
Abstract
Background:
Cerebral palsy (CP) often affects function of one or both arms. Resting-state magnetic resonance imaging studies identified abnormal neuronal connectivity related to functional deficits in CP, with few studies on dynamic, task-related changes in connectivity. Here, we compare connectivity in participants with CP and typical development (TD) during an upper limb task and relate these to motor performance.
Methods:
Children with CP (n = 15) and TD (n = 15) performed a button-press task with both arms, while recording 64-channel electroencephalography. Inter- and intrahemispheric connectivity between dominant and nondominant premotor, motor, and sensory regions were examined during rest, movement preparation, and execution using a normalized magnitude squared time-frequency coherence analysis (α-band: 8–12 Hz, β-band: 13–35 Hz, γ-band: 36–85 Hz).
Results:
The only group differences were in intrahemispheric connectivity during nondominant arm trials, with CP having higher frontal to central connectivity than TD in all frequency bands in the dominant hemisphere and higher central to parietal beta connectivity in the nondominant hemisphere. Significant main effects for period showed most differences between rest and movement phases. Group by period interactions were also only found during nondominant arm trials (interhemispheric: CP coherence increased more during execution in frontal, central, and parietal regions; intrahemispheric: CP coherence decreased less during execution in nondominant and dominant frontal to parietal regions). Clinical and movement scores were moderately related to connectivity in CP, with poorer nondominant arm function significantly correlated with higher inter- and intrahemispheric coherence.
Conclusions:
Group differences emerged mainly during intrahemispheric nondominant arm trials across frequency bands with higher coherence in CP associated with greater functional limitation.
Impact Statement
In contrast to assessing brain connectivity with MRI in children with CP, the use of EEG enables the investigation of this during a functional task, and the sample is not limited by head movements that preclude the attainment of high-quality MRI data in many with CP. The finding of increased task-specific intrahemispheric brain connectivity in bilateral CP, the magnitude of which was related to the degree of functional limitations, suggests a new target for rehabilitation as well as a sensitive outcome measure for clinical trials aimed at improving brain and motor function in CP.
Introduction
Cerebral palsy (CP) often affects the use of one or both upper limbs, resulting in significant functional motor impairments (Østensjø et al., 2004). Spastic CP is the most prevalent subtype of CP (Himpens et al., 2008), which can be further categorized by distribution as unilateral or bilateral. Those with bilateral CP typically have more limited mobility, with more than half classified as Gross Motor Function Classification Scale (GMFCS) level III or higher, whereas 90% of individuals with unilateral CP are classified as level I or II (Gorter et al., 2004). Regardless of the subtype, motor deficits and the resulting functional limitations can have a significant impact on the quality of life of individuals with CP (Tarsuslu and Livanelioglu, 2010).
The brain mechanisms underlying impaired neural control in CP have been investigated primarily with functional magnetic resonance imaging (fMRI) during resting state (Woodward et al., 2019), voluntary movements (Van de Winckel et al., 2013), or responses to sensory stimuli (Wingert et al., 2010), although several studies have utilized magnetoencephalography (MEG) (Kurz et al., 2015) and more recently functional near-infrared spectroscopy (de Campos et al., 2020) or electroencephalography (EEG; Inuggi et al., 2018). However, data are more limited in those with bilateral CP than unilateral CP. In addition, the observed patterns of neural abnormalities in CP and their relationships with impairments appear to be inconsistent across studies, with some studies reporting an increased level of activation in motor-related brain regions in bilateral CP during motor tasks (Kurz et al., 2017; Hinchberger et al., 2023), which was associated with greater deficits, while others finding that an increased level of brain activation in CP was related to better function (Phillips et al., 2023). Conversely, a significantly lower level of brain activation in bilateral CP, specifically in the high-frequency band, was also observed when compared with age-matched controls in other studies (Kurz et al., 2014; Hoffman et al., 2019).
Measures of task-related intracortical connectivity, shown to be abnormal in CP (Rose et al., 2011), may provide a more coherent and dynamic representation of differences in regional brain activation patterns observed in bilateral CP. Structural brain connectivity (diffusion tensor imaging) and functional connectivity (fMRI or EEG), both measured during resting state, were altered in unilateral CP (George et al., 2021; Pannek et al., 2014; Weinstein et al., 2014; Papadelis et al., 2019; Woodward et al., 2019) and bilateral CP (Papadelis et al., 2014; Englander et al., 2015; Ballester‐Plané et al., 2017; Qin et al., 2018; Damiano et al., 2022; Vallinoja et al., 2024). These were generally characterized by lower directionality (e.g., lower functional anisotropy) and efficiency (i.e., higher radial diffusivity), representing restrictions and/or inefficiency in neural communications.
Differences in structural and/or functional connectivity, however, still may not fully explain the mechanisms underlying the functional deficits in bilateral CP that manifest during task performance (i.e., during movements). Instead, effective connectivity (Friston, 1994, 2011; Horwitz, 2003) may provide greater insights into the neural control of movement in CP, as it evaluates time-dependent changes in connectivity during task performance (Aertsen and Preißl, 1991; Friston, 2011). Distinct task-related changes in connectivity during movement preparation and execution can be measured and have been utilized previously to characterize movement deficits in other neurological disorders, for example, Parkinson’s disease (Wu et al., 2010, 2011), stroke (Palmer et al., 2019; Hoshino et al., 2021), and/or dystonia (Jin et al., 2011; Thirugnanasambandam et al., 2020). Significant activity-dependent changes in neural communication, if observed, could lead to more specific characterizations of the relationships between cortical activity and functional impairment in bilateral CP.
The goal of this study was to examine dynamic changes in brain connectivity in CP during a functional upper extremity task (i.e., a button-press task) using EEG. In particular, we examined inter- and intrahemispheric brain connectivity during movement preparation and execution, which are independent processes with distinct neural bases (Haith et al., 2016). We examined the coherence measures in the α-, β-, and γ-bands, which are commonly used to gauge the degrees of communication/connectivity between brain regions for sensorimotor processing during motor task (Manganotti et al., 1998); coherence in these bands was also used to characterize impaired sensorimotor processing in different neurological disorders such as stroke (Wu et al., 2015; Zhou et al., 2021; Vecchio et al., 2023), Parkinson’s disease (Cassidy and Brown, 2001; Oswal et al., 2013), and dystonia (Jin et al., 2011; Lagerweij et al., 2024). We hypothesized that abnormal cortical connectivity in CP would be identified in both the preparation and execution periods, as a significant degree of impairment in upper limb motor planning was previously found in CP (Martinie et al., 2021). Notably, a significant between-group difference (CP vs. neurotypical) in brain potentials (i.e., event-related synchronization and/or desynchronization) was found during both the planning and execution periods of lower extremity (Kurz et al., 2014) and upper extremity (Inuggi et al, 2018) movements. We also hypothesized that the degree of abnormal brain connectivity in both periods would correlate directly with the magnitude of functional impairment.
Materials and Methods
Participants
The study protocol was approved by the Institutional Review Board of the National Institutes of Health (Protocol #13-CC-0110). Informed assent and consent were obtained from each participant and his/her parent, respectively. Inclusion criteria were a diagnosis of bilateral CP, age 5–17 years, and ability to reach and grasp objects with both hands (Manual Ability Classification System [MACS] levels I–III). Children with uncontrolled seizures were excluded, and all were asked to refrain from caffeine for 24 h prior to the experiment.
A total of 32 participants, 16 with bilateral CP (9 females; mean age: 13.6 years; range: 6.5 years to 17.4 years) and 16 age-matched peers with typical development (TD; 7 females; mean age: 13.8 years; range: 8.1 years to 18.2 years), were recruited. Mean age was not significantly different between groups (p = 0.40). Data from two participants, one in each group, were later excluded from the analysis, as a majority of their trials had to be removed after EEG preprocessing because of noise.
The etiology of bilateral CP was periventricular leukomalacia secondary to preterm birth for the majority of participants. Demographic data for participants are summarized in Table 1.
P
CP, cerebral palsy; GMFCS, Gross Motor Function Classification Scale; IVH, Intraventricular Hemorrhage; MACS, Manual Ability Classification System; MRI, magnetic resonance imaging; PVL, periventricular leukomalacia; TD, typical development.
Instrumentation
A custom-made apparatus containing three buttons was used for the button-press task. Each button (11 cm [H] × 9.8 cm [W]] was secured on a wooden block (90.6 cm [L] × 13.7 cm [W]) and spaced evenly apart (27 cm between the centers) (Fig. 1). Each button was surrounded by light-emitting diodes secured under a rubber panel. This apparatus was placed on the center of the table.

Experimental set-up showing a participant resting the hand on the table (initial position) as an Light-Emitting Diode (LED) button was activated.
A 12-camera motion capture system (Vicon Motion Systems, Denver, CO) was used to record hand movements. Reflective markers were placed bilaterally on the dorsal midpoints of the wrists, over the second and fifth metacarpals, and on the fingernails of the thumb and index fingers. Digital videos were also captured to evaluate task performance.
A 64-channel, wireless, active EEG system (Brain Products, Morrisville, NC) with electrodes positioned in a snug fitting cap according to the 10–20 international system was used to record brain signals. Electrode impedance for each channel was maintained below 20 kΩ. EEG data were referenced to FCz and collected at 1000 Hz.
Protocol
The Box and Block Test (BBT) of upper limb coordination, which has been widely used to assess upper extremity function in neurotypical children (Mathiowetz et al., 1985) and in those with CP (Araneda et al., 2019), was first administered for each hand for all participants before the experiment.
During the button-press task, participants initially rested the designated hand on the start position indicated by a small “x” mark on the table (see Fig. 1). The table height was adjusted for each participant so that the elbow flexion angle was approximately 90°, with the hand in the resting position, i.e., palm resting face down on thigh, with the forearm pronated. Participants were asked to press the button that lit up as quickly as possible and then return the hand to the start position. A microcontroller randomly activated one of the three LEDs, which was deactivated as soon as they pressed the button. Variable inter-stimulus intervals of 4 to 8 sec were used between trials. Participants performed two sets of 25 trials (50 total) with each hand, and the order of the hand to be used was randomized across participants. A pulse was delivered via a trigger to the microcontroller for data synchronization.
Children rested between hand trials and were also told that they could ask to rest at any time during the experiment as needed. Sessions were conducted during the day, between 9:00 a.m. and 3:00 p.m., depending on family preference and under controlled conditions (standardized instructions and identical set-up for each participant in a quiet room with minimal distractions) that were consistent throughout the study.
Data analysis
EEG analysis
EEG data were preprocessed in MATLAB using EEGLAB toolbox (Delorme and Makeig, 2004), as detailed in our previous publications (Hinchberger et al., 2023). Briefly, these steps include line noise removal (60/120 Hz), removal of bad channels, creation of a merged file including all conditions, visual inspection, and removal of noise periods, downsampling to 250 Hz, application of Artifact Subspace Reconstruction, re-referencing to a common average, and 1 Hz high-pass filtering. We then applied the independent component analysis weights computed from processed data to the individual datasets for each condition. ICLabel, a machine-learning algorithm that generates a percent confidence for each independent component (IC) regarding its origin (cortical, ocular, muscular, or artifactual), was applied, and ICs deemed of noncortical origin were removed. Time series data were then epoched from 2 sec (t = −2 sec) before the movement initiation (which was set to 0 sec) to the time point when the button was pressed (t = tb ).
Corticocortical connectivity was assessed via coherence between EEG channels in the following three areas, i.e., premotor (left frontal: F1, F3, F5; right: F2, F4, F6), motor (left central: C1, C3, C5; right: C2, C4, C6), and sensory (left parietal: P1, P3, P5; right: P2, P4, P6), resulting in three pair groups for each hemisphere (intrahemispheric: F-C, F-P, C-P) and three pair groups between hemispheres (interhemispheric: F-F, C-C, P-P). Each pair group contained nine electrode pairs (3C1 × 3C1 = 9).
For each pair of electrodes within each group, the time-dependent EEG–EEG coherence values were estimated using a sliding window of 250 msec with an increment of 100 msec, within the MATLAB environment (MathWorks, Inc., Natick, MA) employing a script by Neurospec (www.neurospec.org) (Halliday et al., 1995). Briefly, the magnitude-squared coherence at frequency
(
Coherence values above the 95% confidence level were then z-transformed to yield normally distributed values with a standard deviation of approximately 1 (Rosenberg et al., 1998).
The z-transformed coherence was estimated within each of the three time periods, i.e., baseline (TP1): before stimulus, −2 sec < t < −1 sec; movement preparation (TP2): between stimulus (ts
) and movement initiation,
Kinematic analysis
Movement time (MT) was computed from the kinematic data obtained from the motion capture system. Reach start was defined as when hand velocity first exceeded 0.05 m/sec following rest, and reach end was defined as when the button was pressed. Reach time was calculated as the time from reach start to end.
Statistical analysis
To identify between-group differences in the connectivity measures, a general linear mixed model with repeated measures was performed on the peak coherence values for each intra- and interhemispheric pair, with the three time periods as a within-subject variable and the group as a between-subject variable (TD or CP). Here, a multivariate analysis was first performed on the three coherence values in the three frequency bands (α-, β-, and γ-bands), which was followed by separate univariate tests on the peak coherence values in individual frequency bands if the result of the multivariate analysis was statistically significant (p < 0.05).
To examine within-group variability for the connectivity measures in CP and their relationship with functional impairments and age, Pearsonr correlation tests were performed. We also examined potential relationships between age and the clinical scores.
Results
Clinical measures and kinematic outcomes
BBT scores for individuals with CP were significantly lower than those of TD (nondominant: CP 31.2 ± 17.2, TD 57.7 ± 7.0; p < 0.01; dominant: CP 39.7 ± 14.2, TD 57.0 ± 7.9; p < 0.01). Their movements were also significantly slower (nondominant: CP 0.69 ± 0.42 sec, TD 0.43 ± 0.08 sec; p = 0.02; dominant: CP 0.52 ± 0.17 sec, TD 0.42 ± 0.07 sec; p = 0.04) (Table 2).
B
CP, cerebral palsy; TD, typical development.
General linear model results
All participants were able to complete the task accurately with both hands. Trials where the participant failed to turn off the correct button in the allotted time or needed to hit it more than once to turn it off were eliminated. The group with TD completed a mean of 40.7 and 43.6 trials and the group with CP a mean of 38.4 and 37.8 trials with the dominant and nondominant hands, respectively, out of a total of 50 trials. There were no significant differences across hands (p = 0.14) or groups (p = 0.61) or a group by hand interaction (p = 0.61).
Interhemispheric connectivity
No significant group differences were found in interhemispheric connectivity. For both groups, movement period had a significant impact on interhemispheric connectivity, as the coherence values of all three pairs (F-F, C-C, P-P) increased significantly over the course of the task (all p values <0.01), with most of the changes between rest and the other two periods. A general pattern was seen across groups and hands with a decrease in coherence from rest to preparation and then a minimal or small increase with movement execution.

Interhemispheric coherence values during

Representative time-frequency plots of the intrahemispheric
G
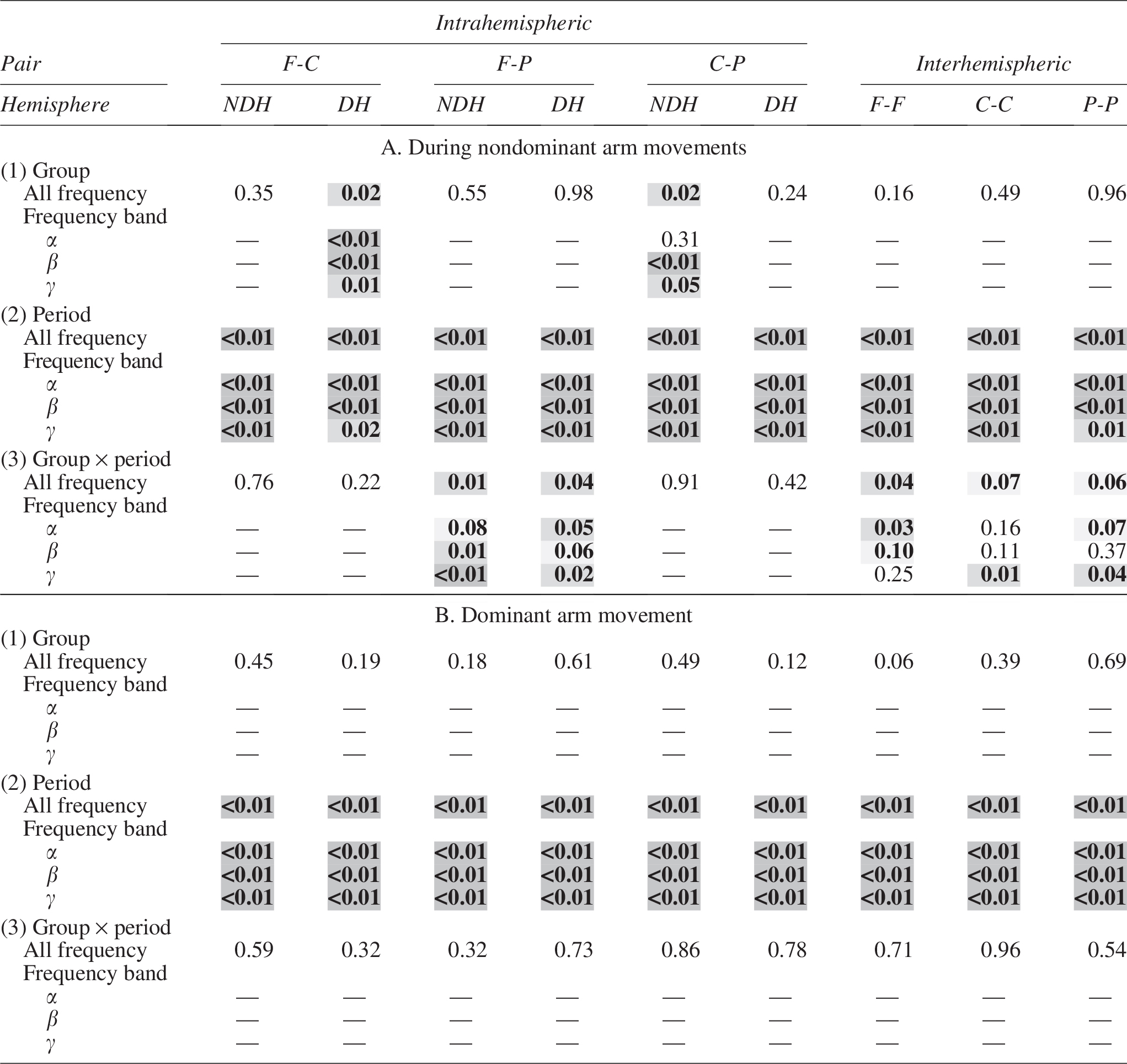
Shown are the p values for (1) group effect, (2) period effect, and (2) group × period interaction during A. nondominant arm movements and B. dominant arm movements. NDH, Nondominant Hemisphere; DH, Dominant Hemisphere.  p < 0.1;
p < 0.1;  p < 0.05;
p < 0.05;  p < 0.01.
p < 0.01.

Intrahemispheric coherence values during
Within-group correlations of connectivity with motor performance and age
MT
For individuals with CP, the between-subject variability in MT for the nondominant arm could be explained in part by the variability in the movement-related increases in intra- and interhemispheric connectivity, quantified by the correlation coefficient values (Table 4A). In CP, significant positive correlations between movement duration and peak coherence values were found in both contralateral and ipsilateral pairs including the motor areas (F-C and C-P), and the differences were observed mainly in the lower frequency bands (α and β) (Table 4A; Fig. 5). Significant correlations were found during dominant hand trials in two cases (α-band of F-C pair in the dominant hemisphere and γ-band of C-P pair in the nondominant hemisphere during TP3; Table 4B) in CP.

Correlation between the intrahemispheric coherence (F-P and C-P of nondominant hemisphere) for 16 participants with CP and movement time for the nondominant arm. Individuals with CP who exhibited higher peak coherence values took longer to complete the movements.
P
Shown are r values (p-values) between peak coherence values of intra- and inter-hemispheric pairs and movement completion times for participants with CP.
BBT
Between-subject variability in the clinical score (BBT) was also explained in part by the intra- and interhemispheric connectivity values across participants with CP. Significant inverse correlations between BBT scores and coherence values of CP subjects were also found in the two pairs, mostly in the lower band (α) (Table 5; Fig. 6).
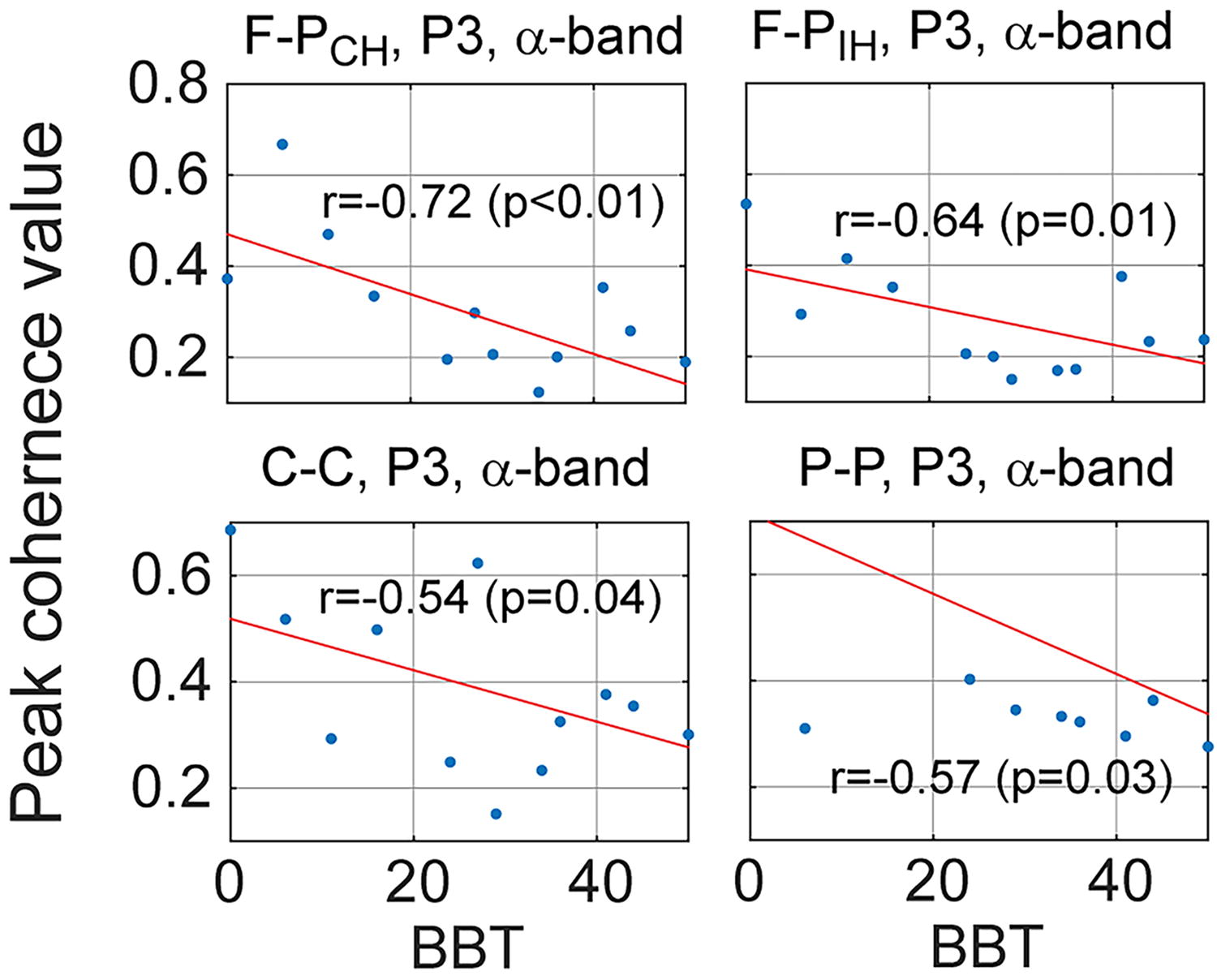
Correlation between the F-P peak coherence values of 16 participants with CP and their Box and Block Test scores such that those with lower Box and Block Test scores tended to exhibit higher peak coherence values before and during movements.
P
Shown are r values (p-values) between peak coherence values of intrahemispheric and interhemispheric pairs and the Box and Block Test (BBT) scores in participants with CP.
Age
Age was significantly correlated with the connectivity measures (i.e., peak inter- and intrahemispheric coherence) only in 50 out of 324 cases (15.4%). In addition, age was correlated with neither MT (r = 0.17, p = 0.53 for TD; r = 0.09, p = 0.72 for CP) nor clinical score (BBT) (r = 0.33, p = 0.28 for TD; r = −0.24, p = 0.39 for CP) in both groups.
Discussion
The goal of this article was to quantify functional brain connectivity in a cohort with bilateral CP compared with those with TD during an upper limb button-press task using EEG. As hypothesized, we found greater intrahemispheric connectivity in bilateral CP in multiple frequency bands only when using the nondominant hand. In the nondominant hemisphere, the C-P coherence was greater in CP in the β-band, indicating the greater difficulty with the nondominant hand in motor execution. F-P connectivity increases in CP were seen in all three bands in the dominant hemisphere, indicating greater reliance on the ipsilateral hemisphere in CP when planning, executing and coordinating movements with the nondominant hand. Furthermore, we demonstrated that increased connectivity in CP was directly correlated with poorer motor performance. There were no significant group differences in interhemispheric or intrahemispheric coherence in dominant hand trials.
We have previously quantified EEG coherence or connectivity during quiet standing and found that it was greater on the nondominant (injured) hemisphere in a group with unilateral CP with no differences across sides in those with TD (George et al., 2021). In that same cohort, greater activation was seen in CP in multiple frequency bands when walking, even though temporal–spatial gait data were similar across groups (Short et al., 2020). Our previous EEG studies in children with bilateral CP have examined event-related changes in activation in the α- and β-bands during upper limb tasks. In those studies, children with CP tended to exhibit generally greater activity than those with TD when performing the same task, particularly when using the nondominant hand, which in some cases was linked to greater impairment (Hinchberger et al., 2023) and in other cases to greater function (Phillips et al., 2023).
The existing data on functional connectivity in bilateral CP are mostly based on resting-state MRI and vary across studies, which is not surprising given the small sample sizes and heterogeneous nature of CP. The most remarkable finding in the resting-state data from Burton et al. (2009) was the spatially extended connectivity maps in the two selected somatosensory parietal areas shown to be represented during finger movements in bilateral CP compared with TD, whereas the connectivity of other parietal areas (e.g., left and right Brodmann areas 1 and 2) was of either similar or smaller extent in CP. In the current study, the frontal to parietal intrahemispheric coherence was the one that demonstrated a group by period interaction with CP demonstrating less of a decrease from rest to movement preparation and from preparation to execution compared with those with TD, which may indicate loss or inefficiency of more distant pathways.
Mu et al. (2018) examined resting-state MRI in spastic bilateral CP and TD and found increased connectivity in CP in the primary and supplementary motor regions and between the thalamus, cerebellum, and precentral gyrus, but decreased connectivity within the basal ganglia and between the motor regions and the fusiform and lingual gyrus. Damiano et al. (2022) found no group difference in resting-state MRI; however, there was a large effect size for higher connectivity within the precentral gyrus across hemispheres in CP compared with those with TD. Data from Qin et al. (2018) are sharply in contrast to others in that they found decreased connectivity in the primary somatosensory, cerebellar, frontoparietal, and salience networks with lower connectivity related to poorer mobility as indicated by the GMFCS and lower Activities of Darily Living (ADL) scores. It is possible that methodological differences among these studies may have contributed to the discrepancies in their findings. Vallinoja et al. (2024) evaluated functional brain connectivity using both MEG and functional connectivity MRI in the same participants who were instructed to try not to think of anything in particular while focusing on cross hairs in both conditions. Results for functional connectivity MRI demonstrated significant increases within parietal clusters on each side in CP versus TD, with no significant increases in interhemispheric connectivity. MEG results, in contrast, showed significant increases in interhemispheric connectivity for both the α- and β-bands between the left parietal and right sensorimotor regions, and in the β-band between the right and left sensorimotor regions. The different findings across methods were unexpected, but the authors attributed these to the dramatically different time scales over which connectivity was computed for each method, even though coherence or connectivity is defined in each case as the temporal dependence of anatomically distinct brain regions. Given these methodological differences, it may only be valid to compare results across studies with similar methods.
EEG coherence studies in bilateral CP are very limited. A study by Koeda and Takeshita (1998) evaluated coherence in those with bilateral CP who had been born preterm and found that interhemispheric coherence in the occipital region was lower in the α-band than in TD but was elevated for inter- and intrahemispheric motor-related pairs in the β-band. A recent study by Pavlenko et al. (2024) used EEG coherence as an outcome measure for neurorehabilitation. While they did not discuss baseline coherence, they did find a relationship after training between higher coherence in the right C4 to CP04 regions when imagining left hand movement and higher Barthel scores.
Time-frequency analyses of EEG signals, as utilized in the current study, offer distinct advantages over previous studies on resting-state connectivity as these can detect task-relevant changes in connectivity over the course of task performance. Resting-state connectivity may be related to, but not fully representative of, the more dynamic brain processes during specific tasks. An obvious advantage of methods like MEG and EEG is the ability to evaluate task-specific activation more precisely with the capability to even quantify differences between the motor planning and execution periods as was done here. Each of the two distinct processes of motor planning and execution may be impaired in CP and thereby show increased connectivity compared with those with TD, although the results for these two periods may have different relationships with motor performance measures. Our results showed that for individuals with bilateral CP, interhemispheric brain connectivity in CP had more significant increases in the central and parietal regions from preparation to execution than in those with TD and greater C-P and F-C intrahemispheric connectivity. More importantly, the degree of abnormality was correlated with the degree of functional impairments assessed by spatiotemporal movement abnormalities and clinical scores. Thus, the outcome of this study provides a more ecologically valid account of the relationship between abnormal brain connectivity and functional impairment in CP.
There are several different mechanisms that may be associated with the correlation between connectivity and functional impairment in CP. CP often involves loss of brain gray and/or white matter, and there are multiple studies that show decreased brain volumes in motor-related regions using structural MRI and decreased fractional anisotropy seen in diffusion tensor imaging results because of diffuse white matter injury, so it seems almost counter-intuitive that coherence would have greater spatial extent in regions where the damage is presumed to have occurred. Possible explanations include increased intracortical connections to compensate for loss of descending pathways (Burton et al., 2009), recruitment of more brain resources indicating a loss in efficiency (Short et al., 2020), or heightened attentional resources during motor tasks to ensure better performance. Each of these would likely be needed to a greater extent in those with greater damage to the brain, thus supporting links to poorer function. There may also be cases in which the damage is too pronounced to enable these compensatory mechanisms, and therefore, less connectivity may be linked to poorer function (Schneider et al., 2021). Finally, gaining expertise in a motor skill may also lead to greater efficiency, in which case a decrease in connectivity may be associated with motor improvements (Woodward et al., 2019). Given that one or more of these scenarios are possible, interpretation of results is not always straightforward.
A few potential limitations of the study should be acknowledged. We enrolled children across a large age span, which could have contributed to the within-group variability in the connectivity and clinical measures; however, the groups were well-matched by age, so this unlikely contributed to the individual and within-group variability in the connectivity and clinical measures. As an additional step, we correlated each dataset with age, and fewer than 20% showed a significant correlation with age. Another limitation was that participants with CP had to have sufficient bilateral hand function to be included in the study, with all in the three most functional MACS levels and none in the two least functional ones. This likely reduced the number of significant results as well as the magnitude of the differences had it been possible to study individuals across the full spectrum of upper limb function in CP; however, we still demonstrated clear differences across groups as well as significant within-group correlations between brain activation and function. Lastly, in this study, we computed channel-level coherence values, as the number and/or spatial resolution of the identified source locations varied across subjects, which makes it difficult to perform connectivity analysis between all regions of interest. However, coherence values at the channel level could be affected/corrupted by volume conduction (Srinivasan et al., 2007; Haufe et al., 2013; Van De Steen et al., 2019). We found, however, that in most cases, the peak coherence was not found in the pairs with the shortest interelectrode distance, which would be more likely if the coherence was indeed affected by volume conduction. In addition, the time point at which the coherence peak was observed in many cases was not the time when the highest power was observed at individual electrodes. The observed changes in connectivity (i.e., observed coherence peaks) thus are not likely caused by volume conduction from the same artifacts/sources. Here, we chose to report peak instead of mean coherence, which is important to note when comparing across studies.
Conclusion
This study confirms previous MRI findings of increased intracortical connectivity in bilateral CP compared with an age-matched group with TD using EEG during a functional upper limb task. Increases in connectivity in the sensorimotor regions appear even more widespread in bilateral compared with unilateral CP, which is not surprising given the greater degree of neurological involvement typically seen in this subtype. Strong relationships to function found here further suggest connectivity increases as a potential mechanism underlying motor impairments in CP.
Footnotes
Authors’ Contributions
S.W.L.: Methodology, Data analysis (lead), Writing—first draft (lead); T.C.B.: Conceptualization, Investigation, Methodology, Supervision, Writing—review and editing; J.E.K.: Data analysis, Writing—review and editing; D.L.D.: Conceptualization, Investigation, Methodology, Project administration, Resources, Data analysis, Supervision, Writing (first draft, review and editing).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Intramural Research Program of the NIH Clinical Center (Protocol 13-CC-0110).