
Abstract
Introduction:
The influence of the cerebellum in poststroke aphasia recovery is poorly understood. Despite the right cerebellum being identified as a critical region involved in both language and cognitive functions, little is known about functional connections between the cerebellum and bilateral cortical hemispheres following stroke. This study investigated the relationship between chronic poststroke naming deficits and cerebello-cerebral resting-state functional connectivity (FC).
Methods:
Twenty-five cognitively normal participants and 42 participants with chronic poststroke aphasia underwent resting-state functional magnetic resonance imaging. Participants with aphasia also underwent language assessment. We conducted regions of interest (ROI)-to-ROI analyses to investigate the FC between the right cerebellar Crus I/II (seed ROI; Cereb1r/Cereb2r) and bilateral cortical language regions and compared these results to cognitively normal controls. Single-subject connectivity parameters were extracted and used as independent variables in a stepwise multiple linear regression model associating Boston Naming Test (BNT) score with FC measures.
Results:
FC analyses demonstrated correlations between the right cerebellar Crus I/II and both left and right cortical regions for both cognitively normal controls and stroke participants. Additionally, aphasia severity and lesion load had an effect on the cerebello-cerebral network connectivity in participants with aphasia. In a stepwise multiple linear regression, controlling for aphasia severity, time poststroke and lesion load, FC between the right Cereb2–left Cereb1 (standardized beta [std B]= −0.255, p < 0.004), right Cereb2–right anterior MTG (std B = 0.259, p < 0.004), and the right Cereb2–left anterior STG (std B = −0.208, p < 0.018) were significant predictors of BNT score. The overall model fit was R 2 = 0.786 (p = 0.001).
Conclusion:
Functional connections between the right cerebellum and residual bilateral cerebral hemisphere regions may play a role in predicting naming ability in poststroke aphasia.
Impact Statement
The right cerebellum is a critical region involved in both language and cognitive functions. However, the influence of the cerebellum in poststroke aphasia recovery is poorly understood. Functional connectivity (FC) analyses in our study demonstrated positive correlations between the right cerebellar Crus I/II and bilateral cortical language regions in chronic poststroke individuals. Further, FC between the right cerebellum and residual regions in bilateral cerebral hemispheres were significant predictors of Boston Naming Test score, which suggest the potential role in predicting naming ability in poststroke aphasia.
Introduction
An estimated one-third of all stroke survivors experience aphasia (Pedersen et al., 2004), a disorder of language, which can lead to significant social isolation (Nicholas et al., 2020), reduced return to work (Matérne et al., 2019), and depression (Kauhanen et al., 2000). This, in turn, may severely impact functional outcomes and lead to reduced quality of life (Bullier et al., 2020; Sheppard and Sebastian, 2021). Despite the goal of developing novel and influential treatments for participants with poststroke aphasia, there are still many unanswered questions related to the underlying pathophysiology of poststroke aphasia.
Processes contributing to both aphasic language generation and recovery may differ depending on time poststroke, in the acute, subacute, or chronic time frames (Hillis et al., 2000; Sebastian et al., 2016; Stockert et al., 2020). For example, recovery in the acute and subacute periods may be due to the resolution of diaschisis (resolution of temporary neuronal dysfunction in regions connected to the lesion), or reperfusion of the ischemic penumbra (cortical regions at risk of infarction but salvageable with restoration of blood flow) (Carrera and Tononi, 2014; Hillis et al., 2006; Kiran, 2012; Wawrzyniak et al., 2022). These processes are not observed in the chronic phase. Furthermore, focal structural damage that occurs in the acute stroke setting may also lead to language dysfunction in regions of the brain spatially distant to the lesion site but functionally connected through integrated, large-scale networks (Boes et al., 2015; Engel et al., 2001; Klingbeil et al., 2019; Siegel et al., 2018). With improved understanding of the structural and physiological etiology of poststroke aphasia, recovery mechanisms, and timing of recovery, better-informed treatment approaches may be utilized.
Resting-state functional magnetic resonance imaging (rsfMRI) is an emerging method to understand network pathology in poststroke aphasia (Klingbeil et al., 2019). Measuring the temporal coherence of slow intrinsic blood oxygen level dependent (BOLD)-signal fluctuations during rest allows the evaluation of regional interactions, termed resting-state functional connectivity (FC), between gray matter areas within the brain (Fox and Raichle, 2007). rsfMRI is particularly useful for studying aphasia, as the results are not confounded by task demands. Previous rsfMRI investigations have shown that FC is disrupted across a distributed network of language and cognitive regions following a stroke, and various factors influence the network connectivity including language severity, lesion size, and time poststroke (Hagoort, 2014; Harvey et al., 2013; Nair et al., 2015; New et al., 2015; Ramage et al., 2020; Sandberg, 2017; Sebastian et al., 2016; Siegel et al., 2016; Yang et al., 2017; Zhao et al., 2018; Zhu et al., 2014). Studies have also shown that the integrity of resting-state functional networks may predict aphasia treatment response following stroke (Falconer et al., 2024; Iorga et al., 2021).
For example, Sandberg (2017) showed that, compared with healthy age-matched adults, participants with chronic poststroke aphasia had reduced FC in several resting-state networks including the semantic network, and connectivity for resting-state networks increased with decreasing aphasia severity and decreased with increasing lesion size. In another study, participants with subacute poststroke aphasia had lower FC involving the left fronto-parietal network (LFPN) than healthy controls, and stroke participants with lower connectivity had more severe comprehension deficits (Zhu et al., 2014). A follow-up 2 months poststroke showed that improvements in comprehension were associated with increased LFPN FC, reflecting correspondence between a shift toward more normal FC and the restoration of more typical language functions. In contrast, using voxel-mirrored-homotopic connectivity in participants with acute lacunar stroke, Yang et al. found increased inter-hemispheric resting-state FC in the inferior frontal gyrus (IFG), anterior superior temporal gyrus, and lingual gyrus compared with controls (Yang et al., 2017). They also found participants with worse language performance exhibited increased inter-hemispheric homotopic connectivity in the superior temporal gyrus.
A large-scale study by Siegel et al. (2016) examined FC in subacute stroke participants across multiple resting-state networks. They found that subacute stroke participants had reduced FC between homologous regions in the ipsilesional and contralesional hemispheres relative to healthy controls. Additionally, FC was as effective as lesion location at predicting severity of language impairment, whereas memory deficits were better predicted by FC than by lesion location. In another study, Falconer et al. showed that mean FC of four predefined brain networks (language, default mode, dorsal attention, and salience networks) were independent predictors of response to language therapy in participants with poststroke aphasia (Falconer et al., 2024). Although many studies have utilized rsfMRI to explore cortical networks, there are few that examined the role of the right cerebellum in poststroke aphasia.
Seminal anatomical and behavioral studies have shown that the right cerebellum is involved in a range of cognitive processes, including language (Leiner et al., 1991; Petersen et al., 1990; Raichle et al., 1994; Schmahmann and Sherman, 1997); see (Price, 2012) for review. Prior studies have demonstrated the right cerebellar lobules VI and VII (including Crus I/II) are engaged during a variety of language and cognitive processes (Mariën et al., 2014; Stoodley and Schmahmann, 2010). Functional neuroimaging has demonstrated cerebellar activation during tasks involving word generation and both semantic and phonemic fluency (Baillieux et al., 2009). The cerebellum is also involved in motor speech planning (Riecker et al., 2000), verbal working memory (Boehringer et al., 2013; Pope and Miall, 2012; Stoodley et al., 2012), syntax processing (Stoodley et al., 2012), and reading and writing (Mariën et al., 2014).
Cerebello-cortical connections exist, with both feedforward and feedback projections, including the dentate-rubro-thalamo-cortical tract and the cortico-ponto-cerebellar tracts, with cortical regions including the frontal, parietal, temporal, and occipital lobes (Karavasilis et al., 2019; Keser et al., 2015). More specifically, the right cerebellum connects with regions that are active during syntactic and semantic processes, including the supramarginal gyrus (SMG), medial superior frontal gyrus, left inferior frontal gyrus (MTG), and the precuneus (Binder et al., 2009; Callan et al., 2003; Price, 2012). Hubrich-Ungureanu et al. demonstrated that in healthy volunteers, right-handed subjects with left hemisphere dominance demonstrated fMRI activation patterns involving the fronto-parietal cortex and the contralateral right cerebellar hemisphere while performing a silent verbal fluency task (Hubrich-Ungureanu et al., 2002). In another study, utilizing positron emission tomography imaging, healthy subjects demonstrated activation of the right lateral cerebellum during tasks of word generation (Papathanassiou et al., 2000). Finally, a large metanalysis of cerebellar contributions to language and cognition found activation peaks during expressive language tasks involving the cerebellum (Keren‐Happuch et al., 2014). The extent to which these connections are altered following cortical stroke has yet to be thoroughly explored.
Neuromodulation of the right posterolateral cerebellum with transcranial direct current stimulation (tDCS) in cognitively normal controls impacts the FC between the cerebellum and cortical language networks (D’Mello et al., 2017; Turkeltaub et al., 2016). In addition, treatment of poststroke aphasia with right cerebellar neuromodulation has demonstrated improvement in language skills in the chronic phase (Sebastian et al., 2020), further suggesting contribution of the cerebellum to aphasia recovery. The right cerebellum could potentially be an optimal site for tDCS treatment in poststroke aphasia because it often remains structurally intact in individuals with poststroke aphasia. Furthermore, cerebellar tDCS could be a valuable tool in handling heterogeneous participants with varying lesion size and site in the left hemisphere. However, the influence of cerebello-cerebral resting-state FC in the chronic phase of aphasia recovery is poorly understood (Van Dun and Mariën, 2015). Understanding the functional connections between the right cerebellum and left hemisphere language regions may help us determine who is likely to respond more favorably to cerebellar brain stimulation treatment.
This study investigated the relationship between cerebello-cerebral resting-state FC and poststroke naming deficits. A focus was placed on naming because difficulty with naming is the most common deficit in poststroke aphasia. Given that the right posterolateral cerebellum lobule VII (including Crus I/II) is an important region for language processing, it may contribute to naming recovery after stroke. We hypothesized that increased resting-state FC (positive correlations) between the right posterolateral cerebellum and the left hemisphere language regions would be associated with better naming performance. In addition, we also investigated the effect of aphasia severity, lesion size, and time poststroke on FC. We hypothesized that higher aphasia severity and lesion load would be associated with reduced cerebello-cerebral network connectivity.
Materials and Methods
Study procedure and participants
Forty-two, right-handed stroke participants (mean age 58.6 years age range 29–89 years; 13 female) with chronic (>6 months poststroke) aphasia due to left hemisphere stroke were included in this study (see Table 1 for demographic details). This was a retrospective study of prospectively collected data on individuals who previously experienced a stroke in the left hemisphere. Data were obtained from a database of individuals who participated in aphasia treatment and/or longitudinal stroke recovery studies at the Johns Hopkins University School of Medicine from 2013 to 2022. Inclusion criteria for this analysis were as follows: left hemisphere stroke, right-handed, at least 6 months poststroke, and fluent speakers of English. Participants with multiple strokes (regardless of stroke type) were admitted into this study. Participants were excluded if they had history of non-stroke related speech/language impairment or other neurological deficits affecting the brain, no research MRI scans available, and lesion in the right cerebellum.
Stroke Demographics
BDAE, Boston Diagnostic Aphasia Exam; BNT, Boston Naming Test.
We also included 25 cognitively normal right-handed participants (mean age 63 years, age range 49–78 years; 11 females) from the Biomarker for Older Controls at Risk for Dementia (BIOCARD) project who had MRI scans available. The BIOCARD project is an ongoing longitudinal prospective cohort study designed to identify variables among cognitively normal individuals that predict subsequent development of mild to moderate symptoms of Alzheimer’s and dementia. Please see Albert et al. (2014) and Oishi et al. (2022) for further details. This study included MRI data collected between January 2015 and October 2019 and from 25 participants who were cognitively normal (please see Supplementary Table S1 for normal participants demographics).
All study participants provided written and informed consent, and the study was approved by The Johns Hopkins University Institutional Review Board.
Language testing
All participants with stroke underwent language testing including the short version of the Boston Naming Test (BNT) (Mack et al., 1992) and the short version of the Boston Diagnostic Aphasia Examination (BDAE) (Goodglass et al., 2001). The latter was administered to calculate the aphasia severity scores.
Neuroimaging
Participants were scanned at the F.M. Kirby Research Center for Functional Brain Imaging at the Kennedy Krieger Institute on a 3T Philips MRI magnet with a 32-channel head coil. For the stroke patients, rsfMRI was collected using an echo-planar imaging (EPI) sequence with the following parameters: number of slices = 35; field of view (FOV) = 240 × 240 mm2; voxel size = 3 × 3 × 3 mm3; time repetition (TR)/TE = 2000/30 ms; flip angle = 90°. The duration of each scan session was 420 s and comprised 210 functional volumes. Magnetization-prepared rapid gradient echo (MPRAGE) scans were also using following parameters: TR = 6.8 ms, TE = 3.1 ms, shot interval 3000 ms, flip angle = 8°, FOV = 240 × 256 mm2, 170 sagittal slices covering the whole brain, voxel size= 1 × 1 × 1.2 mm3, and scan duration = 5 min 59 sec. T2-weighted spin-echo images were acquired axially with a slice thickness of 2 mm with no gaps, FOV = 212 × 212, TR/TE:4171/12 ms. FLAIR images were acquired axially with 5 mm slice thickness and FOV:512/512, TR/TE:8000/140 ms.
For the control participants, rsfMRI was collected using an EPI sequence with the following parameters: number of slices = 48; FOV = 212 × 212 mm2; voxel size = 3.3 × 3.3 × 3.3 mm3; TR/TE = 3000/30 ms; flip angle = 75°. The duration of each scan session was 420 s and comprised 140 functional volumes. MPRAGE scans were also obtained using the following parameters: TR = 6.8 ms, TE = 3.1 ms, shot interval 3000 ms, flip angle = 8°, FOV = 240 × 256 mm2, 170 sagittal slices covering the whole brain, voxel size =1 × 1 × 1.2 mm3, and scan duration = 5 min 59 sec).
Lesion tracing and overlay map generation
Lesion tracings were quantified manually through the use of MRIcron software (nitrc.org/projects/mricron) by a stroke neurologist experienced in lesion segmentation using high-resolution T2-weighted, FLAIR, and T1 weighted images (Keser et al., 2021). Lesion load included bilateral white matter hyperintensities (WMH) and infarct volumes, as previous studies have shown that WMH, even in the right hemisphere may be associated with language functions poststroke (Basilakos et al., 2019; Wright et al., 2018). For the normalization of the volumes of WM hyperintensities and infarcts, the total intracranial volume (ICV) was estimated by summing all segmented white matter, gray matter, and cerebrospinal fluid (CSF) from each subject by using T1-weighted maps. The lesion load was calculated as the percentage of total lesion volume to ICV (lesion load = lesion volume/ICV × 100). Lesions on T1 weighted images were converted to a common space using SPM Clinical Toolbox (Rorden et al., 2012). A lesion overlay map was calculated from all lesions and superimposed on a template brain using MRIcron (Rorden et al., 2007) (Fig. 1).
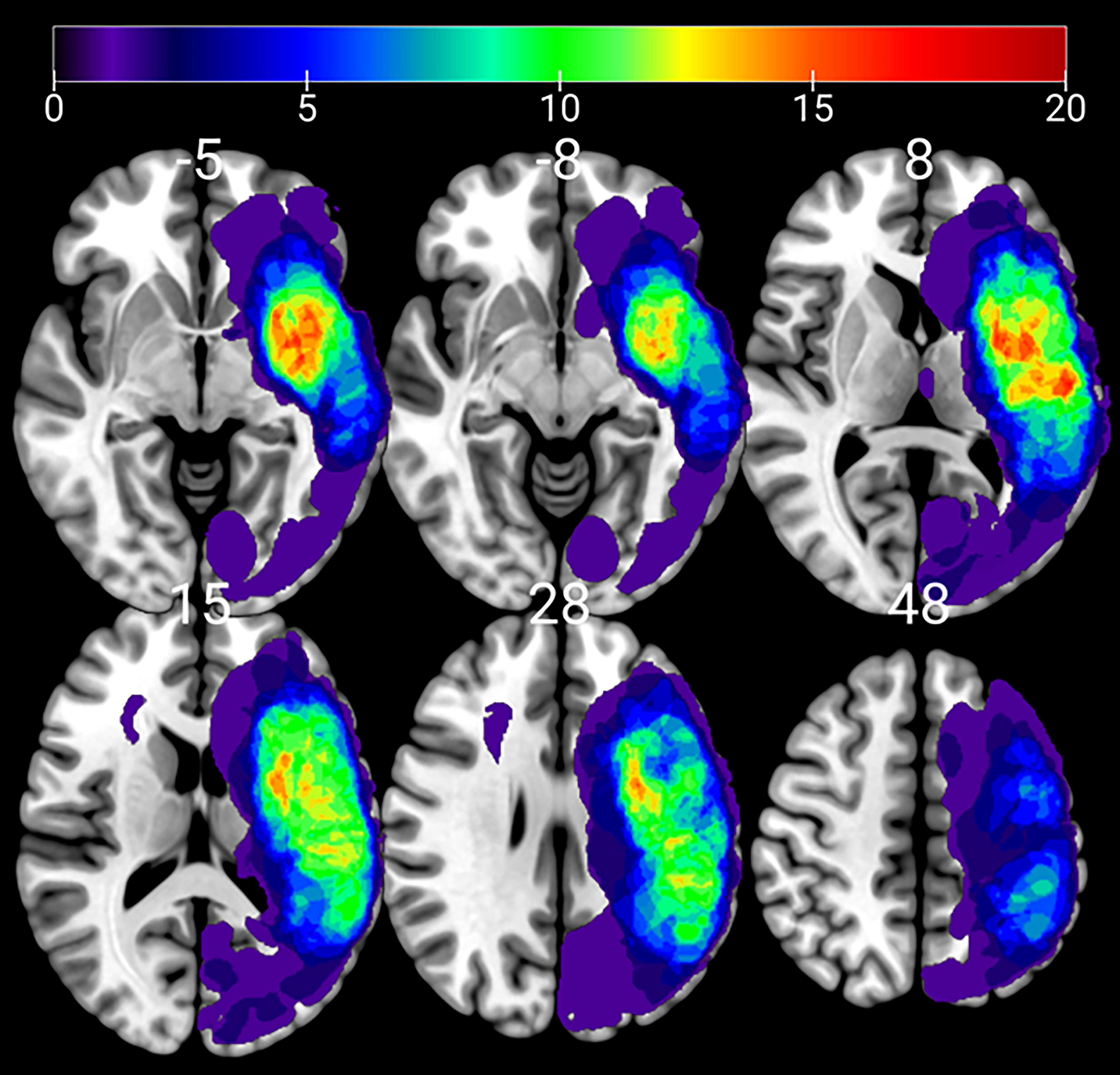
Lesion overlay map of the individual lesions of all 42 patients. Maps were overlaid on a MNI152 T1-template using MRIcroGL. Color scale indicates the number of patients with a lesion in a particular voxel.
Resting-state fMRI pre-processing and data analysis
Results included in this article come from analyses performed using CONN (Whitfield-Gabrieli and Nieto-Castanon, 2012) (RRID:SCR_009550) release 22.a (Whitfield-Gabrieli and Nieto-Castanon, 2012) and SPM (Penny et al., 2011)(RRID:SCR_007037) release 12.7771 implemented in MATLAB (version R2023b) (Nieto-Castanon, 2020). We performed separate analyses for stroke participants and healthy controls.
Preprocessing
Functional and anatomical data were preprocessed using a flexible preprocessing pipeline (Nieto-Castanon, 2020) including realignment with correction of susceptibility distortion interactions, slice timing correction, outlier detection, direct segmentation and MNI-space normalization, and smoothing. Functional data were realigned using SPM realign and unwarp procedure (Anderson et al., 2001), where all scans were coregistered to a reference image (first scan of the first session) using a least squares approach and a six parameter (rigid body) transformation (Friston et al., 1995), and resampled using b-spline interpolation to correct for motion and magnetic susceptibility interactions. Temporal misalignment between different slices of the functional data (acquired in ascending order) was corrected following SPM slice-timing correction procedure (Henson et al., 1995; Sladky et al., 2011), using sinc temporal interpolation to resample each slice BOLD timeseries to a common mid-acquisition time. Potential outlier scans were identified using artifact detection tools (ART) (Whitfield-Gabrieli et al., 2011) as acquisitions with framewise displacement above 0.9 mm or global BOLD signal changes above 5 standard deviation (Nieto-Castanon, 2022; Power et al., 2014), and a reference BOLD image was computed for each subject by averaging all scans excluding outliers. Functional and anatomical data were normalized into standard MNI space, segmented into gray matter, white matter, and CSF tissue classes, and resampled to 2 mm isotropic voxels following a direct normalization procedure (Calhoun et al., 2017; Nieto-Castanon, 2022) using SPM unified segmentation and normalization algorithm (Ashburner and Friston, 2005; Ashburner, 2007) with the default IXI-549 tissue probability map template. Lastly, functional data were smoothed using spatial convolution with a Gaussian kernel of 8 mm full width half maximum.
It should be noted that after the segmentation step, all scans were visually checked by the first author to verify that the segmentation was done correctly. In addition, stroke participants scans were also checked to ensure the lesion was segmented into the CSF mask. The lesion is classified as a noise source, so it does not contribute noisy data to the effect of interest. Stroke participants scans were also visually checked after the normalization step to rule out any skewing effects of the lesion (Nieto-Castanon, 2020).
Denoising
Functional data were denoised using a standard denoising pipeline (Nieto-Castanon, 2020) including the regression of potential confounding effects characterized by white matter timeseries (5 CompCor noise components), CSF timeseries (5 CompCor noise components), motion parameters and their first order derivatives (12 factors) (Friston et al., 1996), outlier scans (below 38 factors) (Power et al., 2014), session effects and their first order derivatives (2 factors), and linear trends (2 factors) within each functional run, followed by bandpass frequency filtering of the BOLD timeseries (Hallquist et al., 2013) between 0.008 Hz and 0.09 Hz. CompCor (Behzadi et al., 2007; Chai et al., 2012) noise components within white matter and CSF were estimated by computing the average BOLD signal as well as the largest principal components orthogonal to the BOLD average, motion parameters, and outlier scans within each subject’s eroded segmentation masks. From the number of noise terms included in this denoising strategy, the effective degrees of freedom of the BOLD signal after denoising were estimated to range from 52.2 to 56.1 (average 55.8) across all control participants and 47.9–60.4 (average 58.7) across all stroke participants (Nieto-Castanon, 2022).
Selection of regions of interest
Resting-state FC analysis included interhemispheric, intrahemispheric, and cerebello-cerebral connectivity of language regions of interest (ROI). We conducted ROI-to-ROI analyses with a priori ROIs as the seed regions. The ROIs consisted of regions that comprise the language networks including regions in the left hemisphere, right hemisphere, and cerebellum. Language ROIs were defined based on prior language meta-analyses and reviews in neurologically cognitively normal controls and participants with aphasia (Binder et al., 2009; Keren-Happuch et al., 2014; Price, 2012; Stoodley and Schmahmann, 2009; Vigneau et al., 2006; Vigneau et al., 2011). The ROIs included the following regions: right cerebellar Crus I/Crus II (Cereb1 and Cereb2, language seed ROIs) (Stoodley and Schmahmann, 2009); left Cereb1 and Cereb2; bilateral cerebellar lobules seven (Cereb7) and eight (Cereb8); middle frontal gyrus (MidFG); IFG pars opercularis (IFG oper) and pars triangularis (IFG tri); anterior superior temporal gyrus (aSTG), posterior superior temporal gyrus (pSTG); anterior MTG (aMTG), posterior MTG (pMTG); anterior inferior temporal gyrus (aITG), posterior inferior temporal gyrus (pITG); angular gyrus (AG); anterior SMG (aSMG), and posterior SMG (pSMG). Please see Supplementary Figure S1, which illustrates the location of the ROIs.
First-level analysis
ROI-to-ROI connectivity matrices were estimated characterizing the patterns of FC with 32 ROIs from the Harvard–Oxford atlas (Desikan et al., 2006) and the Automated Anatomical Labeling (AAL) atlas (Rolls et al., 2015). FC strength was represented by Fisher-transformed bivariate correlation coefficients from a weighted general linear model (weighted-GLM [Nieto-Castanon, 2020]), defined separately for each pair of seed and target areas, modeling the association between their BOLD signal timeseries. In order to compensate for possible transient magnetization effects at the beginning of each run, individual scans were weighted by a step function convolved with an SPM canonical hemodynamic response function and rectified.
Second-level analyses
Second-level analyses were performed using a General Linear Model (GLM) (Nieto-Castanon, 2020). For each individual voxel, a separate GLM was estimated, with first-level connectivity measures at this voxel as dependent variables and other subject-level identifiers as independent variables. We included three covariates for the second level analysis for stroke participants with aphasia: lesion load, aphasia severity, and time poststroke. This was done to determine whether and how these factors play a role in cerebro-cerebellar network connectivity. Voxel-level hypotheses were evaluated using multivariate parametric statistics with random-effects across subjects and sample covariance estimation across multiple measurements. Inferences were performed at the level of individual clusters (groups of contiguous voxels). Cluster-level inferences were based on nonparametric statistics from randomization/permutation analyses (Bullmore et al., 1999; Nieto-Castanon, 2020), with 1000 residual-randomization iterations. Results were thresholded using a combination of a cluster-forming p < 0.05 voxel-level threshold, and a familywise false discovery rate (FDR) corrected p-FDR <0.05 cluster-mass threshold (Chumbley et al., 2010).
Additional analysis
To assess the relationship between naming performance and cerebello-cerebral network connectivity, we first extracted the single-subject FC parameters from CONN for each ROI that showed significant connectivity with the seed ROI (right Cereb1 and right Cereb 2) for stroke participants with aphasia. We then performed a stepwise multiple linear regression on the individual connectivity strengths and BNT scores. Covariates included aphasia severity (BDAE aphasia severity score), lesion load, and time poststroke. p-Values <0.05 were considered statistically significant. Analyses were conducted using SPSS version 27. This analysis was not performed in CONN Toolbox due to an incomplete model and instead investigated outside of CONN.
Results
Lesion overlay
Individual lesions for all 42 participants seen on MRI were overlaid on a Montreal Neurological Institute 152 (MNI152) template using MRIcroGL (Fig. 1). Lesion density was highest in the left frontotemporal region (middle cerebral artery territory).
FC analysis
Results of the ROI-to-ROI FC analysis can be seen in Figures 2 and 3 for both the cognitively normal controls and stroke participants. Please see Supplementary Table S2 and Supplementary Table S3 for stroke participants and normal controls connection strength and p values.
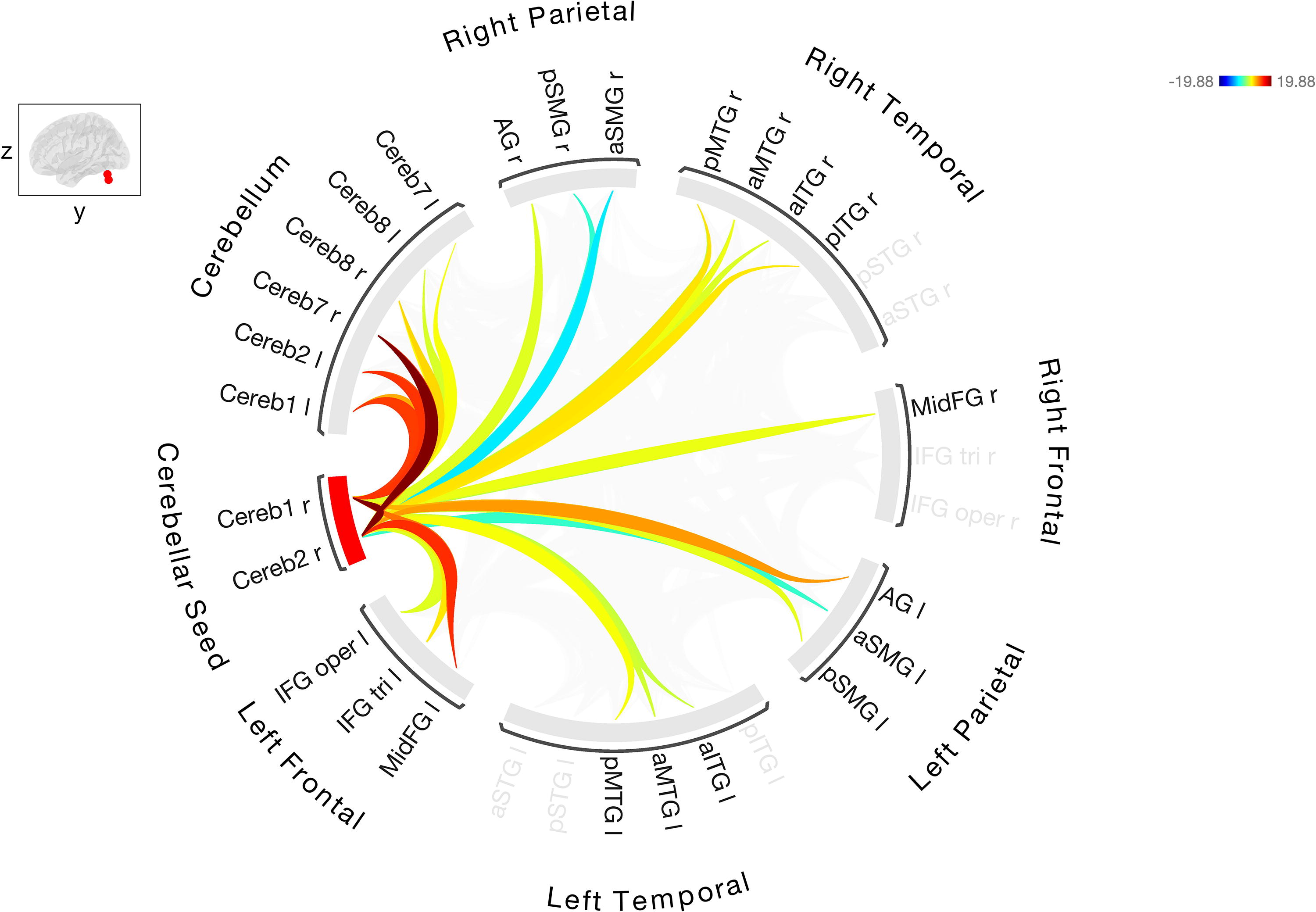
Resting-state FC between the right Cerebellar Crus I/II, seed ROI with the other cerebellar and cortical ROIs in healthy controls. Warmer colors indicate increased degree of connectivity (positive correlations) and cooler colors indicate decreased connectivity (negative correlations). FC, functional connectivity; ROI, regions of interest.
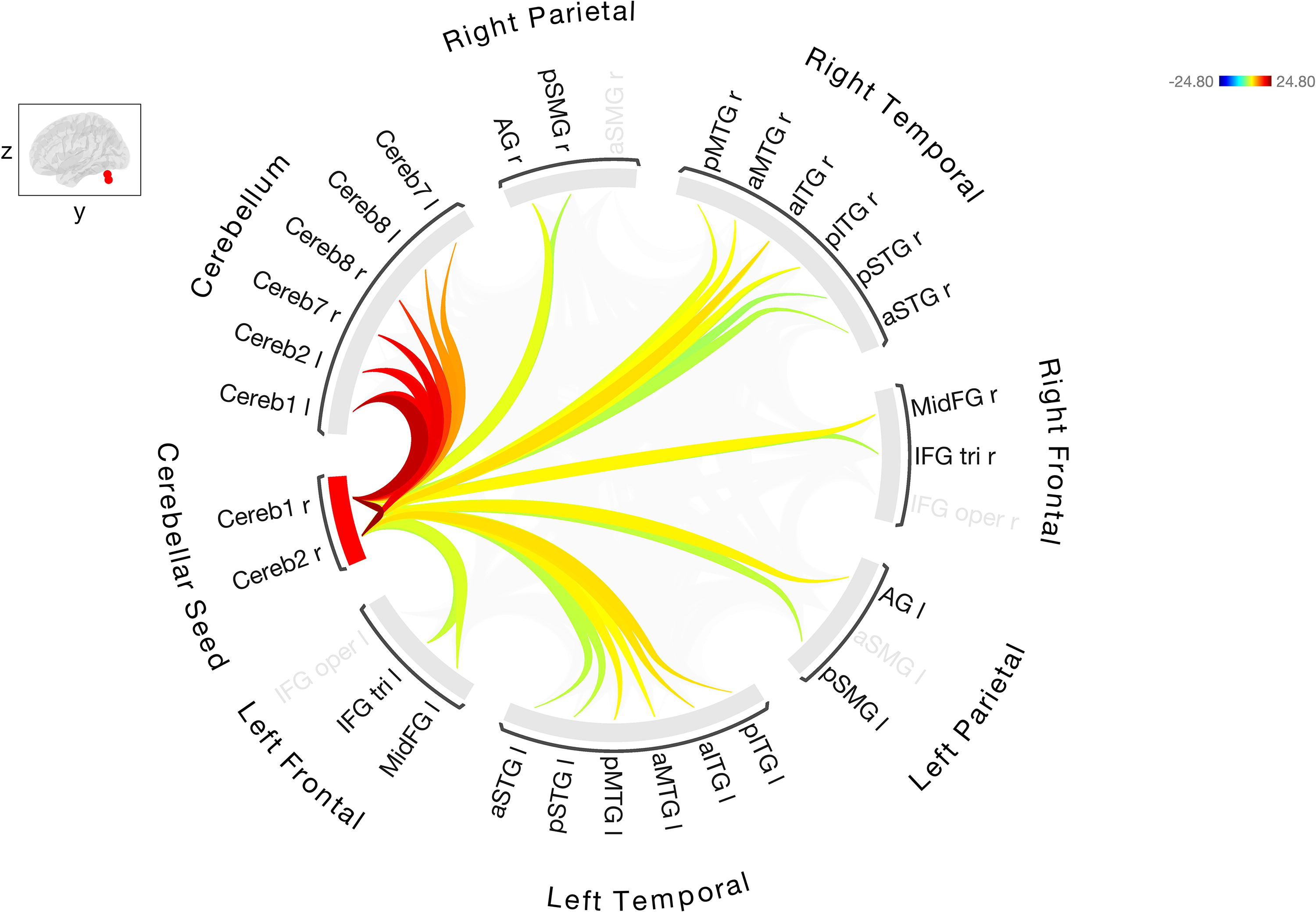
Resting-state FC between the right Cerebellar Crus I/II, seed ROI with the other cerebellar and cortical ROIs in stroke participants. Warmer colors indicate increased degree of connectivity (positive correlations) and cooler colors indicate decreased connectivity (negative correlations).
FC analyses in cognitively normal controls demonstrated FC between the cerebellar seed region Crus I/II(Cereb1 and Cereb2) and left and right hemisphere language ROIs in the fronto-temporo-parietal regions. The strongest cerebello-cerebral connections were between seed regions and left MidFG (right Cereb2 to left MidFG [T (24) = 13.15, p < 0.0001]; right Cereb1 to left MidFG [T (24) = 10.56, p < 0.0001]). Controls showed positive correlations (stronger connectivity) between Cereb1 and 2 and left frontal (IFGoper, IFGtri, MidFG), left parietal (aSMG, pSMG AG), left temporal (aITG, aMTG, pMTG), right frontal (MidFG), right temporal (aITG, pITG, aMTG, pMTG), and bilateral Crus 7/8. Controls showed negative correlations (inhibitory connectivity) between Crus I/II and left aSMG, and right aSMG and pSMG.
FC analysis in the participants with stroke showed positive correlations between the right cerebellar Crus I/II (Cereb1 and Cereb2) and bilateral frontal, temporal, and parietal regions. The strongest cerebello-cerebral connections were between right Cereb 2 and bilateral ITG (Cereb 2 to left pITG [T (41) = 7.75, p < 0.0001] and Cereb2 to right aITG [T (41) = 7.69, p < 0.0001]). Other significant positive correlations to the left hemisphere included the left frontal (IFG tri and MidFG), left temporal (aSTG, pSTG, aMTG, pMTG, aITG, pITG), and left parietal (pSMG and AG). Positive correlations between the right cerebellar seed regions and right cortical regions were also observed, including frontal (IFG tri and MidFG), temporal (aSTG, pSTG, aMTG, pMTG, aITG, pITG), and parietal (pSMG and AG). Positive correlations were also observed between seed cerebellar regions and other bilateral cerebellar language regions. A significant positive correlation between seed region right Cereb1 and left STG was absent in cognitively normal controls. Negative correlation that existed in the cognitively normal controls in the left STG was positive in stroke participants.
Effect of aphasia severity and lesion load
Additional analyses were performed to determine the effect of total lesion load, aphasia severity, and time poststroke on FC within the group of stroke participants with aphasia. The BDAE aphasia severity score was used as a measure of aphasia severity. Controlling for lesion load altered the FC as well as the strength of the connections (Fig. 4 and Supplementary Table S4). The positive correlations that were observed between right Cereb1 and Cereb2 and left frontal and parietal regions were no longer significant after controlling for lesion load. Significant FC between cerebellar seed region and bilateral anterior/posterior ITG and negative connectivity between seed region and left AG was observed. Controlling for aphasia severity also altered the connectivity and there were only two connections that were significant between right cerebellar seed regions and bilateral language ROIs: left aSMG and right aSTG (Fig. 5 and Supplementary Table S5). It should be noted that stroke participants were quite heterogenous in lesion load and BDAE aphasia severity. Many stroke participants had large lesions involving the fronto-temporo-parietal regions. Although the relationship between aphasia severity and lesion load was negative (r = −0.47, p = 0.02), not all patients with the largest lesion had the lowest BDAE score. Adding the covariate of time poststroke did not change the FC results (Supplementary Fig. S2).
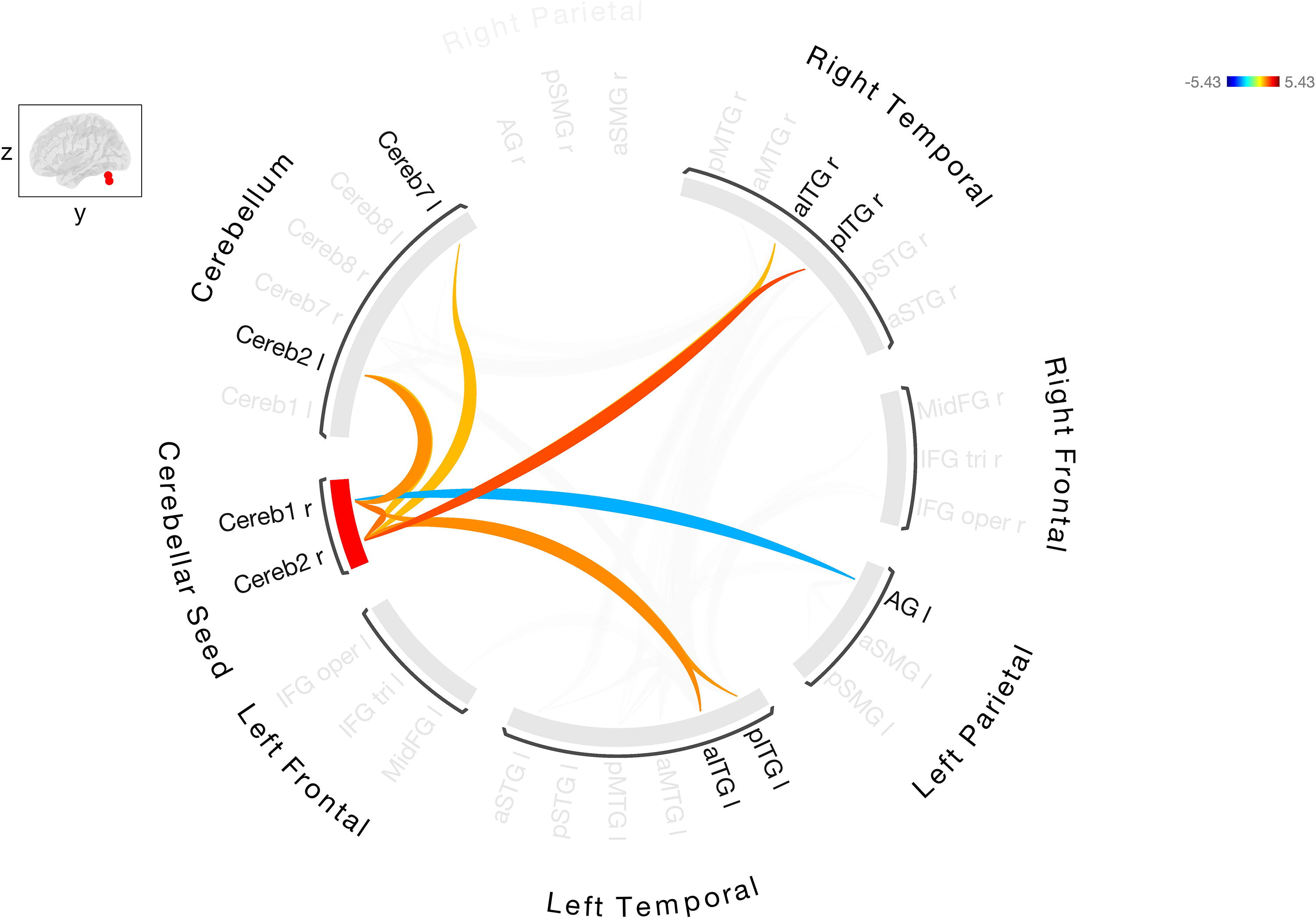
Resting-state FC between the right Cerebellar Crus I/II, seed ROI with the other cerebellar and cortical ROIs in stroke participants after controlling for lesion load. Warmer colors indicate increased degree of connectivity (positive correlations) and cooler colors indicate decreased connectivity (negative correlations).
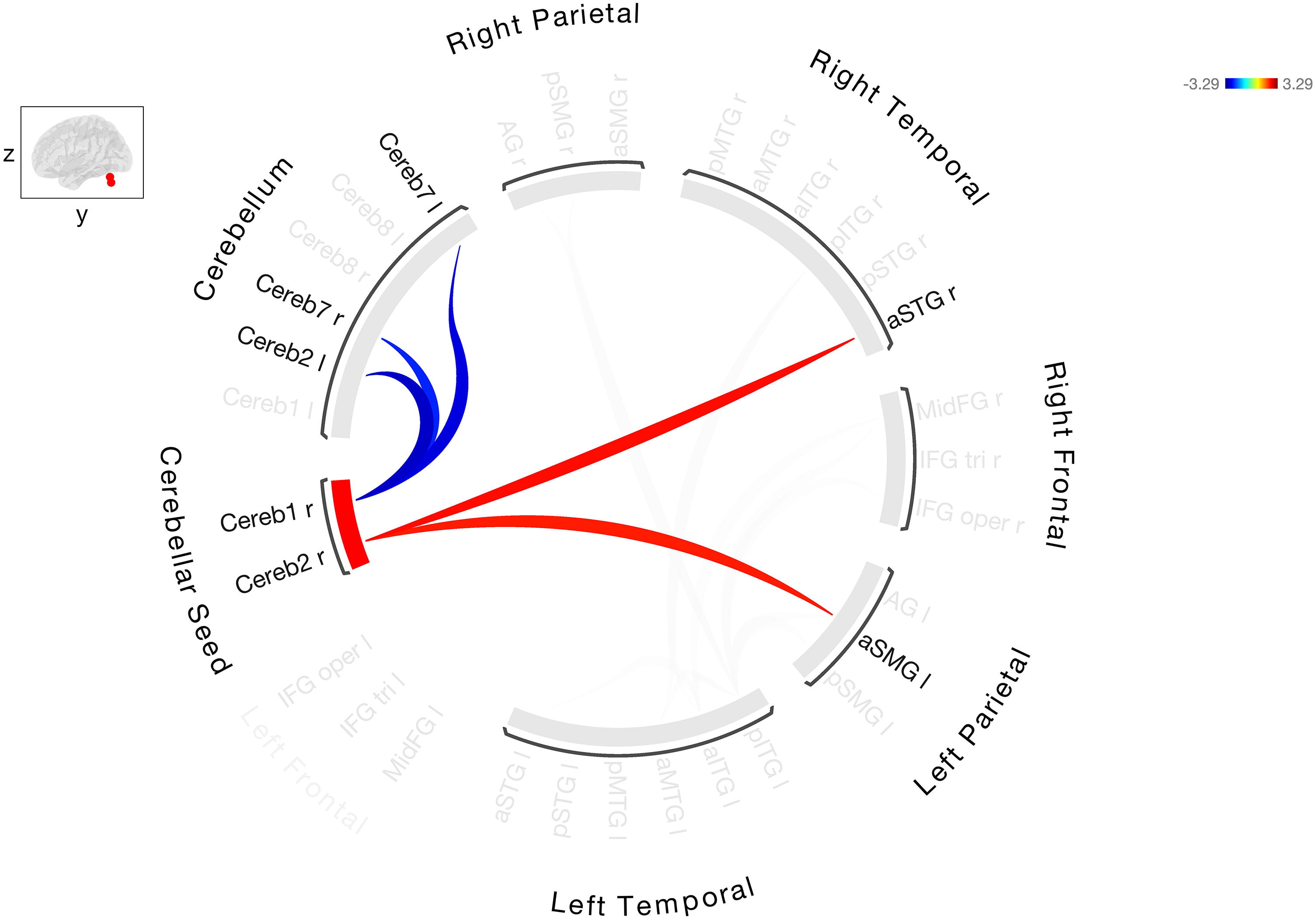
Resting-state FC between right Cerebellar Crus I/II, seed ROI with the other cerebellar and cortical ROIs in stroke participants after controlling aphasia severity. Warmer colors indicate increased degree of connectivity (positive correlations) and cooler colors indicate decreased connectivity (negative correlations).
FC correlation with naming
In a stepwise multiple linear regression, with time poststroke, lesion load, and aphasia severity specified as control variables/covariates, FC between the right Cereb2–left Cereb1 (standardized beta [std B] = −0.255, p < 0.004), the right Cereb2–right aMTG (std B = 0.259, p < 0.004) and the right Cereb2–left aSTG (std B = −0.208, p < 0.018) were significant predictors of BNT score. The overall model fit was R 2 = 0.786 (p = 0.001, significant after Bonferroni correction). This indicates that the effect of specific cerebello-cerebral FC metrics on naming are above and beyond the effect of aphasia severity, lesion load, and time poststroke.
Discussion
To our knowledge, this is the first study to investigate the FC between the right posterolateral cerebellum and bilateral cortical language regions in stroke participants with chronic aphasia. Our results showed that both the right cerebellar Crus I and II (Cereb1, Cereb2) demonstrate resting-state FC to contralateral (left) and ipsilateral (right) cortical regions involved in language processing and that integrity of these connections are predictors of naming ability in the chronic phase following stroke. The FC pattern observed in cognitively normal controls is consistent with previous studies and shows a robust connectivity between the cerebellar right Crus I/II seed region and fronto-parietal language-cognitive networks (Buckner et al., 2011; D’Mello et al., 2017; Gao et al., 2017; Hubrich-Ungureanu et al., 2002; Turkeltaub et al., 2016). This further supports the observation that a major portion of the human cerebellum is functionally coupled to cerebral association areas (Habas et al., 2009; Krienen and Buckner, 2009; O’Reilly et al., 2010).
In our study, the FC patterns observed in the participants with stroke were similar to that observed in the cognitively normal controls. However, it should be noted that although patterns were similar, the strength of connections may differ as the two groups were not directly compared due to differences in imaging acquisition parameters. There were some key differences between the cognitively normal controls and those with stroke. For example, the cognitively normal controls demonstrated strong FC between the right cerebellar seed regions (Crus I/II) and left IFG, pars opercularis. This connection was absent in participants with stroke. On the other hand, participants with stroke showed strong positive FC between the right cerebellar seed regions and left STG, whereas cognitively normal controls showed no FC between right cerebellar seed regions and left STG. In addition, participants with stroke showed additional FC between right cerebellar seed regions and right language regions that were absent in cognitively normal controls (Supplementary Tables S2 and S3). We speculate that this could be in part due to lesion load and cortical reorganization of residual left hemisphere regions. Studies have shown that when participants have lesions affecting critical left hemisphere language regions, they utilize residual tissue within the left hemisphere and also recruit right hemisphere language regions for language processing (Griffis et al., 2017; LaCroix et al., 2021; Postman-Caucheteux et al., 2010; Sebastian and Kiran, 2011; Skipper-Kallal et al., 2017; Turkeltaub et al., 2011; Wilson and Schneck, 2021).
We found that adding the covariates of aphasia severity and lesion load changed the cerebello-cerebral network connectivity in stroke participants, but not time poststroke. When lesion load was added as a covariate, the only significant cerebello-cerebral connection was between right Cereb1 and 2 and bilateral inferior temporal gyrus. This could be because the left inferior temporal gyrus was not lesioned in most stroke participants. However, there were other regions not lesioned in most participants (left middle frontal gyrus) that showed no significant connection with the right cerebellar language regions. This indicates that the regions not affected by the lesion can still have altered FC patterns. It should be noted that the lesion load in our study also included bilateral WMH. The WMH in our stroke cohort was overall symmetric and involved both anterior and posterior regions. Thus, it is unclear if the WMH may likely be contributing to the observed effect. Adding the covariate of aphasia severity also resulted in only two significant connections between the right cerebellar seed region and bilateral cortical language ROIs (left anterior SMG and right anterior superior temporal gyrus). Similar to lesion load, aphasia severity led to extensive reduction in the FC in the cerebello-cerebral network. The results suggest that higher aphasia severity might result in widespread dysfunction in the core language network resulting in reduced FC. Many of our stroke participants who had severe aphasia also had large lesions, which may have contributed to overall dampened FC. Our findings are in line with other studies that have shown that aphasia severity and lesion volume affect resting-state networks in stroke participants with aphasia (Falconer et al, 2024; Meier et al., 2023; Siegel et al., 2016; Sandberg, 2017).
We also found that decreased FC between the right cerebellar seed regions and the left cortical language regions was associated with poorer naming performance. The left superior temporal gyrus and right MTG and their connections to cerebellar Crus II were predictive of BNT score. Interestingly, even after controlling for aphasia severity, left hemisphere damage (lesion load) and time poststroke, significant predictors of naming score included bihemispheric cerebello-cortical connections. These findings are consistent with other studies indicating that the left STG and the right MTG are critical to naming and naming recovery (Baldo et al., 2013; Hillis et al., 2018; Hope et al., 2017; Xing et al., 2016). These findings also align with previous reports (including ours) that stronger FC in cortical networks corresponds to better naming performance. For example, we examined longitudinal resting-state FC changes associated with naming function in individuals with naming deficits after left posterior cerebral artery stroke, who had preserved frontal, temporal, and parietal language areas (Sebastian et al., 2016). We found that ipsilesional and contralesional cortical FC including connections between left STG and right MTG improved from acute to chronic phase in the following weeks only in participants who showed good naming recovery. Ramage and colleagues (Ramage et al., 2020) showed that stronger FC between the left IFG–left MFG and the right MTG–right IFG predicted higher naming scores in patients with chronic aphasia.
Our findings support the conclusion that resting-state FC is often disrupted following stroke in both lesioned and nonlesioned areas of the brain and the integrity of various resting-state networks like domain general, sensorimotor, visuospatial, and language is associated with stroke outcomes (Carter et al., 2012; He et al., 2007; Machner et al., 2020). Possible mechanisms contributing to this dampened FC include retrograde cerebellar changes in structure, metabolism, and neuronal activity observed to occur following cortical stroke (Förster et al., 2014) and crossed cerebellar diaschisis. This later phenomenon is often reported after cortical stroke, which results in disruption of communication and functional inactivation, hypoperfusion, and hypometabolism of the contralateral cerebellum (Lin et al., 2009). Data from our laboratory offer support for the theory that cortical strokes can trigger secondary degeneration of cerebellar regions. In a recent study (Keser et al., 2023), we examined the microstructural integrity of the cerebellum and the cortico-cerebellar connections and their role in picture naming in chronic stroke participants with aphasia. Results showed that a significant mean diffusivity increase for the right cerebellar segments and the right dentato-rubro-thalamo-cortical tract, and the integrity of these tracts was associated with picture naming. We speculate that this could reflect the progressive structural alterations resulting from Wallerian degeneration in chronic stroke (Schulz et al., 2017).
In summary, our results suggest that the cerebellar-cortical connection is an important player in naming performance following cortical stroke. These findings are in line with findings from other FC studies showing that the cerebellum is an important node within the distributed network for stroke recovery (Park et al., 2011; Rosso et al., 2013; Várkuti et al., 2013; Wang et al., 2010). In particular, changes in FC between the ipsilesional motor cortex and the contralesional cerebellum correlated with individual upper-extremity function improvement (Várkuti et al., 2013). Our findings also support the assertion that the reorganization of the spared brain regions beyond the left hemisphere is crucial for poststroke language recovery. It has been demonstrated that for left hemispheric strokes, regions of the intact right hemisphere may compensate for lost or impaired contralateral functioning (Balaev et al., 2016). For example, Hope and colleagues report that right-hemisphere regions undergo systematic structural adaptations to support language functions, even in the chronic phase of recovery, and that the degree of these changes relates to word retrieval (Hope et al., 2017). Thus, the right cerebellum may be an important region for poststroke language recovery. Our study indicates that activation of the cerebellum may be a potential alternative therapeutic target for neuromodulation for individuals with chronic stroke and decreased FC between the right cerebellum and left cortex. There is already promising evidence that cerebellar neuromodulation can drive improvements in language skills of those with aphasia (Marangolo et al., 2018; Sebastian et al., 2020).
Our study has several limitations. First, our sample size is small, and this limits the generalizability of our results; new and larger samples are necessary to confirm our results. Second, our study design is cross-sectional; therefore, we are unable to investigate the longitudinal resting-state connectivity between the cerebellum and cortex, which, in turn, would help to better understand the physiology of language recovery poststroke. Third, we do not have acute perfusion/cerebral blood flow imaging data in all of our participants, which meant we were unable to rule out the influence of crossed-cerebellar diaschisis. Fourth, we found aphasia severity had an effect on cerebello-cerebral language network connectivity. Studies have shown that participants with aphasia exhibit disrupted FC not only in language-specific networks but also in domain-general networks, including the dorsal attention, salience, and default mode networks. However, it is unclear if the reduction in FC was observed in other networks as we only included language-specific networks. In addition, we assessed aphasia severity using the BDAE aphasia severity rating scale. This is a 5-point rating scale compared with the 100-point scale of the Aphasia-Quotient on the Western Aphasia Battery Revised (WAB-R, Kertesz, 2007 ). It is unclear if using the WAB-R could have resulted in differences when controlling for aphasia severity. Finally, we were unable to directly compare stroke participants with control participants due to differences in temporal resolution (TR) and signal-to-noise ratio between scans. Data interpolation was not possible and as a result, differences in TR would influence statistical results and connectivity measures.
Conclusions
In summary, these results suggest that cerebello-cerebral resting-state FC may be an important component of poststroke naming recovery. This investigation proposes that functional connections between the right cerebellum and both residual left and right hemisphere regions may also contribute to naming ability in poststroke aphasia. Opportunities to enhance cerebello-cortical connections exist and may be a new target for aphasia rehabilitation strategies poststroke.
Footnotes
Authors’ Contributions
Conceptualization (R.S., A.E.H.); Data curation (J.H.K., R.S., A.E.H.); Formal analysis (J.S., A.S., Z.K.); Funding acquisition (R.S., A.E.H.); Investigation (J.S., R.S.); Methodology (J.S., R.S.); Project administration (R.S., J.H.K., S.C., D.C.T.); Resources (R.S.); Software (J.S., R.S., Z.K.); Supervision (R.S., A.E.H.); Validation (J.S., R.S.); Visualization (J.S., R.S.); Roles/writing—original draft (J.S., R.S.); Writing—review and editing (J.S., J.H.K., S.C., Z.K., D.C.T., A.E.H., R.S.).
Author Disclosure Statement
J.S.—full-time salaried faculty member, Weill Cornell Medicine. Site PI for industry-sponsored clinical trials: (1) Merz Pharmaceuticals. Prospective, randomized, double-blind, placebo-controlled, multicenter study to investigate the efficacy and safety of NT 201 in the treatment of lower limb spasticity caused by stroke or traumatic brain injury in adult subjects, followed by an open label extension with or without combined upper limb treatment. (2) Ipsen Pharmaceuticals. A multicenter, interventional, post-marketing, randomized, double-blind, crossover study to evaluate the clinical safety and efficacy of AbobotulinumtoxinA (Dysport®) in comparison with OnabotulinumtoxinA (Botox®) when treating adults with upper limb spasticity. J.H.K.—no disclosures. S.C.—no disclosures. Z.K.—full-time salaried faculty member, Mayo Clinic. Editorial member for Elsevier’s Practice Update. D.C.T.—full-time salaried faculty member, Johns Hopkins University; grant support NIH National Institute on Aging (NIA) R01AG075111, NIH National Institute on Deafness and Other Communication Disorders (NIDCD)/NIA R01 DC011317; Review Editor, Frontiers in Neurology, Applied Neuroimaging; Associate Editor, Frontiers in Neurology, Stroke; member of the Collaboration of Aphasia Trialists, International Stroke Rehabilitation and Recovery Alliance, and Primary Progressive Aphasia Task Force of the National Aphasia Association. A.E.H.—full-time salaried faculty member, Johns Hopkins University; grant support R01 DC 05375, P50 NIDCD P50 DC014664, R01 DC015466, R25NS065729; Editor-in-Chief of Stroke; Associate Editor, Practice Update Neurology. R.S.—full-time salaried faculty member, Johns Hopkins University; grant support NIH NIDCD R01 DC019639. Review Editor for Frontiers in Neurology.
Funding Information
This study was made possible by support from National Institutes of Health (NIH; NIDCD) through K99/R00DC015554, R01DC05375, P50DC014664, and R56/R01DC019639. The MRI equipment in this study was funded by NIH grant 1S10OD021648. The BIOCARD Study (officially entitled “Biomarkers of Cognitive Decline Among Normal Individuals: the BIOCARD cohort”) is supported by a grant from the NIA (U19-AG033655).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Data S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.