
Abstract
Abstract
The endocannabinoid system (ECS) favors the intake and storage of energy by acting through both central and peripheral mechanisms. In particular, the ECS is present in the adipose tissue, where endocannabinoids affect adipocyte proliferation, differentiation, and secretion. Stimulation of cannabinoid type 1 (CB1) receptors in white adipocytes increases glucose uptake and lipogenesis, while inhibiting lipolysis, mitochondrial biogenesis, and adipocyte transdifferentiation. Similarly, in brown adipocytes, CB1 activation decreases mitochondrial biogenesis and thermogenic capacity. Overall, ECS-driven processes in the adipocyte therefore lead to energy preservation and can contribute to obesity under conditions of energy surfeit. This review provides an up-to-date overview on the role of the ECS in adipocyte physiology and of its involvement in obesity and metabolic disease.
Introduction
Obesity is a multifactorial, chronic disease of epidemic proportion characterized by the pathological expansion of fat mass in the organism. This pathological increase in fat mass is due to the combination of size increase of preexisting adipocytes (hypertrophy) and de novo adipocyte differentiation (hyperplasia), leading to important endocrine and metabolic dysfunctions at the level of both the adipose organ and the organism as a whole.
The endocannabinoid system (ECS) plays a crucial role in favoring fat accumulation in mammals through various processes spanning from its direct action on the adipocyte to the stimulation of energy intake and the inhibition of energy expenditure via both central and peripheral mechanisms.1,2 This lipid-signaling system, which consists of two different G protein-coupled receptors, cannabinoid receptor type 1 (CB1) and cannabinoid receptor type 2 (CB2), their endogenous polyunsaturated fatty acids ligands, the endocannabinoids (ECs), and the enzymes involved in their biosynthesis and degradation,3,4 is ubiquitously located 5 and evolutionarily well preserved,6,7 suggesting its primary importance in the regulation of the biological mechanisms responsible for energy preservation. Consequently, the ECS is critically involved in the development of obesity, a condition in which this system is overactivated (reviewed in Refs.2,8,9) and it represents a target for treatment.2,9
In this review, we will describe the presence and function of the ECS in the adipose tissue, how the ECS in the adipocyte might participate to obesity and metabolic disease, and whether it might represent a therapeutic target for the treatment of these pathological conditions.
The adipose tissue
For many years the adipose tissue was considered as a static organ, an unsophisticated depot of fat designated to store energy and to thermally isolate the organism. 10 In 1994, a ground-breaking discovery unexpectedly changed this view: Zhang et al. identified the appetite-suppressant cytokine leptin 11 as a hormone secreted by the adipose tissue, unveiling for the first time the dynamic and endocrine nature of this organ. Since then, many fat-produced molecules, called adipokines, have been identified to have crucial effects in the regulation of energy balance and metabolism 12 and the active role of the adipose organ in physiology and physiopathology has been recognized.
The adipose tissue can be distinguished in white and brown adipose tissue (WAT and BAT, respectively), based on its location and on the type of cells, white or brown adipocytes that constitute the organ. More recently, a third cellular type, called beige or brite adipocyte, has been described. 13 The adipose tissue clearly does not uniquely contain adipocytes or their precursor cells. It also has an important vasculature and it is richly innervated. Blood vessels and nerves therefore connect the adipose organ to the organism and guarantee the integrated regulation of endocrine and metabolic responses.
The principal functions of white adipocytes are fat storage and adipokine secretion, conferring the endocrine feature of the adipose organ. 14 Morphologically, 90% of the cell volume is composed by a single fat droplet together with few mitochondria and a flattened nucleus. 15 In mammals, the WAT is mostly located in subcutaneous and visceral depots. 14 The accumulation of fat in the latter anatomical region is recognized as one of the most significant risk factors for metabolic and cardiovascular diseases, 16 while, in contrast, the subcutaneous fat exerts protective actions. 17
Differently from white adipocytes, brown adipocytes are morphologically characterized by a high number of mitochondria, several small fat droplets within the cytoplasm, and a rounded nucleus. 15 Their principal function is not energy storage, but the nonshivering thermogenesis, due to the expression of mitochondrial uncoupling proteins, such as UCP1, which uncouples the oxidative phosphorylation from the adenosine triphosphate (ATP) synthesis creating futile cycles resulting in the production of heat.18,19 This process is activated mainly through sympathetic innervation of the BAT, with norepinephrine acting on β3-adrenoceptors expressed on the brown adipocyte surface. 18 In mice, the BAT has an interscapular, subscapular, and cervical location, 20 while in adult humans, it is principally located in the neck and upper chest regions. 19
The third type of adipocytes, the beige adipocytes, has been recently discovered.13,21 They are morphologically brown-like adipocytes, presenting multiple fat droplets and expressing UCP1 when exposed to cold or other stressors, but located within WAT depots, 22 especially in inguinal subcutaneous regions. 14 The presence of beige adipocytes in the WAT is due to a process called browning of WAT. 21 The ability of the BAT and beige adipocytes to dissipate energy has raised therapeutic expectations, currently the object of extensive studies in the context of obesity research.23,24
Interestingly, genetic studies have also demonstrated that white and brown adipocytes, in addition to having a dissimilar morphology and a complete distinct physiological role, have a divergent lineage. 25 They both originate from a common mesenchymal precursor cell, 26 but brown adipocytes share part of their developmental path with skeletal muscle cells. 25 The origin of the beige adipocyte is currently debated and it is the topic of intense research. 14
Discovery of the ECS in the Adipocyte
The presence of the CB1 receptor in adipocytes and the direct involvement of the ECS in adipocyte physiology were pinpointed for the first time by two independent groups in 200327,28 (see also Table 1). In the first work, we proposed the existence of a food intake-independent mechanism contributing to the leaner phenotype of mice with genetic deletion of CB1 (CB1-KO), suggesting that the reduced body fat of this animal model could result from both central and peripheral mechanisms. Endorsing this concept, we showed that CB1 was expressed in white adipocytes and that stimulation of CB1 in cultured primary mouse adipocytes from the WAT increased activity of the lipoprotein lipase (LPL), a key enzyme regulating triglyceride hydrolysis and fat storage. 27 In the second work, Bensaid et al. described a three- to fourfold increase of CB1 receptor mRNA expression in differentiated 3T3-F442A adipocytes and in the WAT of obese (fa/fa) rats compared to undifferentiated adipocytes and lean rats, respectively. 28 Protein quantification further confirmed the expression of the CB1 receptor in 3T3-F442A adipocytes, which was upregulated in differentiated cells, 28 suggesting a role for the ECS in the adipocyte differentiation process.
Qualitative and Quantitative Analysis of Endocannabinoid System Expression in White Adipocytes
For each year, authors are cited in alphabetical order.
AEA, anandamide; CM, confocal microscopy; DA, differentiated adipocytes; ELISA, enzyme-linked immunosorbent assay; FAAH, fatty acid amide hydrolase; IC, immunocytology; LC-MS, liquid chromatography–mass spectrometry; OEA, oleoylethanolamide; PEA, palmitoylethanolamide; RT-PCR, reverse transcriptase-polymerase chain reaction; Sc, subcutaneous; UDA, undifferentiated adipocytes; Vi, visceral; WB, western blot.
Since these initial findings, numerous studies have explored the existence of a functional ECS in adipose cells. In particular, a few investigations carried out in cultured rodent adipocytes confirmed not only the presence of CB1 but also identified the whole biochemical pathway for the EC synthesis and degradation.29,30 At this point, the possibility that the ECS was involved in the regulation of the adipocyte functions was becoming very intriguing. Several investigators started to explore the presence of the ECS also in human adipocytes, confirming the existence of a complete and functional ECS in both visceral and subcutaneous adipose cells.31–33 Accordingly, adipocytes produce measurable levels of the two main ECs, anandamide (AEA) and 2-arachidonoylglycerol (2-AG), 30 as well as EC-related compounds, such as palmitoylethanolamide (PEA) and oleoylethanolamide (OEA).34,35
Several studies then further explored the quantitative alterations in the expression of ECS constituents during adipocyte differentiation (see also the next chapter), providing consistent evidence of higher CB1 and FAAH (fatty acid amide hydrolase, the enzyme degrading AEA) mRNA levels in human isolated white adipocytes compared to undifferentiated cells.31,33 CB2 is also present in human adipocytes, but adipocyte differentiation does not consistently alter its expression.33,36
Besides, in the context of obesity, the study of the expression of CB receptors, EC levels, and related EC enzymes in the WAT has led to different results. While in the WAT from obese rodents CB1 mRNA 28 and protein 37 were found to be increased or not altered, 38 a decreased expression of CB1 mRNA was instead reported in the WAT from obese humans, a finding explained as the consequence of a negative feedback mechanism due to the increase of circulating ECs.31,39 Using radiolabeled substrates, a recent study has evaluated FAAH and MGL (monoacylglycerol lipase, the enzyme degrading 2-AG) activity in both subcutaneous and visceral adipocytes from both Zucker rats and obese humans, with or without diabetes. 40 The FAAH activity was increased in the adipocytes of obese Zucker rats, but not in the adipocytes of Zucker diabetic rats compared to Zucker lean rats, while the MGL activity was increased in both. 40 However, in obese humans, the FAAH and MGL activity in adipocytes was not altered by diabetes, dyslipidemia, or other markers of metabolic dysfunction. The authors therefore concluded that increased circulating EC levels are not the result of a decreased degradation in the adipose tissue, differently from what suggested by previous investigations.31,40 Of note, circulating EC levels are associated with body fat distribution. Obese subjects with increased intra-abdominal obesity have higher circulating 2-AG compared to subjects of similar body mass index (BMI) but with lower visceral fat. 41 Furthermore, a strong correlation exists between circulating 2-AG levels and high plasma triglyceride and low HDL-cholesterol in viscerally obese men. 42
Some studies have also described fat depot-specific alterations of CB receptors, ECs, and EC-related enzymes, associated with the different metabolic state of the subjects.36,43,44 Interestingly, caloric restriction together with exercise can alter the expression of these ECS components in obese humans, particularly at the level of the visceral abdominal fat depot. 43
Similar to what described earlier for white adipocytes, a functional ECS has been also found in mouse brown adipocytes. 45 Obesity, however, does not alter the expression of CB1 in BAT. 37 Interestingly, pharmacological acute stimulation of β3-adrenoceptors in murine BAT and brown adipocytes increases EC levels and the mRNA expression of CB1 and EC biosynthetic enzymes, implying that the ECS in brown adipocytes may be recruited to counteract β3 adrenoceptor-dependent BAT activation. 46
Role of the ECS in the Adipocyte
Role of the ECS in adipogenesis, lipogenesis, and lipolysis
Once demonstrated the existence of the ECS in white and brown adipocytes, several studies have investigated its functional role in these cells.
Adipogenesis is a well-known cell differentiation process by which preadipocytes become mature adipocytes under specific signals, including insulin, the hormone produced by the β-cells of the pancreas. The key features of differentiated white adipocytes are morphological change, increased expression of white adipocyte differentiation markers such as peroxisome proliferator-activated receptor γ (PPARγ), growth arrest, and thereby high expression of lipogenic genes to achieve energy storage. 47
Treatment of mouse 3T3-F442A adipocytes with the CB1 receptor agonist HU210 was found to increase the expression of PPARγ and the accumulation of lipid droplets. 30 All these effects were prevented by the coincubation of CB1 agonists with the CB1 antagonist rimonabant, pointing to the direct role of CB1 receptors in adipogenesis and in favoring lipid accumulation in adipocytes.27,30 AEA, however, is also known to act directly on PPARγ as a PPARγ agonist, causing adipocyte differentiation and lipid accumulation in both primary adipocyte culture and 3T3-L1 cells through a PPARγ-dependent mechanism.48,49 Similarly, treatment with the CB1 receptor agonist WIN-55,212 also increased proliferation of mouse adipocytes 3T3-L1, as demonstrated by the MTS-Formazan test, and the expression of the CB1 receptor, further supporting an involvement of the ECS in adipocyte proliferation and differentiation. 50 On the contrary, adipocyte differentiation stimulates by approximately twofold the binding efficiency of CB1 receptors, and by approximately twofold and approximately threefold, respectively, the catalytic efficiency of the AEA transporter and AEA hydrolase, suggesting that increased ECS activity may be required by fully differentiated adipocytes to work properly. 29
In turn, AEA can exert an antiadipogenic action when converted into prostaglandin F2α ethanolamide (PGF2αEA) in cells. 51 Thus, AEA stimulates adipogenesis through CB1 and PPARγ, while at the same time providing a potentially important termination mechanism for its proadipogenic effect. Of note, PGF2αEA levels are decreased in the WAT of mice fed a high-fat diet (HFD), likely because of the increased requirement for new adipocytes under this condition. 51
CB1 receptor expression and adipocyte differentiation are then directly regulated by the peroxisome proliferator-activated receptor δ (PPARδ), which is known to modulate PPARγ-stimulated adipocyte differentiation. 52 Silencing of PPARδ increased both CB1 receptor expression and 3T3-L1 adipocyte differentiation, whereas adenovirus-mediated overexpression of PPARδ significantly reduced both these events. 37
Data also suggest that the function of CB1 receptor in the regulation of adipocyte differentiation may be fat depot specific. Visceral adipocytes from CB1-KO mice showed decreased differentiation and increased apoptosis compared to wild-type (WT) controls. In contrast, subcutaneous adipocytes from CB1-KO mice had an accelerated differentiation and a reduced rate of apoptosis. 53 These effects, however, are also related to the diet, since when CB1-KO are fed an HFD, they have expression levels of adipocyte proliferation and differentiation markers similar to those observed in HFD-fed WT mice, with comparable activity of LPL. 54
The ECS is not only involved in white adipocyte proliferation and differentiation but plays also a role in regulating lipogenesis. Incubation of primary mouse adipocytes with the CB1 receptor agonist WIN-55,212 increased LPL activity in a CB1-dependent manner. 27 CB1 receptor activation can also stimulate lipogenesis by inhibiting cAMP formation. 30 In turn, cAMP inhibition stimulates lipogenesis by inhibiting the AMP-activated protein kinase (AMPK), which inhibits fatty acid synthesis by decreasing acetyl-CoA carboxylase (ACC) activity and malonyl-CoA availability. 55 Accordingly, Δ 9 -THC reduces AMPK activity in both the visceral and subcutaneous adipose tissue of mice, stimulating fatty acid synthesis particularly in the visceral fat. 56 Similarly, activation of CB1 receptors by peripheral administration of the CB1 agonist HU210 increased the expression of the lipogenic transcription factor SREBP-1c and of its targets, the lipogenic enzymes acetyl-CoA carboxylase 1 (ACC1) and fatty acid synthase (FAS), favoring fatty acid synthesis in the adipose tissue. 57 Stimulation of fatty acid synthesis and triglyceride biosynthesis, with increased expression of LPL, FAS, stearoyl-CoA desaturase 1 (SCD-1), and diacylglycerol acyltransferase 2 (DGAT-2), has been also found in 3T3-L1 adipocytes treated with the CB1 agonist WIN-55,212. 50
Conversely, when energy is needed, lipolysis occurs in the adipose tissue to liberate free fatty acids, which will undergo β-oxidation to generate ATP. 58 Chronic administration of the CB1 antagonist rimonabant to diet-induced obese mice has been shown to increase the expression of lipolytic enzymes, such as carnitine-acyl-CoA transferase (CAT), carnitine palmitoyltransferase 2 (CPT2), and crotonase in the WAT. 59 Rimonabant also induced the expression of β3-adrenergic and growth hormone receptors, known as essential lipolysis regulators. 59 However, the lipolytic action of rimonabant in vivo seems to be due to an induction of sympathetic activity rather than a direct effect on adipocytes. 60
Role of the ECS in glucose uptake and metabolism
In agreement with the stimulatory action on lipogenesis, the ECS also facilitates glucose entry in the adipocyte, a step required for the synthesis of fatty acids and triglyceride biosynthesis. AEA increases insulin-stimulated glucose uptake in 3T3-L1 differentiated adipocytes through a CB1-dependent mechanism. 29 In the same cell line, treatment with the CB1 antagonist AM251 leads to an increase in glucose transporter type 4 (GLUT4) protein and in its encoding gene Slc2a4. 61 Moreover, like insulin, activation of CB1 in human primary adipocytes promotes glucose uptake by favoring translocation of GLUT4 on the plasma membrane through increased [Ca2+]i and PI3-kinase activity.36,62 However, seemingly in contrast with this evidence, pharmacological blockade of CB1 receptors in diet-induced obese mice favors the expression of GLUT4 and glycolysis in adipocytes. In particular, in WAT, rimonabant was shown to increase the expression of the glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and other glycolytic enzymes such as the glycogen phosphorylase and synthase, the phosphofructokinase, and the β-enolase. 59 In this case, rimonabant was likely re-establishing obesity-compromised metabolic pathways and it was therefore suggested that these changes may have accounted at least, in part, for rimonabant-induced correction of insulin resistance. 59
Role of the ECS in mitochondrial activity
Chronic administration of rimonabant to diet-induced obese (DIO) has been also shown to induce the expression of enzymes involved in the tricarboxylic acid cycle, such as fumarase, aconitase, and oxoglutarate dehydrogenase. Interestingly, the expression of the carrier protein that exports ATP from mitochondria, the adenine nucleotide carrier (ANT), together with a component of the respiratory chain, the cytochrome C oxidase subunit VIa, were also upregulated by rimonabant. 59
Conversely, Tedesco et al. observed a decrease in mitochondrial biogenesis, mitochondrial DNA, and mitochondrial mass after CB1 stimulation by arachidonyl-2′-chloroethylamide hydrate (ACEA) in mouse and human primary white adipocytes.63,64 Key regulators of mitochondrial biogenesis such as the peroxisome proliferator-activated receptor gamma coactivator 1-α (PGC-1α), nuclear respiratory factor-1 (NRF-1), and transcription factor A (Tfam), as well as the endothelial nitric oxide synthase (eNOS), were found to be downregulated. Furthermore, these effects were blocked by nitric oxide donors, while ACEA was ineffective in adipocytes from eNOS-KO mice. Nevertheless, CB1 silencing did not totally prevent the inhibitory effect of ACEA on mitochondrial biogenesis and eNOS expression. Nevertheless, it appears that CB1 stimulation decreases mitochondrial biogenesis and eNOS expression through the p38 mitogen-activated protein kinase (MAPK) and AMPK phosphorylation, since ACEA increased p38 MAPK phosphorylation in treated adipocytes, while eNOS knockdown and p38 MAPK silencing both prevented ACEA effects. 64
Thus, through the modulation of both intracellular lipid and glucose metabolism and mitochondrial activity, the ECS can directly impact the physiology of the white adipocyte.
The ECS and the adipokines
The ECS can also affect the synthesis and release of several adipokines produced by white adipocytes and that are known to impact energy balance and metabolism.
Matias et al. together with Bellocchio et al. demonstrated that the activation of CB1 receptors by the agonist HU210 or WIN-55,212 inhibited expression of adiponectin, an adipokine known to have insulin sensitizing properties, 65 in 3T3-F442A or 3T3-L1 adipocytes, respectively.30,50 Conversely, Bensaid et al. showed that expression of adiponectin was enhanced both in WAT and in cultured mouse 3T3-F442A adipocytes by the CB1 antagonist rimonabant; an effect that was CB1 dependent. 28 Accordingly, later studies have shown that chronic rimonabant administration to obese mice and humans restores circulating adiponectin levels,66,67 an effect that seems important for the reversal of liver steatosis, but not for the reduction of adiposity and improved glycemic control induced by rimonabant in vivo. 68
Treatment of 3T3-L1 mouse adipocytes with WIN-55,212 also increased the mRNA expression of visfatin and apelin, two hormones involved in glucose metabolism, 65 whereas rimonabant inhibited their expression. 50 More recently, it has been demonstrated that in omental fat cultured explants from obese humans, rimonabant increased the expression of omentin, an anti-inflammatory adipokine secreted by the stromal vascular cells, 65 while decreasing the expression of proinflammatory cytokines. 69
Alternatively, adipokines can affect ECS function. In particular, leptin negatively regulates EC production in white adipocytes, either directly 30 or through its effects in the mediobasal hypothalamus. 70 Similar to leptin, also insulin decreases EC levels, in this case by stimulating FAAH expression. 71 However, these negative feedback mechanisms acting on the ECS in the adipocyte are lost in obesity, which is characterized by resistance to the actions of both leptin and insulin.70,71
Role of the ECS in the browning process of WAT
As briefly mentioned earlier, in response to appropriate stimuli as cold, exercise, and activation of β3-adrenoreptors, the WAT can undergo a process known as browning where it takes on characteristics of BAT, notably with the induction of UCP1 expression and the increased presence of multiple lipid droplets and mitochondria, leading to beige adipocytes. 21 Perwitz et al. demonstrated that the treatment of mouse immortalized white SV40 adipocytes with rimonabant increased the number of mitochondria and expression of UCP1 and of its coactivator PGC-1α in a time-dependent manner. 72 Furthermore, oxygen consumption of these cells was similar to the consumption observed in brown adipocytes, suggesting that inhibition of CB1 promoted transdifferentiation of white adipocytes into mitochondrial-rich, thermogenic brown-like adipocytes. 72 More recently, Krott et al. have shown that exposure to cold or acute administration of a β3-adrenoreptor agonist causes an upregulation of EC levels and biosynthetic enzymes in the WAT of mice, implying that ECS activity in WAT is part of an autocrine-negative feedback loop controlling β3-adrenoceptor-induced WAT browning. 46 The role played by CB1 in this phenomenon, however, seems to be fat depot specific. Indeed, subcutaneous, but not visceral, adipocytes from CB1-KO mice were found more sensitive toward a conversion into a beige phenotype, showing increased UCP1 and PGC-1α expression as well as augmented mitochondrial biogenesis and oxygen consumption compared to adipocytes from WT mice derived from the same type of fat depot. 53 In addition, relative balance in the actions of ECs and EC-related compounds might also regulate the browning process, as suggested by data obtained from mice lacking NAPE-PLD (N-acyl phosphatidylethanolamine phospholipase D, the enzyme responsible for the synthesis of AEA and OEA) in adipocytes. 73 These mutant mice develop obesity and metabolic alterations, which are associated with the downregulation of browning markers such as UCP1 in the WAT. 73 Accordingly, adipocyte NAPE-PLD deletion impairs adaptation to cold exposure. 73
Role of the ECS in the brown adipocytes
As briefly mentioned before, the sympathetic nervous system (SNS) plays a key role in the regulation of thermogenesis. Several studies published over the past few years suggest that the ECS regulates thermogenesis in the BAT by mainly inhibiting SNS activity, although a few investigations also point to a direct action of this system on the brown adipocyte.
Rimonabant-induced BAT thermogenesis seems in great part mediated by the SNS, since BAT denervation attenuates thermogenesis and UCP1 expression in the BAT.74,75 CB1 receptors expressed within the central nervous system (CNS) and peripheral sympathetic neurons are involved in these phenomena, as CB1 deletion in the forebrain and sympathetic neurons increases mitochondrial biogenesis and thermogenesis in the BAT. 76 Similarly, genetic overexpression of MGL in the forebrain, which would induce 2-AG hydrolysis, causes resistance to DIO through increased SNS-dependent BAT thermogenesis and mitochondrial activity. 77 These effects can be further localized to the hypothalamus, since we have demonstrated that mice with decreased expression of CB1 in this brain structure or lacking CB1 receptors in Single-minded 1 (Sim1) neurons, which constitute the majority of neurons of the hypothalamic paraventricular nucleus, have increased SNS activity and thermogenesis in the BAT under different experimental conditions.78,79 As mentioned earlier, β3-adrenoreptor activation by CL316,243 enhances EC levels in BAT and CB1 expression in primary brown adipocytes, 46 suggesting that the ECS is recruited in brown adipocytes to locally modulate the effects of sympathetic activity. Boon et al. have also demonstrated that rimonabant treatment of brown T37i adipocytes in vitro enhances UCP1 protein content and oxygen consumption rate. 80 These effects were still present in the BAT of mice treated with rimonabant and maintained at thermoneutrality or in mice treated with the peripherally restricted CB1 receptor antagonist AM6545, indicating direct peripheral activation of BAT. 80 Thus, although the majority of studies suggest an inhibitory effect of the ECS on thermogenesis due to the ability of the system to act on neural circuits and blunt SNS activity, some evidence exists of a direct action on the brown adipocyte.
Targeting the ECS in the Adipocyte for Obesity Treatment
The evidence reviewed above and summarized in Figure 1 clearly illustrates the contribution of the ECS to adipocyte physiology. However, whether this involvement may be relevant in vivo remains substantially unexplored.
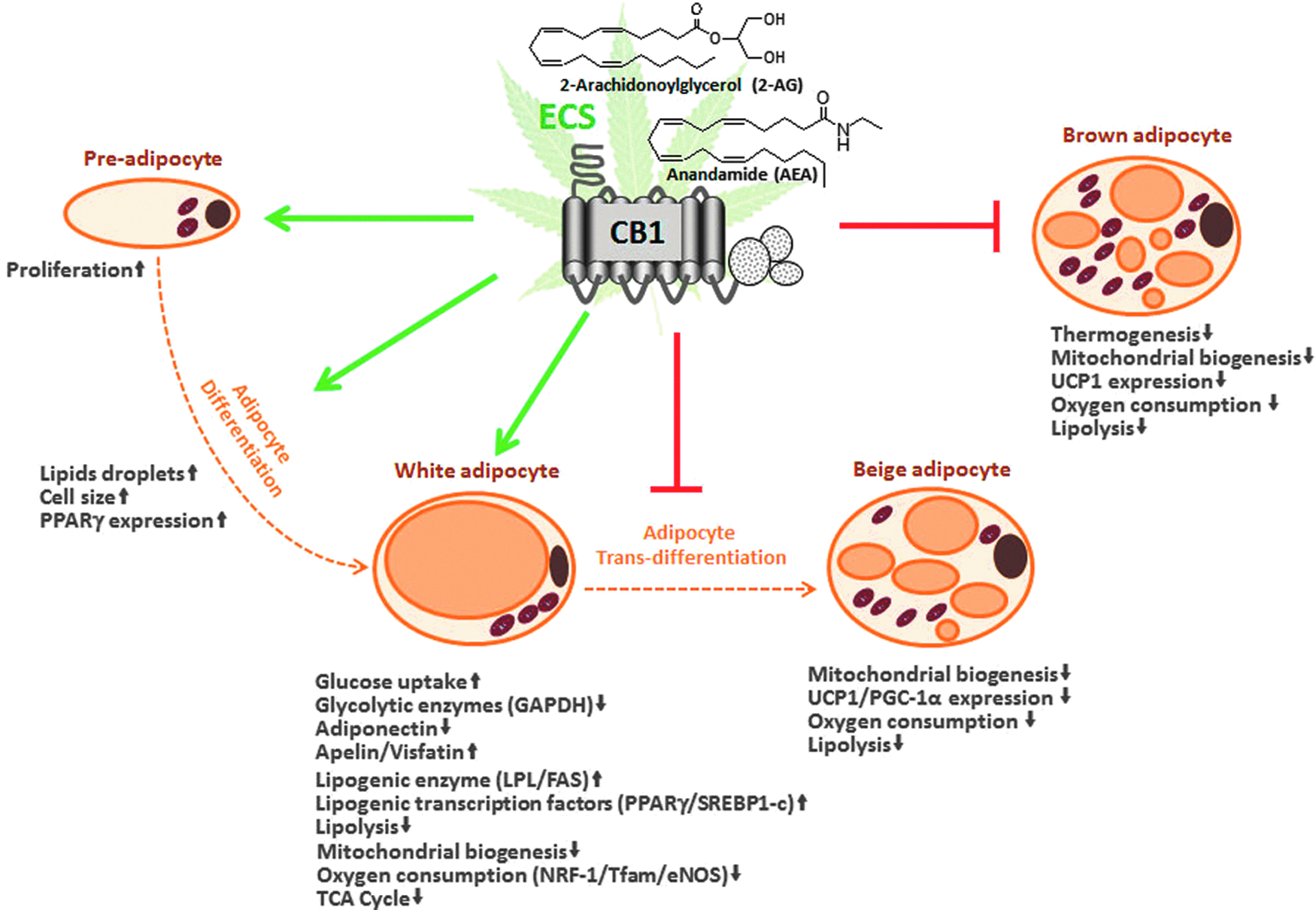
The ECS is implicated in very different processes affecting adipocyte physiology, spanning from stimulation of preadipocyte proliferation to determination of adipocyte phenotype. ECS activity in the white adipocyte stimulates glucose uptake and lipogenesis, while decreasing lipolysis, mitochondrial biogenesis, and expression of markers involved in white adipocyte transdifferentiation. Similarly, in the brown adipocyte, ECS activity decreases mitochondrial biogenesis and oxygen consumption, thus decreasing thermogenic capacity. Overall, ECS-driven processes in the adipocyte favor energy preservation and can therefore contribute to obesity. ECS, endocannabinoid system.
Preliminary data generated using adipocyte-specific CB1-KO mice suggest that adipocyte CB1 receptors regulate WAT expansion, in agreement with in vitro studies described in previous chapters, and directly impact the development of obesity and insulin resistance. 81 Indeed, adipocyte-specific CB1-KO mice are protected from these pathological conditions. 81 These findings therefore imply that therapies targeting CB1 receptors on adipocytes should beneficially impact metabolism. This conclusion is in agreement with recent data showing that blockade of CB1 receptors with JD5037, a peripherally restricted CB1 inverse agonist, decreased hyperleptinemia in diet-induced mice by inhibiting leptin expression and secretion from adipocytes via both prejunctional and postjunctional mechanisms. 82 This in turn seems to account for the decreased food intake caused by the administration of JD5037. 82
Thus, it is reasonable to propose that targeting adipocyte CB1 receptors in obesity might beneficially affect lipid fluxes and the production of adipokines (leptin, adiponectin) with effects that would facilitate body weight loss (i.e., decreased food intake) and improve lipid and glucose metabolism. Moreover, adipocyte CB1 receptors might be an interesting target for fat browning therapy, as suggested by both in vitro and ex vivo data reviewed in the previous chapters. The study of the browning process of the adipose tissue has attracted a lot of attention in recent years and represents a promising venue for the development of new antiobesity drugs. 83 As peripherally restricted CB1 antagonists or inverse agonists have been demonstrated to affect adipocyte function to decrease obesity and metabolic dysfunction, 82 their action on the browning process and consequently on thermogenesis should be further investigated. Alternatively, a possibility would be to target CB1 receptors specifically on the adipocytes, but this would require technological advances enabling selective drug delivery to specific cell types that at present are not yet available. Finally, another possibility is to modulate the levels of the ligands instead of directly targeting CB1 receptors. Specifically, nutritional interventions such as diet supplementation with n3 long chain polyunsaturated fatty acid (LCPUFA)-containing oils might represent a valid strategy aimed at reducing EC levels, particularly in peripheral organs. Indeed, a diet enriched with n3-LCPUFA reduced AEA and 2-AG levels in the visceral adipose tissue of Zucker rats, 84 while the same treatment decreased to a much lesser extent only 2-AG levels in the brain. 85 Interestingly, dietary n3-LCPUFA might modulate the levels of adipocyte phospholipids that act as EC precursors, since incubation of mouse 3T3-F442A adipocytes with docosahexaenoic acid (DHA) decreased 2-AG and AEA levels and the amounts of arachidonic acid esterified on both the sn-2 and sn-1 position of phospholipids. 86
In conclusion, the overview we have provided clearly illustrates the relevant roles played by the ECS in the adipocyte. Further investigation of these roles might result in the discovery of novel ECS mechanisms of action, which could lead to the identification of novel drug targets for obesity and metabolic disease.
Footnotes
Acknowledgments
The authors' work is supported by INSERM (Institut National de la Santé Et de la Recherche Médicale), Aquitaine Region, ANR (Agence National Recherche, Grant Numbers: ANR-13-BSV4-0006 to D.C., Labex BRAIN ANR-10-LABX-43 to D.C., equipex OPTOPATH ANR-10-EQX-008-1 to D.C.), and FFRD (Fondation Francophone Recherche Diabète, to D.C.).
Author Disclosure Statement
D.C. is among the founders of the start-up company Aelis Farma. She is also the stakeholder and consultant for the same company.