g., verb generation, detection of word sounds) (Holland et al., 2001; Hurschler et al., 2015) and passive stimuli (e.g., stories listening) (Vannest et al., 2014). Numerous MRI studies have characterized structural and functional neurobiological correlates of oral language abilities (Holland et al., 2007; Price, 2012; Vannest et al., 2009), as well as the process by which brain areas supporting language, visual, and other abilities are integrated to form a functional “reading network” (Horowitz-Kraus and Hutton, 2015). MRI has also been utilized to quantify differences in brain structure and function related to home literacy environment and story format (e.g., illustrated, animated) in preschool-age children (Hutton et al., 2019b; Hutton et al., 2020c). However, given technical challenges (e.g., cooperation with fMRI tasks, motion), the majority of pediatric studies involving the neural basis of language and literacy skills have involved older children and young adults (Bitan et al., 2007; Blumenfeld et al., 2006; Cao et al., 2009; Hoeft et al., 2007a; Hoeft et al. 2007b).
The neural basis of phonological abilities and their disruption in language and reading difficulties (e.g., dyslexia) has been described in school-age children (Kovelman et al., 2012a; Kovelman et al., 2012b; Mathur et al., 2020; Wang et al., 2021). Tasks utilized include passive listening (e.g., word-rhyme judgment) and active (e.g., rhyme generation) responses (Lurito et al., 2000; Cao, 2017). Brain areas most often identified in this work include left-lateralized inferior frontal gyri (IFG) and superior temporal gyri (STG) (Mathur et al., 2020) and also left-superior frontal, middle temporal gyri (MTG), bilateral frontal, bilateral middle frontal, and bilateral fronto-temporal gyri (Booth et al., 2004; Cao et al., 2008; Cao et al., 2006; Hoeft et al., 2007a; Hoeft et al., 2007b). An fMRI study involving typically developing 5- to 6-year-olds performing an auditory word-rhyming task identified recruitment of the left Dorsolateral Prefrontal Cortex (DLPFC) (Kovelman et al., 2012a; Kovelman et al., 2012b), thought to support rhyme awareness (i.e., detecting whether words rhyme or not). In contrast to language-specific research, a neural signature for prosody detection in 5- to 7-year-old children was mapped to the R-hemisphere, mirroring left-sided phonological areas (Chen et al., 2022; Kovelman et al., 2012a; Kovelman et al., 2012b; Plante et al., 2006). This is consistent with fMRI literature involving adults that has consistently mapped prosodic processing to the R-hemisphere, especially frontal-temporal areas (Friederici and Wartenburger, 2010; Kreitewolf et al., 2014; Sammler et al., 2018). However, the neurobiological basis of nascent prosodic and/or rhyming abilities in young children before kindergarten entry has not previously been studied.
The objective of the current study was to explore relationships between emergent rhyming abilities and functional connectivity (FC) within and between atlas-defined brain networks (e.g., language, attention) in a sample of healthy, typically developing, preliterate preschool-age children. It used a novel, continuous (long-block) story listening paradigm, where age-appropriate rhymed and unrhymed stories were presented in their entirety during fMRI scanning. Given its role in prosody detection, the hypothesis was that FC involving right-sided temporal and inferior frontal areas would be greater during the rhymed story, where such rhythms occur, possibly reflecting a neural signature of nascent rhyme awareness. Furthermore, we hypothesized greater engagement of the dorsal (top-down) attention network during the rhymed story, given the appeal of rhymed stories at this formative age (Johnson and Hayes, 1987; Király et al., 2017; Read et al., 2014; Read, 2014).
Materials and Methods
Overview/design
The current study is a secondary analysis of data collected for a cross-sectional, MRI-based study involving impacts of home reading practices and digital media use on brain structure and function supporting emergent literacy skills in preschool-age children (Hutton et al., 2020b; Hutton et al., 2020a).
Participants/setting
Healthy children between 3 and 5 years old were recruited at a pediatric academic center and primary care clinics in a large Midwestern city. Eligibility criteria were (1) gestation ≥36 weeks, (2) age 36–52 months, (3) no prior or current kindergarten attendance, (4) no documented history of head trauma with loss of consciousness or neurodevelopmental condition likely to confer cognitive delay, (5) native English-speaking custodial parent, and (6) no contraindications for MRI such as metal implants, orthodontic braces, or claustrophobia.
Ethics statement
Written informed consent was obtained from a parent before all study procedures. The consenting parent was provided with reasonable financial compensation (ClinCard) for time and travel. This study was approved by the Cincinnati Children's Institutional Review Board. All research was performed in accordance with human subjects’ protection guidelines in accordance with the Declaration of Helsinki principles.
Rhyming measure
The Preschool and Primary Inventory of Phonological Awareness (PIPA) is a direct assessment of phonological skills for ages 3.0–6.9 years old, including children who are preliterate. This has six subtests (estimated 5 min each), five of these designed to assess aspects of sound awareness and the sixth-letter-sound knowledge (Dodd et al., 2003). For the current study, only the rhyme awareness (rhyming) subscale was administered. Scoring generates a raw and standardized score, percentile rank, and proficiency level (below basic, basic, and proficient).
Other behavioral measures
Clinical research coordinators collected demographic information and administered the ScreenQ to the child’s parent in a private room before the MRI scan. Other established measures of language (Expressive Vocabulary Test, 2nd Edition; EVT-2), processing speed (Comprehensive Test of Phonological Processing, rapid object naming subscale; CTOPP-2), and composite emergent literacy skills [The Reading House (Hutton et al., 2019)] were also administered to the child before MRI and have been reported previously (Hutton et al., 2020b; Hutton et al., 2020a).
Following MRI, the child was administered two 5-item, multiple-choice quizzes developed by the study team (both estimated kindergarten reading level), regarding factual content in each of the two stories presented in the scanner. Each quiz had a total score ranging from 0 to 5 points.
Descriptive analyses
Descriptive statistics were computed for demographic and other variables, specified in a statistical analysis plan. Poverty status was defined using 2020 U.S. Department of Health and Human Services criteria, using the midpoint of income category relative to household size (U.S. Department of Health and Human Services, 2020). Analyses were conducted using SAS v9.4 software.
MRI
Details of play-based acclimatization techniques before MRI are described previously (Hutton et al., 2015). The protocol involved structural and functional MRI, the latter including resting state and active tasks. For the current study, a novel fMRI paradigm was developed, involving two age-appropriate, published stories of similar reading difficulty (Lexile level, MetaMetrics®), theme, and length, which were read aloud in their entirety in a sound booth by the same member of the study team and recorded. The unrhymed story was The Big Orange Splot (Daniel Pinkwater, 1977, Scholastic), and the rhymed was The Strange Shirt Spot (Dr. Seuss, 1951, Random House). Spacing was added in these recordings in customary locations (e.g., between page spreads) using audio editing software to make them the same length for use in the paradigm (3:31 min). These were then incorporated into MRI using ePrime software (Psychology Software Tools, Pittsburgh, PA). At the MRI visit, children were asked if they had heard either story before to account for this potential confounding variable. No children responded affirmatively.
Children were awake and nonsedated during MRI, which was conducted using a 3-Tesla Philips Ingenia scanner with a 32-channel head coil. High-resolution, 3D T1-weighted anatomical images were acquired for registration to a standard template (Repetition Time (TR)/Echo Time (TE) = 8.1/3.7 msec; duration 5.25 min; Field of View (FOV) = 256 × 256 mm; matrix = 256 × 256; in-plane resolution = 1 × 1 mm; slice thickness = 1 mm; number of slices = 180, sagittal plane). Blood oxygen level dependent (BOLD) fMRI data used in the current study were continuously acquired for just over 7 min using a spin-echo echo planar imaging sequence (repetition time = 1 sec, echo time = 30 ms, flip angle = 62 degrees, 2.5 mm in-plane resolution, 3 mm slice thickness, multiband factor = 4), during which children listened to the two stories (rhymed and then unrhymed, 3:31 min each) in their entirety. The child was asked to verify suitable audio quality in the scanner before story presentation. Five seconds with no audio stimulus separated the two-story conditions. The only visual stimulus presented during the story conditions was a black fixation cross (+) on a white background.
MRI analyses
BOLD fMRI data were spatially and temporally preprocessed using the CONN FC toolbox, version 21a (Whitfield-Gabrieli and Nieto-Castanon, 2012). BOLD data were rigidly co-registered across time and brought into alignment with the 3D T1-weighted image. Nonlinear registration to MNI152 space was performed using SPM12’s Unified Segmentation Algorithm (Ashburner and Friston, 2005) with age-matched tissue probability priors (Wilke et al., 2008). Temporal preprocessing and denoising included (1) regression of framewise motion parameters (3 rotations, 3 translations) and their first-order derivatives from the BOLD signal, (2) regression of the first five principle components of the BOLD signal from each of the white matter and cerebrospinal fluid compartments (Muschelli et al., 2014), (3) censoring of motion-contaminated outlier frames, identified as having >0.9 mm framewise displacement OR having global signal intensity >5 standard deviations above the mean, (4) regression of the hemodynamic-response convolved story onset and durations, and (5) bandpass filtering (0.008–0.09 Hz).
FC was computed as the Fisher-transformed Pearson correlation coefficient of the average residual. BOLD signal was calculated from each of 132 anatomically delineated regions of interest (ROI) spanning the whole brain; 106 cortical and subcortical ROIs were defined by the Harvard–Oxford atlas (Makris et al., 2006), whereas 26 cerebellar ROIs were defined by the Automated Anatomical Labeling atlas (Tzourio-Mazoyer et al., 2002). The CONN toolbox comes preinstalled with these 132 ROI definitions as well as grouping information dividing the ROIs into 22 a priori functional networks. The groupings were determined using complete-linkage hierarchical clustering and optimal leaf ordering (Bar-Joseph et al., 2001) applied to Cambridge 1000-connectomes resting state dataset (Biswal et al., 2010). Use of these network definitions, derived from connectivity patterns of an independent dataset, should reduce selection and circularity bias and also facilitate any reproducibility studies by virtue of being readily available to independent researchers. A full listing of these network definitions is provided in Supplementary Table S1. Of note, 4 of the 22 networks comprise our significant results, listed here for the reader’s reference: the posterior default mode network (pDMN), the right lateralized fronto-parietal network [FPN (R)], the right lateralized language network [Lang (R)], and the dorsal attention network (DAN). The ROIs belonging to these four networks are (1) the posterior cingulate gyrus and precuneus in the pDMN; (2) the right frontal pole, the right superior frontal gyrus, and the right middle frontal gyrus in the FPN (R); (3) the pars opercularis of the right inferior frontal gyrus, the temporo-occipital region of the middle temporal gyrus, and the posterior division of the right supramarginal gyrus in the Lang (R); and 4) the temporo-occipital part of the right and left inferior temporal gyri and the right and left superior parietal lobules in the DAN.
Parametric multivariate cluster-level statistical inferencing was performed (Jafri et al., 2008) to test the null hypothesis that associations between PIPA and FC were equal during rhyming and non-rhyming story block listening, controlling for age, sex, and maternal education level. Maternal education was chosen as a proxy for socioeconomic status (SES), as it has been cited as most strongly associated with child cognitive and social–emotional development (Jackson et al., 2017). Cluster-level inferencing was performed on the 22 a priori networks described earlier and detailed in Supplementary Table S1. To account for multiple comparisons testing, false discovery rate (FDR) correction was applied for all analyses using thresholds of α = 0.05, with a two-sided test.
Results
Sample characteristics and PIPA scores
A total of 45 children completed fMRI with acceptable image quality for these analyses, applying criteria described earlier (age 53.7 ± 7.7 months-old, range 37–63; 26 girls, 19 boys). None of the children reported having heard either story before, which was affirmed by their accompanying parent. The mean total PIPA score (rhyming subscale) for those completing fMRI was 5.8 (± 3.5; range 1–12), which corresponded to skill levels of 9 Emerging/Below Basic, 23 Basic, and 13 Proficient.
Demographic and cognitive data are summarized in Table 1.
Demographics and Rhyming Measure Scores
Poverty status determined using 2022 U.S. Department of Health and Human Services income-to-household size criteria.
PIPA, Preschool and Primary Inventory of Phonological Awareness; SD, standard deviation.
Cognitive–behavioral analyses
Higher total PIPA rhyming scores were positively associated with child age (p < 0.001) and household income level (p < 0.05) but not significantly associated with child sex or maternal education level. PIPA rhyming scores were also positively associated with scores on other language and emergent literacy measures administered at the visit (EVT-2 vocabulary, CTOPP rapid object naming, Get Ready to Read emergent literacy composite, The Reading House emergent literacy composite), which have been published previously (Hutton et al., 2020b; Hutton et al., 2020a).
Story comprehension
The mean total score on the comprehension quiz for the rhymed story was 2.8 ± 1.1 (1–4) and for the non-rhymed story was 1.9 ± 1.3 (0–4). The score for the rhymed story was significantly higher (paired t-test, p < 0.001). Scores on both quizzes were positively correlated with total PIPA rhyming subscale score (p < 0.05).
MRI analyses
Controlling for an FDR at α = 0.05, two clusters exhibited significantly greater, positive associations between PIPA score and FC during the rhyming block compared with the unrhymed block: FC between the right-lateralized language network and DAN [F(2,39) = 9.85, p-FDR = 0.044] and also the posterior default mode network (DMN) and the right FPN [F(2,39) = 10.95, p-FDR = 0.043]. These findings remained statistically significant controlling for demographics child age, sex, and maternal education level and are shown in Figure 1 and Figure 2, respectively. Between the right-lateralized language network and DAN, post hoc tests revealed significant positive associations with PIPA scores in 6 out of 12 between-network connections. Between the posterior DMN and right lateralized fronto-parietal network, all six between network connections exhibited significant positive associations with PIPA score. These findings are summarized in Figure 3.
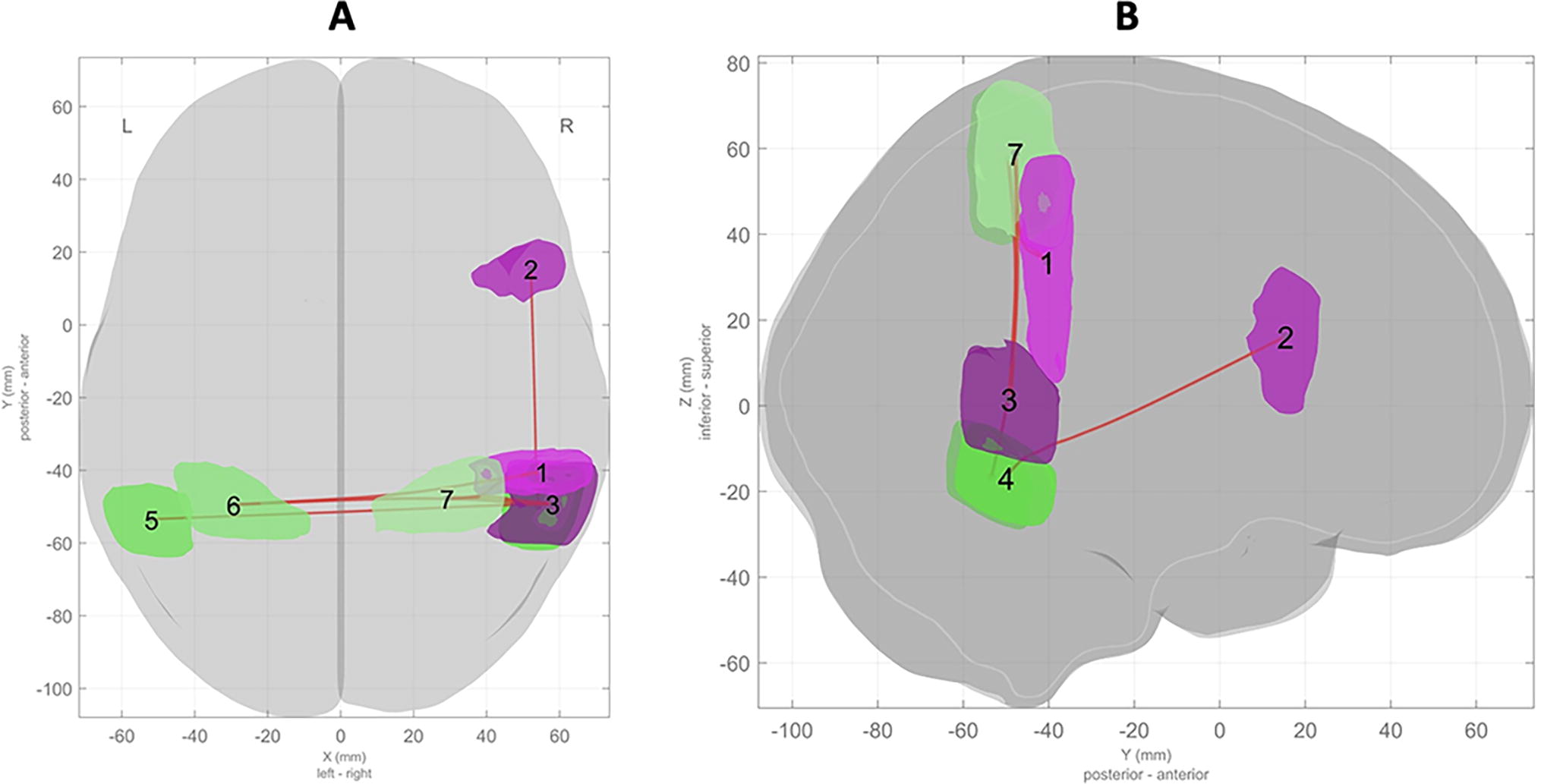
Association between rhyming ability and FC between brain areas supporting attention (DAN) and prosody (language). These are

Association between rhyming ability and FC between brain areas supporting imagery (DAN) and executive functions (F-P). Note: These are

Functional connectivity matrix showing higher FC between brain areas during the rhymed versus unrhymed story in children with higher rhyming abilities. Functional connectivity matrix showing higher FC between atlas-defined brain areas assigned to functional networks. Red indicates higher FC controlling for age, sex, and maternal education, with p-FDR < 0.05. Networks and areas assigned to them are (1) posterior DMN (pDMN; posterior cingulate gyrus, precuneus), (2) right fronto-parietal (FPN; frontal pole, middle frontal gyrus, superior frontal gyrus), (3) right-sided language/prosody (lang; posterior supramarginal gyrus, opercular inferior frontal gyrus, temporo-occipital aspect of middle temporal gyrus), (4) dorsal attention network (DAN; right temporo-occipital aspect of inferior temporal gyrus, left temporo-occipital aspect of inferior temporal gyrus, right superior parietal lobule, left superior parietal lobule). pDMN, posterior default mode network.
Discussion
Brain development is a dynamic process influenced by genetic and environmental factors (Gilmore et al., 2018; Knudsen, 2004). Essential structural and functional networks are established by age 2 and then shaped by experiences (Gilmore et al., 2018). Emergent literacy is a neurodevelopmental continuum involving skills that increase throughout life (“unconstrained,” e.g., vocabulary) and those that manifest during finite windows (“constrained,” e.g., phonological awareness). Currently, little is known about the neural basis of rhyming, a constrained skill that is among the first phonological abilities to emerge, typically during the preschool age range (3–5 years old). This reflects an important gap in knowledge, as low or absent rhyming ability in children has been shown to predict broader language and reading difficulties. Improved insights into the neurobiology of rhyming may also help better characterize the influence of genetic and environmental factors on this skill during early childhood, such as shared reading at home. The degree in which rhyming stories are processed differently in a child’s brain and have the potential to enhance nascent rhyming abilities has also not previously been studied. The purpose of the current study was to use MRI to examine relationships between early rhyming abilities and FC in atlas-defined brain areas and networks.
The finding of higher FC between brain areas supporting focused attention (in the DAN) and prosody (right-sided frontal-temporal areas) suggests that children with better rhyming abilities are more able to attend to the rhythmic/prosodic aspect of a rhymed narrative. This is consistent with our hypothesis and compelling, in that right-sided brain areas were exclusively involved, as described in neuroimaging literature where prosodic processing has been mapped to right frontal-temporal areas. This also aligns with extensive educational literature citing the appeal of rhymed narratives at this age in terms of child attention, interest, and comprehension (Johnson and Hayes, 1987; Király et al., 2017; Read et al., 2014; Read, 2014). While involving a rudimentary posttest, superior recall of the rhymed story presented during MRI, despite similar difficulty and interest levels, is also consistent with this prior evidence. It is reasonable to speculate that prosody in a rhymed story may fuel an “Attention Enhancer” involving right-sided frontal-temporal areas and the dorsal network, in turn enhancing processing of other aspects of the story (e.g., language, imagery). For younger children lacking rhyme awareness and those with low or absent rhyming abilities, attention to prosody (e.g., cadence, rhythm) may confer similar benefits—as seems to be the case in nursery rhymes and songs—though this warrants further study. A proposed model for an Attention Enhancer is provided in Figure 4.

Proposed “Attention Enhancer” model for rhymed story processing. Green arrows represent relationships involving a rhymed story and orange arrows those involving an unrhymed story, with thicker reflecting their magnitude. Here, the story narrative is proposed to engage the DAN, with enhanced functional connectivity involving right-lateralized language areas during the rhymed story (the attention “enhancer”), suggested to reflect greater prosodic processing. In turn, engagement and functional connectivity of networks supporting imagery (DMN) and executive functions (frontal-parietal network; F-P) is enhanced during the rhymed story.
The second major finding in this study involves higher FC between brain areas supporting executive functions (FPN) and higher-order visual imagery (DMN) during the rhymed relative to the unrhymed story. This suggests that children with better rhyming abilities more efficiently engage this neural infrastructure, possibly catalyzed by the Attention Enhancer described earlier. Specifically, the FPN supports working memory (Ferguson et al., 2021; Lara and Wallis, 2015), an executive function that is vital for semantic processing, including integration of visual imagery, which involves the DMN (Daselaar et al., 2010; Hutton et al., 2020c; Zvyagintsev et al., 2013). As a result, children with greater abilities may be better equipped to visualize and process rhymed stories, driving greater interest and comprehension relative to unrhymed stories. It is unclear whether these benefits manifest in the absence of an Attention Enhancer effect (i.e., whether they are dependent on attention to rhyme and/or prosody), such as in children lacking rhyme awareness. Although surprising and not hypothesized here, this finding is consistent with prior studies affirming the appeal of rhymed stories during early childhood in terms of interest and comprehension (Bryant et al., 1989; Johnson and Hayes, 1987; Király et al., 2017; Read et al., 2014) and also with the current finding of superior recall of the rhymed versus the unrhymed story presented during MRI. Although in need of further study, this finding suggests an additional biomarker of rhyme processing in young children, involving imagery and executive networks.
Although the current study findings are of interest in terms of potential neural mechanisms, they also provide insights into means to tailor and target shared reading guidance for families. For example, explicitly encouraging parents to emphasize letter-sound mappings during rhyming stories where attention is high may encourage nascent phonological and emergent literacy abilities by engaging language and executive brain networks. Developing materials for use in literacy programs for young children, especially from disadvantaged backgrounds, emphasizing such practices during shared reading may be useful in this regard. These could complement rhymed books that are widely distributed by early literacy programs such as Reach Out and Read (ROR) and Dolly Parton’s Imagination Library (Funge et al., 2017; Klass et al., 2009). The appeal of rhyming songs and stories during early childhood is also well documented across languages and cultures and encouraged in guidance to “Talk, sing, read every day!” (Office of Early Childhood Development, 2023). Findings in the current study are also reasonably aligned with other cited benefits of rhymed stories, including enhanced memorization and attention to story structure (supported by the frontal-parietal network) and phonemic awareness (prosody, language network) (Johnson and Hayes, 1987; Király et al., 2017; Read et al., 2014; Read, 2014). Rhyming stories also tend to promote bonding between children and grown-up story readers (i.e., it is a shared, fun experience), suggesting other potential neurobiological drivers of benefits (e.g., oxytocin release) (Bryant et al., 1989; Lang et al., 2020; Read, 2014; Scatliffe et al., 2019). Larger, longitudinal studies are needed to quantify the impact of guidance and interventions on rhyming, phonological, and other emergent literacy skills from early childhood through school age.
This study has limitations that should be noted. Although 17% of participants met poverty criteria, the sample was largely of higher income and maternal education, and results might be different with greater socioeconomic diversity. Analyses were limited to children completing fMRI and meeting necessary motion criteria, which may bias results toward those with higher self-regulation and other behavioral characteristics. The cross-sectional nature of the study prohibits comment on causality, which requires a longitudinal design. The posttest used was rudimentary in nature and may not fully capture story comprehension. Finally, the MRI analyses are unable to discern directionality of FC between these brain networks, rendering the attention enhancer model (Fig. 4) speculative at this stage.
This study also has important strengths. It involves a reasonably large sample of young children who are challenging to scan, where there have been few MRI-based studies involving rhyming/phonological skills. It involved a continuous story listening design to compare FC between entire narratives, akin to how children listen to actual stories (i.e., the fMRI task is ecological). Analyses considered demographic covariates age, sex, and maternal education, minimizing the potentially confounding influence of general maturation and home environment (Frangou et al., 2021; Girault et al., 2020; Wu et al., 2013). Most notably, significant results were found controlling for maternal education, which has been cited as a major SES-related predictor of child cognitive development (Jackson et al., 2017). MRI analyses applied conservative FDR correction, reducing the likelihood of false-positive results. Perhaps most important, the current findings align with both MRI and education-based literature describing neural signatures of prosody and other skills noted and cognitive and relational benefits of rhymed stories during early childhood. Altogether, while several findings are unclear and/or speculative, attributable to the complex nature of early brain development, this study suggests compelling, potential neural biomarkers of nascent rhyming abilities in preschool-age children. Although preliminary, it is an important step toward greater understanding of how underlying neural architecture supporting these skills is formed, the basis of deficits, and how these may be most effectively addressed via existing programs such as ROR and early interventions.
Conclusions
This fMRI-based study identified associations between higher rhyming skills and FC between atlas-defined brain areas in preschool-age children. Children with higher abilities had greater FC between areas supporting prosodic processing and focused attention (proposed as an “Attention Enhancer”) and also between those supporting visual imagery and executive functions. Comprehension of the rhymed story was also higher. These findings are consistent with prior neuroimaging literature in terms of brain areas involved and also with educational research in terms of the appeal of rhymed stories at this age. Further studies are needed to determine directionality of these relationships and also the influence of genetics and environmental factors.
Footnotes
Acknowledgments
The authors would like to thank Amy Kerr for her diligence in collecting these data and the CCHMC Research Foundation for their support of early-career investigators and this work. They also thank Dr. Scott Holland for his support and mentorship.
Authors’ Contributions
J.S.H. conceptualized the study; secured funding; oversaw all aspects including administration, design of the MRI protocol, data curation, and analyses; drafted the initial article and subsequent revisions; and approved the final article as submitted. J.D. collaborated in and oversaw the MRI acquisition protocol, conducted all MRI data analyses, created all derivative tables and figures, assisted with article preparation and revisions, and approved the final article as submitted. T.D. provided guidance on conceptualization of the study, its funding mechanism, and administration; collaborated in interpretation of data analyses; reviewed and revised the article; and approved the final article as submitted. T.H.-K. collaborated in the conceptualization of the study, design, and implementation of the MRI protocol; provided guidance on data analyses and interpretation; reviewed and revised the article and subsequent revisions; and approved the final article as submitted.
Data Availability Statement
All survey and MRI data for this study were newly acquired via methods described. These data will be made available to the scientific community in a deidentified manner upon notice of publication via written request to the corresponding author (J.S.H.). Requests must include description of the project (e.g., project outline) and also acknowledgment of the data source in any grant submissions, presentations, or publications. The rationale for written request is that no repository currently exists, and creation would exceed the scope and current funding resources of the study team. Any costs associated with data transfer will be the responsibility of the requesting parties. Software used in the current analyses is freely available and described in the Methods section.
Authors Disclosure Statement
The authors have no financial relationships relevant to this article to disclose.
Funding Information
This study was funded by a Procter Scholar Award from the Cincinnati Children’s Research Foundation (J.S.H.).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.