
Abstract
Background:
Olfactory deterioration is suggested to be a predictor of some neurodegenerative diseases. Recent studies indicate that physical exercise has a positive relationship with olfactory performance, and a subregion in the prefrontal cortex (PFC) may play an important role in olfactory processing. The PFC is not only related to olfactory function but it also engages in complex functions such as cognition and emotional processing.
Methodology:
Our study compared the functional connectivity between the olfactory cortex and the PFC in healthy individuals who exercised regularly and healthy persons who did not. Those who exercised more than three times/week for at least 30 min each time were considered the exercise group, and those who did not meet this exercise criteria were considered the nonexercise group. We also assessed their odor threshold. Participants were aged 55 years or older, and the two groups were balanced for age, sex, body mass index, and educational level.
Results:
We found that compared with individuals who did not exercise, exercisers had a significantly lower threshold for detecting odors. In addition, the olfactory cortex had stronger connectivity with the PFC in exercisers than in nonexercisers. More specifically, when the PFC was grouped into three subregions, namely, the ventrolateral prefrontal cortex (VLPFC), dorsolateral prefrontal cortex (DLPFC), and frontopolar cortex (FPA), Pearson correlation analysis revealed stronger connectivity between the VLPFC and the orbitofrontal cortex (OFC), between the OFC and the FPA, and between the left and right OFC hemispheres in the exercisers. In addition, Granger causality indicated higher directional connectivity from the DLPFC to the OFC in exercisers than in nonexercisers.
Conclusion:
Our findings indicated that the exercise group not only had better olfactory performance but also had stronger functional connectivity between the olfactory cortex and the PFC than nonexercise group.
Impact Statement
The present research found that individuals in the exercise group had significantly better olfactory thresholds than those in the nonexercise group. In addition, Granger causality indicated higher directional connectivity from the DLPFC to the OFC in exercisers than in nonexercisers. These findings suggest that exercise is associated not only with better olfaction but also with strengthened functional connectivity between the olfactory cortex and the PFC. We believe that this finding will be of great interest to the readers of Brain Connectivity.
Introduction
Evidence for human olfactory function deteriorating with age is robust. One study indicated that the prevalence rate of olfactory dysfunction increases monotonically with each additional decade of age until reaching 39.4% for participants with 80 or more years of age (Hoffman et al., 2016). Numerous studies have shown that people with neurodegenerative diseases (e.g., Alzheimer’s disease or Parkinson’s disease) have impaired olfactory function (Foguem et al., 2018; Naudin et al., 2014; Olofsson et al., 2016). Considering the close correlation between the dysfunction of the olfactory system and neurodegenerative diseases, researchers have suggested that olfactory dysfunction may be a predictor of some neurodegenerative diseases. The Quality Standards Subcommittee of the American Academy of Neurology recommended olfaction deterioration as one of the diagnostic criteria for Parkinson’s disease (Suchowersky et al., 2006). A similar recommendation has been suggested for olfactory impairment as a potential predictor of Alzheimer’s disease (Foster et al., 2008). Among the various measurable olfaction domains, the olfactory threshold, that is, the lowest concentration an odorant can be reliably detected, is considered to have the most solid relationship with the severity of Alzheimer’s disease: the degree of olfactory threshold damage may reflect the severity of Alzheimer’s disease (Murphy et al., 1990). Owing to the importance of olfaction, how to prevent it from declining has engaged many researchers. Recent studies have found that olfactory training improves olfactory performance (Sorokowska et al., 2017) and alters brain structures associated with the performance (Al Aïn et al., 2019). In addition to olfactory training, physical exercise also demonstrates a positive relationship with olfactory performance. A few studies have used psychologist tests to investigate the role of exercise in improving olfactory function or to compare the differences in olfactory performance between exercise and nonexercise cohorts (Manestaret al., 2013; Rosenfeldt et al., 2016; Zhang et al., 2020). Physical exercise not only improves olfactory acuity (Cameron et al., 2016) but also olfactory identification (Rosenfeldt et al., 2016). Many previous studies have suggested that physical exercise is highly related to olfaction (Rosenfeldt et al., 2016; Schubert et al., 2013; Sollai and Crnjar, 2021; Zhang et al., 2020). Plausible mechanisms by which physical exercise may impact olfaction include that exercise increases the levels of nerve growth factor and synapsin I in the olfactory bulb (Chae et al., 2014), enhances the size of olfaction-related brain regions (Köbe et al., 2016), such as the hippocampus, and alters the activation pattern of the brain (Ji et al., 2017). However, to the best of our knowledge, no study has explored the functional connectivity between the olfactory cortex and other brain regions using any brain imaging technique.
The orbitofrontal cortex (OFC) is considered a critical part of the olfactory cortex because odor stimulus information flows from the nasal cavity to the olfactory bulb, then passes through the primary olfactory cortex (and areas such as the amygdala), and finally into the secondary olfactory cortex (which includes the OFC) (Mai and Paxinos, 2011). Several previous studies using neuroimaging techniques have examined the OFC to study how the olfactory system in the brain functions when an odor stimulus is presented (Cecchetto et al., 2019; Frie et al., 2018; Ishimaru et al., 2004; Kokan et al., 2011; Wu et al., 2012). The close association between PFC and executive function was robust (Colom et al., 2013; Gunning-Dixon and Raz, 2003; Raz et al., 1998). Its subregions such as the ventrolateral prefrontal cortex (VLPFC), dorsolateral prefrontal cortex (DLPFC), and frontopolar cortex (FPA) were found to correlate with different cognition functions. VLPFC plays an important role in psychopathology and social cognition (Andreou et al., 2018; Derntl et al., 2012; Lee et al., 2010; Moseley et al., 2013), DLPFC was critical in cognitive control and emotional regulation (Bonelli & Cummings, 2007; Braver et al., 1997; Rugg & Wilding, 2000), and the FPA has a vital function to support complicated cognitive processes, such as multitasking, high-level goal maintenance, and social cognition (Burgess et al., 2007).
The main aim of the present study was to use a noninvasive optical imaging method called functional near-infrared spectroscopy (fNIRS) to explore whether the functional connectivity between the olfactory cortex (OFC) and other brain regions (VLPFC/DLPFC/FPA) is higher among individuals who exercise compared with that among individuals who do not perform regular exercise.
Resting-state brain functional activity refers to the internal state of the brain in a natural state without external stimulation. The spontaneous fluctuations of the activity of the resting brain are the default state of the brain (Biswal et al., 1995). It has been suggested that spontaneous fluctuations in the resting state may be related to task-provoked brain activation (Bruns et al., 2000; Leopold et al., 2003). In the absence of any task or stimulus, the resting brain intrinsically reflects task-related brain activation (Fox et al., 2005). Moreover, other studies have revealed that resting-state networks are highly associated with higher-level cognition. Increased local connectivity is associated with better cognitive performance (Douw et al., 2011) and with higher intelligence (Wang et al., 2011). It has also been suggested that individuals with higher executive function may have greater connectivity of resting-state networks than those with lower executive function (Reineberg et al., 2015). In recent years, the resting-state functional connectivity in the olfactory networks and other brain regions were well documented (Karunanayaka et al., 2017; Lu et al., 2019). Therefore, we posited that default brain functional connectivity in the resting state may reveal neural mechanisms of cognitive and olfactory function, providing fundamental insights into brain activation in the absence of interference.
fNIRS monitors brain activity by measuring the concentration changes in oxygenated hemoglobin (HbO2) and deoxygenated hemoglobin (Hb) with high temporal resolution (on a time scale of ms). Compared with other brain imaging techniques, fNIRS has many advantages for measuring resting-state functional connectivity. First, fNIRS more accurately than electroencephalography or magnetoencephalography determines the location of assessed brain regions because hemoglobin concentrations measured by fNIRS represent brain activity located only directly below the probes; by contrast, localization is more difficult using either electroencephalography or magnetoencephalography owing to the volume conduction effect of these techniques (Srinivasan et al., 2007). Second, compared with functional magnetic resonance imaging, fNIRS is more portable, less expensive, and more robust against movement artifacts. These strengths enable fNIRS to detect resting-state brain functional activity in humans.
In summary, the purpose of the present research was to investigate the functional connectivity between the OFC and other brain regions (VLPFC/DLPFC/FPA) by using fNIRS to examine the brain during the resting state of individuals who regularly exercise and those who do not. The main goal was to provide fundamental insights into how exercise alters olfaction and the connections between the olfactory cortex and other brain areas.
Methods
Participants
The ability of humans to identify odors starts to deteriorate in the fifth decade of life (Doty et al., 1984; Zhang & Wang, 2017). Thus, to study participants across a range of olfactory abilities, only individuals with a potential risk of olfaction impairment, that is, those of 55 years of age or older, but who were otherwise apparently healthy were included in the present study. To achieve healthy cohorts, the inclusion criteria included individuals who had not received a diagnosis of a local respiratory tract disorder (e.g., nasal surgery, active rhinitis, or sinusitis), psychiatric disorder (e.g., depression), or neurologic disease (e.g., brain tumor, Parkinson’s disease). All participants reported their history of alcohol use and cigarette smoking. Because no sex difference exists in olfactory function among older participants (Wang et al., 2019), we did not counterbalance for sex during the selection process.
Participants were grouped by their self-reported exercise habits. It has been recommended that most adults should engage in 30–60min/day (150 min/week) of purposeful moderate exercise (Garber et al., 2011). Thus, participants who exercised more than three times/week for at least 30 min each time were considered the exercise group, and those who did not meet this exercise criteria were considered the nonexercise group. The average exercise times in the previous year for the two groups were significantly different, with the average amount of time spent exercising per week for the exercise group (240.50 ± 80.01 min/week) significantly higher than for the nonexercise group (19.50 ± 22.42 min/week) (p < 0.001). According to the results of a study by Schubert et al. (2013), more time spent in exercise per week is associated with a lower risk of developing an olfactory impairment.
The study protocol was performed in accordance with the Declaration of Helsinki and was approved by the Shanghai University of Sport Ethics Committee (Shanghai, China). All participants provided written informed consent that was obtained consistent with the Declaration of Helsinki guidelines prior to participation in this study.
Procedure
The entire procedure comprised a participant personal information questionnaire, a cognitive test, an olfactory threshold test, and a 10-min resting-state fNIRS. The time for each participant to complete all tests was ∼1 h.
Participant personal information questionnaire
Participant sex, age, height, weight, cigarette smoking and alcohol use history, educational level, surgical procedure history, drug use, regular physical exercise habits, and years engaged in regular physical exercise were collected using a self-reported questionnaire. For details about the questionnaire, see the Supplementary Material online.
Cognitive test
We used the Chinese version of the Mini-Mental State Examination (MMSE) (Folstein et al., 1975) to assess cognitive function. The MMSE is a standardized tool commonly and easily used to diagnose cognitive impairment. Scores on the MMSE range from 0 to 30, with lower scores indicating more severe cognitive impairment.
Odor threshold test
We examined participants’ olfactory threshold with the Sniffin’ Sticks Test using n-butanol as the stimulus odorant. Odorants were presented in commercially available (unfilled) felt-tip pens. The pens had a length of −14 cm, with an inner diameter of 1.3 cm. Instead of liquid dye, the tampon was filled with liquid odorants or odorants dissolved in propylene glycol, to a total volume of 4 mL. For odor presentation, the cap was removed by the experimenter for 3 sec and the pen’s tip was placed 2 cm in front of both nostrils. The test was conducted in a well-ventilated room. For procedural details, please see Hummel et al. (1997). The scores on the olfactory threshold test range from 1 to 16, with a lower score indicating reduced sensitivity, meaning an individual has a higher threshold and can detect only stronger concentrations.
Resting-state fNIRS examination
Due to the reproducibility of eyes-closed was slightly higher than that of eyes-open in resting-state (Duan et al., 2020), the participants were asked to sit quietly in a dark room with their eyes closed for 10 min. They were instructed to remain relaxed and to think as little as possible. When the participants finished the fNIRS examination, we would ask whether they keep awake during the examination and those who did not confirm their alterness were excluded in the final analysis.
A multichannel, continuous wave, fNIRS instrument (NIRScout, NIRx Medical Technologies LLC, Minneapolis, MN, USA) was used to monitor hemodynamic activity during this resting state. Data were collected at two wavelengths (780 and 830 nm) of near-infrared light with a sampling rate of 7.81 Hz. Eight sources and seven detectors of the optical probes comprised a 20-channel configuration and covered both the prefrontal cortex (PFC) and the OFC (Fig. 1). The probes were assigned according to the 10–20 electroencephalogram system with some adjustment to ensure that each emitter was 3 cm from its corresponding detector. The fNIRS measuring cap was placed on the participants’ head according to the manual: the optode holder corresponding to the EEG electrode position Cz (position green 14 on the NIRScap) should be positioned centrally both in the left-right direction (between the left and right preaurical points) as well as in the anterior posterior direction (between the nasion and inion). The ears should be pulled out of the slits completely to allow the cap to fit snugly onto the skin behind the ears. We associated the fNIRS channel locations with the corresponding brain regions based on previous studies (Okamoto et al., 2004, 2009; Tsuzuki et al., 2007). To further analyze the functional connectivity between different brain areas, the 20 channels were grouped into the following four brain areas based on the study by Jurcak et al. (2007): VLPFC (channels 1, 3, 4, 18, 19, and 20); DLPFC (channels 2, 5, 7, 8, 9, 10, 14, 15, and 17); FPA (channels 6, 12, and 16); and OFC (channels 11 and 13) (Fig. 1).
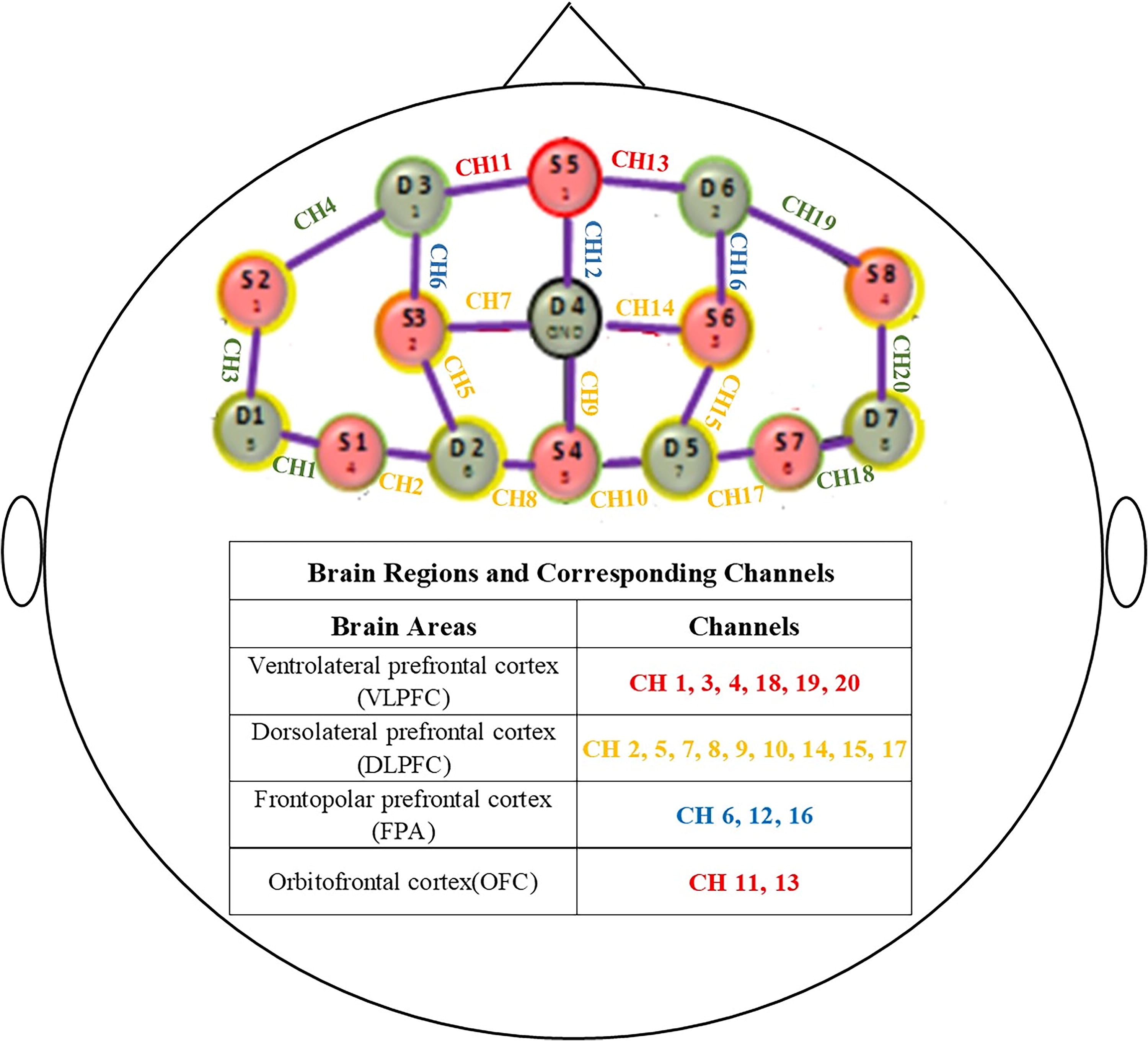
Schematic representation of the placement of the channels used to collect data during fNIRS and the corresponding brain regions. The regions of interest are highlighted as follows: red indicates the OFC; green, the VLPFC; yellow, the DLPFC; and blue, the FPA. CH, channel; DLPFC, dorsolateral prefrontal cortex; fNIRS, functional near-infrared spectroscopy; FPA, frontopolar cortex; OFC, orbitofrontal cortex; VLPFC, ventrolateral prefrontal cortex.
Statistical Analysis
Behavior
We used SPSS, version 22.0 (SPSS Inc.; Chicago, Il, USA) to analyze the behavior data. For quantitative variables (e.g., age, MMSE score, and odor threshold score), we used independent-samples t-tests to compare the difference between the exercise and nonexercise groups. For qualitative (categorical) variables (e.g., male or female percentage), we used the chi-squared test to evaluate the differences between the two groups. Pearson correlation was employed to analyze the relationship between different factors. All data are reported as means ± standard deviations (SDs). A two-sided p < 0.05 was considered statistically significant.
Hemodynamic imaging
Because HbO2 signals have a better signal-to-noise ratio than deoxygenated hemoglobin signals (Niu et al., 2013; Schaeffer et al., 2014), only HbO2 concentration ([HbO2]) data were analyzed using the HOMER2 (MGH-Martinos Center for Biomedical Imaging; Boston, MA) (Huppert et al., 2009) script for MATLAB (MathWorks; Natick, MA).
The data were first processed using the modified Beer-Lambert law (Cope et al., 1988), and then motion artifacts were removed in NIRlab. A bandpass filter between 0.01 HZ and 0.1 HZ was applied to remove any baseline drift and physiological noise (e.g., heartbeats) after removing discontinuous shifts from the data time series. The [HbO2] of each group (the exercise group and the nonexercise group) was averaged for further analysis. Differences between the two groups were considered significant at p < 0.05. A false discovery rate correction was applied to minimize the effects of multiple comparisons.
Pearson correlation
We first assessed the Pearson correlation coefficient of all channels in both group and then used independent-sample t-tests to compare the differences between exercise and nonexercise groups.
Granger causality connectivity
Based on Pearson correlation analysis, we employed Granger causality analyses (GCA) to further estimate the direction of synchronization for channels that exhibited significant correlation. Although first developed in the field of economics (Granger, 2001), Granger causality connectivity has been widely used in the field of brain functional connectivity. This approach enabled us to compare the differences in a pair of channels or regions of interest and in the directional functional connectivity between the different groups (see more details in Barnett and Seth, 2014; Im et al., 2010). We assessed Granger causality with the Hermes tool kit (http://hermes.ctb.upm.es/; for procedural details see Zhang et al., 2017) to analyze brain functional connectivity. Plots of Granger causality for the two groups were made using the tool introduced by Xia et al. (2013). Independent-sample t-tests were used to compare the differences in Granger causality of all channels between the exercise and nonexercise groups.
Results
Participants
Initially, 22 participants met the inclusion criteria and took part in this experiment; however, two participants were excluded from the analysis because one sneezed during the resting-state fNIRS examination and the other did not complete the procedure. Therefore, 20 participants (16 women, 80.0%) were included in the final analysis. The ages of the participants ranged from 55 to 69 years (mean, 60.22 years; SD, 4.50 years). The participants were placed into one of two groups according to their exercise habits. Ten participants were included in the nonexercise group, and ten participants, who had exercised regularly for more than 1 year, were included in the exercise group. The average exercise time of the two groups was significantly different in past 1 year, the average exercise time (min) per week (M ± SD) of exercise groups (240.5 ± 80.01) was significantly higher than nonexercise group (19.50 ± 22.41; p < 0.05) (Table 1). To avoid potential confounding, we balanced the two groups for age, body mass index, and educational level, and none of these factors as well as sex percentage was found to be significantly different between the two groups (Table 1). All participants were nonsmokers, and none of them regularly consumed alcohol.
Demographic Characteristics, BMI, MMSE, and Odor Threshold of Exercise and Nonexercise Groups
Bold font indicates statistical significance. BMI, body mass index calculated as weight in kilograms divided by height in meters squared; MMSE, Mini-Mental State Examination; SD, standard deviation.
Behavior
MMSE scores
The mean (SD) MMSE score was significantly higher in the exercise group (27.89 ± 1.45) than in the nonexercise group (26.22 ± 0.83; p < 0.01) (Fig. 2; Table 1).
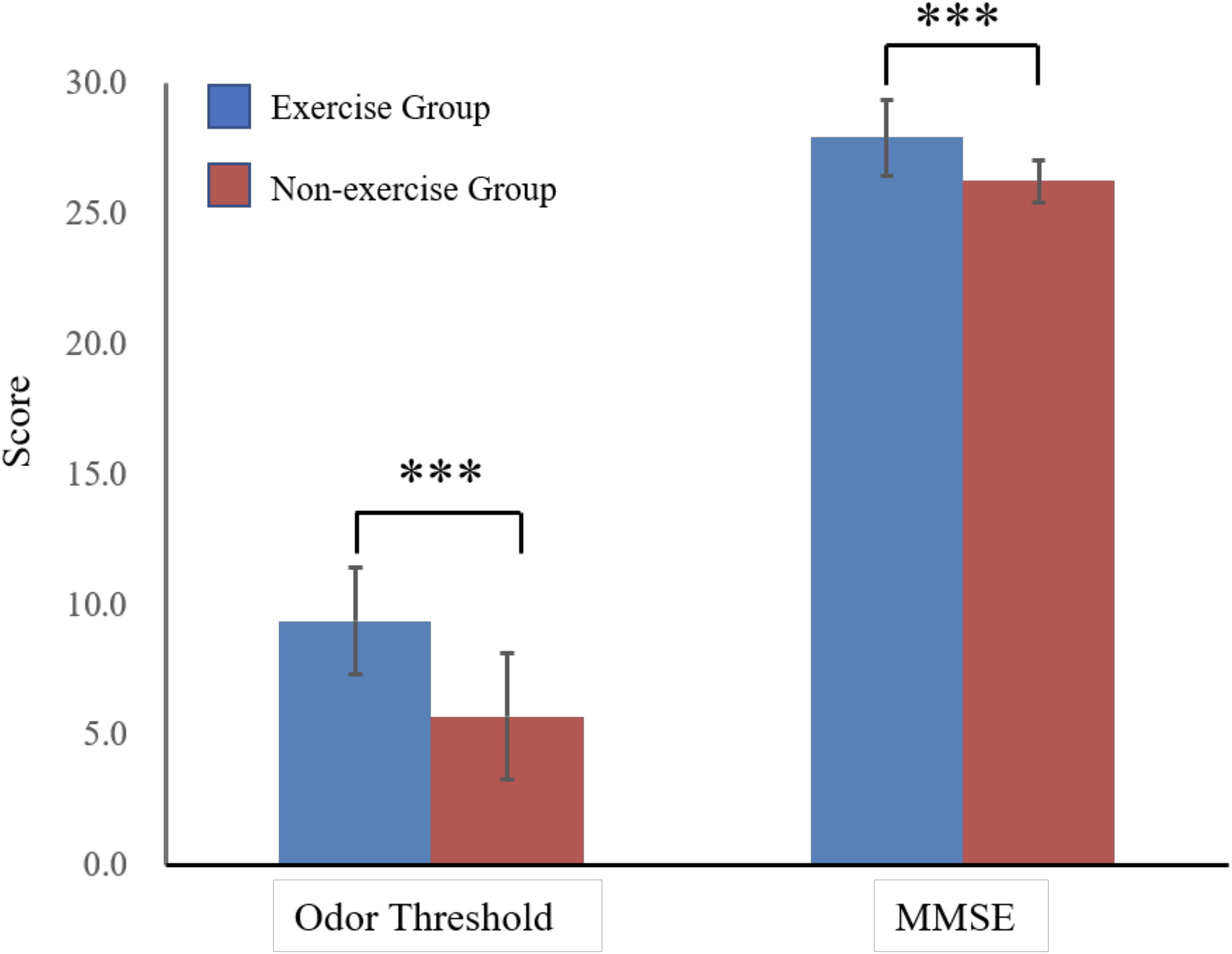
Odor detection threshold scores and Mini-Mental State Examination (MMSE) scores of participants who exercised regularly and of those who did not exercise. ***p < 0.001.
Odor threshold scores
The mean (SD) odor threshold score was significantly higher in the exercise group (9.38 ± 2.04) than in the nonexercise group (5.72 ± 2.43; p < 0.01) (Fig. 2; Table 1), indicating that the exercise group had a significantly lower threshold for detecting odors.
fNIRS results
Pearson correlation between channels
We found that the exercise group had significantly higher correlations than the nonexercise group between the following channels: channels 3 and 13, channels 3 and 12, channels 3 and 17, channels 4 and 11, channels 4 and 12, channels 7 and 16, channels 10 and 19, channels 11 and 12, and channels 11 and 13 (Fig. 3), for detailed values see online supplementary data, all results were FDR corrected.
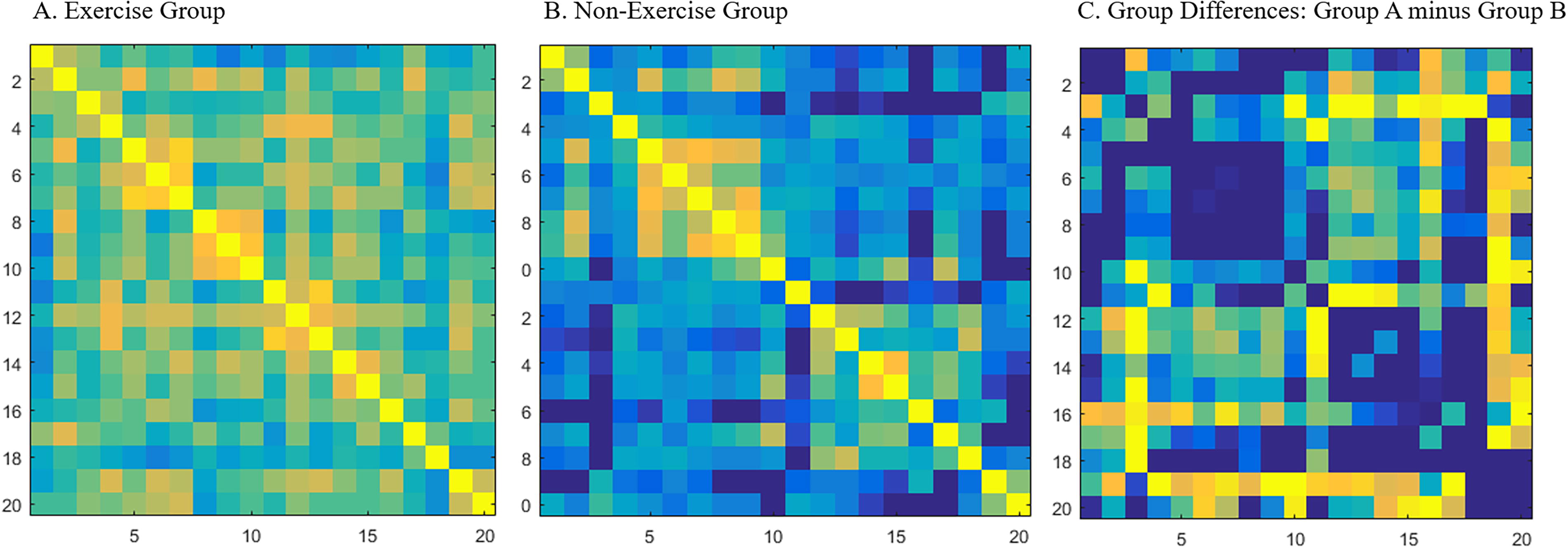
Grouped-averaged correlation matrix maps for the exercise group
Pearson correlations between brain regions
After associating the channels with the appropriate brain regions (see Fig. 1), we found that the significant differences between the exercise and nonexercise groups were mainly between the VLPFC and the OFC/DLPFC/FPA, between the DLPFC and the FPA/DLPFC, between the OFC and the FPA, and between the left and right hemispheres of the OFC. The exercise group had significantly higher correlations in these brain regions than the nonexercise group.
Granger causality in channels
Because Granger causality can reflect directional coupling, there are two directions between each pair of channels. For example, for channel 1 and channel 2, one direction is from channel 1 to channel 2, and the other direction is from channel 2 to channel 1. We analyzed Granger causality first from the lower-numbered channel to the higher-numbered channel, (e.g., from channel 1 to channel 2) and then from the higher-numbered to the lower-numbered channel (e.g., from channel 2 to channel 1).
We found a significant effect of exercise in first direction for the following channels (Fig. 4): from channels 3 to 6, channels 3 to 7, channels 4 to 6, channels 2 to 13, channels 2 to 16, channels 8 to 16, channels 2 to 19, channels 5 to 20, channels 7 to 8, channels 8 to 9, channels 9 to 20, and channels 9 to 17, for detailed values please see online supplementary data, all results were FDR corrected.
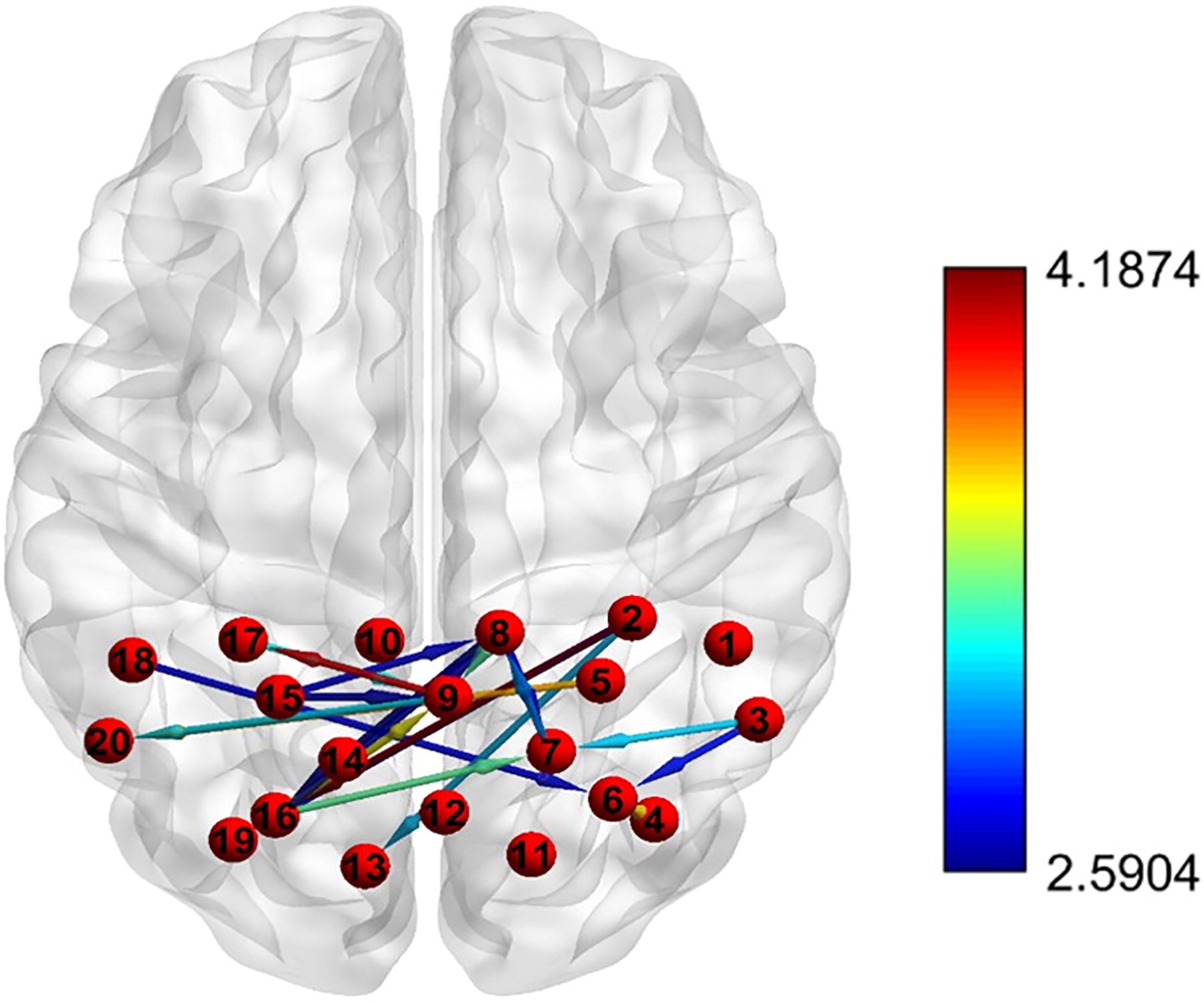
Directional coupling differences between the exercise and nonexercise groups. Connectivity strength is represented by the color of the arrow, which corresponds to the color scale shown on right. Warmer colors (e.g., red) represent stronger connectivity than cooler colors (e.g., blue).
There was significantly higher functional connectivity in the exercise group than in the nonexercise group for the following channels in the opposite direction (see Fig. 4): from channels 8 to 7, channels 15 to 8, channels 15 to 9, channels 17 to 9, channels 16 to 7, channels 16 to 8, channels 16 to 9, and channels 18 to 6, for detailed values see online supplementary data, all results were FDR corrected.
Granger causality shows higher functional connectivity from DLPFC to olfactory cortex in exercisers
After associating the channels with the appropriate brain regions, we found that the exercise group had higher functional connectivity than the nonexercise group in the following brain areas: from DLPFC to OFC/FPA/VLPFC/DLPFC, from VLPFC to DLPFC/FPA, and from FPA to DLPFC.
Correlations among different variates
We compared the associations between the behavior variates and thefNIRSHbO2 concentration in different channels for the two groups, especially for channels 11 and 13. These two channels were located in the OFC, an area considered to be a critical part of the olfactory cortex (Mai and Paxinos, 2011).
The association between olfactory threshold scores and MMSE scores was not significant (r = 0.40, p = 0.08). Significant correlations were detected between the olfactory threshold score and the HbO2 concentration in both channel 11 (r = 0.53, p = 0.017) and channel 13 (r = 0.55, p = 0.013) (Fig. 5). No significant association was found between the MMSE score and the HbO2 concentration in any channel. This latter finding may be because most of the participants (70%) were cognitively healthy (MMSE score ≥27), and for those few participants whose MMSE score was under 27, three of them scored 26, and three of them scored 25.
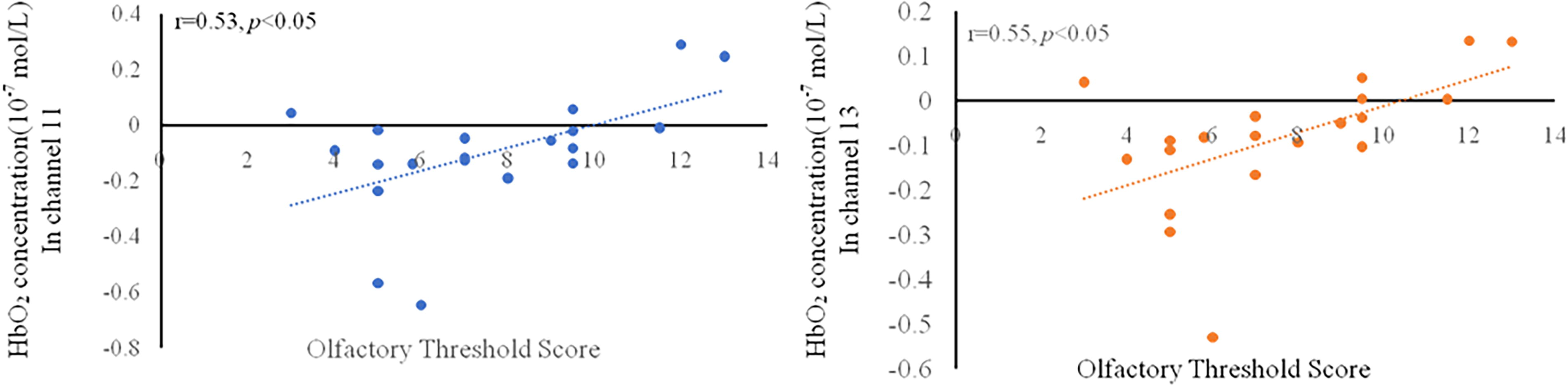
Correlation between olfactory threshold score and HbO concentration in channel 11/13.
To further explore the relationships among exercise, MMSE score, and olfactory threshold score, moderation and mediation analyses were employed. A significant direct effect of exercise on the olfactory threshold score was found both in the mediation analysis (β = −132.33, p < 0.001, Fig. 6A) and the moderation analysis (β = −146.82, p < 0.01, Fig. 6B). There were no significant indirect effects of the MMSE score on the olfactory threshold score in neither the mediation analysis (β = −0.76, p = 0.40, Fig. 6A) nor the moderation analysis (β = −0.93, p = 0.39, Fig. 6B).

Indirect effect of MMSE score on olfactory threshold score.
Discussion
To our knowledge, the present study is the first to use fNIRS to explore the effect of exercise on functional connectivity in the resting state between olfactory-related brain regions and other brain areas. The results suggested that the exercise group had significantly better olfactory sensitivity and stronger functional connectivity, both from olfactory-related cortices to the PFC and within the OFC.
Better behavioral performances in olfaction and cognition in the exercise group
Both MMSE scores and olfactory sensitivity were better in the exercise group than in the nonexercise group. Several studies have suggested a positive effect of exercise on the olfactory threshold. Schubert et al. (2017) found a close association between exercise and the odor threshold, showing that individuals who exercise have a lower risk of olfactory dysfunction years later. A previous study by our group also suggested better olfactory sensitivity in individuals who exercise (Zhang et al., 2020), consistent with our present results. As reflected by the MMSE scores, the present study results also showed that cognitive function was significantly better in the exercise group than in the nonexercise group. Previous evidence that exercise improves cognitive function is robust (Alderman and Olson, 2014; Angevaren et al., 2005; Chang et al., 2015).
Stronger functional connectivity in the exercise group
Stronger functional bidirectional connectivity between the olfactory cortex and the PFC
Both Pearson correlation and GCA showed a higher functional connectivity in the exercise group than in the nonexercise group. Pearson correlation analysis revealed strong connectivity between the VLPFC and OFC, between the OFC and the FPA, and also within the OFC (i.e., between the left and right hemispheres of the OFC). The results of the Granger causality indicated a higher directional connectivity from the DLPFC to the OFC in individuals in the exercise group than in those in the nonexercise groups.
Stronger functional connectivity between other brain regions in the exercise group
The results of Pearson correlation analyses indicated a stronger connectivity between the VLPFC and DLPFC/FPA and between the DLPFC and FPA/DLPFC in the exercise group than in the nonexercise group. In addition, Granger causality revealed that the exercise group exhibited a stronger directional communication than the nonexercise group from the DLPFC to the FPA/VLPFC/DLPFC, from the VLPFC to the DLPFC/FPA, and from the FPA to the DLPFC.
Previous studies have indicated that the DLPFC plays an important role in cognitive control and emotional regulation. This brain region has been associated with working memory (Braver et al., 1997) and episodic memory retrieval (Buckner et al., 1999; Rugg & Wilding, 2000; Wagner, et al., 1998) as well as with goal setting, plan monitoring, modulating anticipation, and using feedback in task performance (Bonelli & Cummings, 2007). Moreover, the DLPFC actively participates in the regulation of emotion (Berlim et al., 2013). The VLPFC was found to be abnormally activated in individuals with schizophrenia during empathy exploration (Derntl et al., 2012; Lee et al., 2010). Abnormal activation of the VLPFC has also been associated with the “jumping to conclusions” reasoning bias (Andreou et al., 2018) and auditory–verbal hallucinations (Moseley et al., 2013). “Jumping to conclusions” means people make decisions without understanding or fully considering all the available information. All these findings suggest that the VLPFC is critical for both psychopathology and social cognition. Located at the forefront of the PFC, the FPA has a vital function to support complicated cognitive processes, such as multitasking, high-level goal maintenance, and social cognition (Burgess et al., 2007). Given these findings, our present results suggested that exercise may not only improve olfactory performance but also may be involved with cognitive and emotional processing brain regions.
Correlations among different variates
We found a significant association between olfactory threshold scores and HbO2 concentrations in channels 11 (r = 0.53, p < 0.05) and 13 (r = 0.55, p < 0.05). Both channels were located in the OFC, a critical area for olfactory processing. The OFC plays an important role in judging odor familiarity, odor valence, and odor intensity (Kobayashi et al., 2010; Petersen et al., 1998; Plailly et al., 2011). Our results suggested that the OFC was also associated with the olfactory threshold. Although several previous studies have shown that poor olfactory function predicts cognitive impairment (e.g., Devanand et al., 2015), the association between olfactory threshold scores and MMSE scores was not significant in our study. No significant association was found between MMSE scores and HbO2 concentrations in any channel in the present study. This apparently discrepant finding may have occurred at least in part because the participants in the present study had relatively high MMSE scores. The findings of our previous study (Zhang et al., 2020), which included 99 healthy participants, support that when the average MMSE score is above 25, the relationship between the MMSE score and other variates may become complex.
In our present study, we used fNIRS to explore the functional connectivity between the olfactory cortex and other brain regions in individuals who exercised regularly and those who did not. Our results suggested that compared with individuals who did not exercise, those who exercised regularly not only had better olfactory performance but also showed evidence of stronger functional connectivity between the olfactory cortex and three subregions of the PFC, including the DLPFC, VLPFC, and FPA. These three brain regions actively participate in cognition and emotion processing. In addition, Gupta and Bahmer (2019) demonstrated that increased functional connectivity may contribute to perception, and based on this notion, stronger functional connectivity in the exercise group may lead to better olfactory performance, which is consistent with our results. Our findings suggested that exercise was highly associated with olfactory performance, the exercise group had higher activation in olfactory cortex and strengthened connectivity with cognition- and emotion-related cortices. Given these findings, our results provide a new perspective on why olfaction is highly correlated with cognition (Roalf et al., 2016).
However, our study also has several limitations that should be considered. First, our sample size is relatively small, with a total of only 20 samples. Despite we reported the effect size, the limited sample size is undoubtedly a significant drawback of this research. Hence, our findings should be regarded as indicative rather than definitive. Future studies should include more participants to confirm our findings.
Second, owing to the limitations of fNIRS, we were unable to detect the deeper olfactory areas, such as the piriform cortex entorhinal cortex and the limbic system. However, given that this is, to our knowledge, the present study was the first study to use fNIRS to examine the association of exercise with olfaction, we believe some new insights have been gained. Third, future research should include exercise interventions to determine whether a causal relationship exists between exercise and olfaction. Fourth, participants were assigned to the exercise and nonexercise groups based on their self-reported amount of time spent in exercise. Future studies should consider a more objective method to distinguish the two groups. Finally, we used the Sniffin’ Sticks Test to examine participants’ odor threshold; olfactometers should be considered to assess olfaction in future studies.
Conclusion
In conclusion, we found that compared with individuals who did not exercise, those who exercised regularly not only had better olfactory performance but also showed evidence of stronger functional connectivity between the olfactory cortex and three subregions of the PFC that actively participate in cognitive and emotional processing.
Footnotes
Authors’ Contributions
X. W. and C. Z.: Involved in conceptualization. C. Z.: Conducted the research and formal analysis and wrote the original draft. X.W.: Incharge of funding acquisition and primary responsibility for the final content. All authors have read and approved the final article.
Data Availability
The authors declare that the datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Shanghai Science and Technology Planning Project [grant number