
Abstract
Background:
The endocannabinoid system is present in multiple organ systems and is involved in smooth muscle regulation, immune function, neuroendocrine modulation, and metabolism of tissues. Limited data are available regarding the presence and role of this system in reproductive tissues. Components of the endocannabinoid system have been identified in myometrial and placental tissues. However, no study has investigated differential expression of the endocannabinoid system in labor.
Objectives:
The purpose of this study was to identify and quantify two components of the endocannabinoid system, the CB1 cannabinoid receptor and cannabinoid receptor interacting protein 1a (CRIP1a) in uterine and placental tissues, and to determine if there is differential expression in tissues exposed to labor. We hypothesized that CB1 cannabinoid receptor concentration would be altered in uterine and placental tissue exposed to labor compared with tissues not exposed to labor.
Study Design:
Uterine and placental tissue samples were collected in nine laboring and 11 nonlaboring women undergoing cesarean delivery. CB1 cannabinoid receptor and CRIP1a presence and quantification were evaluated using western blot, immunohistochemistry, and real-time quantitative polymerase chain reaction. Statistical comparisons of laboring and nonlaboring subjects were made for uterine and placental tissue using a Mann–Whitney test.
Results:
Immunohistochemistry demonstrated positive staining for CB1 cannabinoid receptors and CRIP1a in uterine tissue. The protein abundance of CB1 cannabinoid receptor in uterine tissue was significantly lower in tissues exposed to labor (p=0.01). The protein abundance of CRIP1a was lower in uterine tissue exposed to labor but did not reach statistical significance (p=0.06). mRNA expression of CB1 cannabinoid receptor (p=0.20) and CRIP1a (p=0.63) did not differ in labored compared with nonlabored uterine tissues.
Conclusions:
Our findings of diminished protein density of CB1 cannabinoid receptor in uterine tissue exposed to labor support the hypothesis that the endocannabinoid system plays a role in parturition. Our data add to the growing body of evidence indicating the endocannabinoid system is of importance for successful reproduction and support the need for additional research investigating this complex system as it pertains to labor. ClinicalTrials.gov ID: NCT03752021.
Introduction
Multiple pathways leading to parturition have been proposed and investigated; a complete understanding of the signaling pathways leading to term and preterm labor has yet to be ascertained. The biology of labor is complex and includes interplay among steroid hormones, cytokines, and prostaglandins affecting the maternal–fetal interface and smooth muscle regulation.1–4
The most accepted theory of physiologic term labor in humans is that a functional progesterone withdrawal leads to an inflammatory response, causing a cascade to the final labor pathway. It is thought that preterm labor involves several different pathologic processes that lead to a similar cascade culminating in the final labor pathway. 5 However, decades of research targeting the inflammatory response and myometrial contractility have not resulted in effective therapy for preterm labor. Identification of a therapeutic target or marker that precedes this transition of myometrial quiescence to an active contractile state would allow innovative investigation of novel prevention or treatment strategies.
The endocannabinoid system (ECS) is present in multiple organ systems and is involved in smooth muscle regulation, immune function, neuroendocrine modulation, and metabolism of tissues.6–14 It includes CB1 and CB2 cannabinoid receptors, the endocannabinoid agonists anandamide and 2-arachidonoylglycerol (2-AG), and the enzymes that synthesize and metabolize the endocannabinoid ligands.14,15
The CB1 cannabinoid receptor is a G protein-coupled receptor with cell-specific activity and is encoded by the CNR1 gene. Activation leads to coupling predominantly with Gi/o proteins with effects on calcium channels, mitogen-activated protein kinases, and adenylyl cyclase.14–16 Cannabinoid receptor interacting protein 1a (CRIP1a) is a CB1 cannabinoid receptor-associated protein that is known to modulate CB1 cannabinoid receptor activity.17,18 The CB1 cannabinoid receptor specifically has been shown to influence myometrial contractility in vitro, 19 and has been associated with the onset of labor in a mouse model.20,21
The ECS is of importance in sustaining the microenvironment necessary for early pregnancy success and maintenance.22–27 It plays a significant role in embryo development, transport, and implantation as well as placentation.22–35 Limited data are available regarding the presence and role of this system in the mid- and late-trimesters. Components of the ECS have been identified in uterine and placental tissues.19,35,36 However, to our knowledge, no study has investigated differential expression of the ECS in labor.
The aims of this study were to identify and quantify CB1 cannabinoid receptor and CRIP1a in uterine and placental tissues and to determine if there is differential expression in those tissues exposed to labor. We hypothesized that CB1 cannabinoid receptor concentration would be altered in uterine and placental tissue exposed to labor as compared with tissues not exposed to labor.
Materials and Methods
This was an observational study of women undergoing cesarean section. It was conducted after Institutional Review Board approval at Wake Forest Baptist Medical Center (Winston-Salem, NC) and Novant Forsyth Medical Center (Winston-Salem, NC). Written informed consent was obtained from each participant. Pregnant women with singleton gestations between 22 weeks, 0 days through 42 weeks, 0 days undergoing cesarean section were eligible to participate. Exclusion criteria included cannabinoid use during pregnancy, illicit drug use during pregnancy, nonsteroidal anti-inflammatory drug use within 7 days of delivery, pre-existing diabetes, pre-existing hypertension, hypertensive disorders of pregnancy, epilepsy currently treated with antiepileptic medication, intraamniotic infection, and fetal anomalies.
Baseline demographics were collected from each subject's medical record. Labor was defined as cervical change with regular contractions. Uterine and placental samples were collected at the time of cesarean section by a single surgeon. A 2×0.5-inch uterine sample was obtained from the superior edge of the lower uterine segment incision following delivery of the placenta. The placental samples were taken medial to the placental edge and did not include the cord insertion site. For the placental sample, the fetal surface was removed and the tissue was cut into four or five pieces (1 cm2). All tissue samples were thoroughly rinsed in sterile normal saline, placed in RNase-free sterile containers, flash frozen in the operating room, and stored at −80°C immediately after delivery.
Western blotting
Tissue specimens (150 mg) were pulverized and then homogenized with a Dremel 300 tissue homogenizer in 1.5 mL radioimmunoprecipitation assay buffer (Thermo Fisher Scientific, Waltham, MA) containing proteinase inhibitor cocktail (Thermo Fisher Scientific). Samples were then centrifuged at 1,000×g for 10 min at 4°C. The supernatant was centrifuged at 25,000×g for 20 min at 4°C. Protein concentrations were measured using a bicinchoninic acid assay. 37 Samples were prepared in the loading buffer at 2 μg/μL and heated at 65°C for 8 min. Samples (60 μg) were loaded onto a 4–20% Novex wedgewell gel (Thermo Fisher Scientific) and electrophoresed at 60 V for 5 min followed by 100 V for 90 min, and transferred to a polyvinylidene fluoride membrane (Millipore, Burlington, MA).
Blots were allowed to dry completely and were stored in the dark at room temperature until antibody incubation. Blots were rewetted in methanol for 15 sec, rinsed for 1 min in phosphate-buffered saline (PBS), and blocked for 60 min with Odyssey blocking buffer (LI-COR Biosciences, Lincoln, NE) at 21–23°C. Blots were probed with primary antibodies (4°C for 18 h) for CB1 cannabinoid receptor (catalog no. EB10961; Everest Biotech, Oxfordshire, United Kingdom), CRIP1a (catalog no. sc-515504; Santa Cruz Biotechnology, Santa Cruz, CA), and β-actin (catalog no. 66009; Proteintech, Rosemont, IL). Blots were washed and incubated with an appropriate Li-COR IR dye-conjugated secondary antibody.
Bands were imaged using the Odyssey CLx Infrared Imaging System and quantified by densitometry using Image Studio software (LI-COR Biosciences). Band densities were normalized to β-actin as a loading control and then normalized to the mean of all samples being assigned a value of 1.0.
Quantitative real-time polymerase chain reaction
RNA was isolated using the RNeasy mini kit (Qiagen, Santa Clarita, CA). RNA yield and purity were determined spectrophometrically using a NanoDrop (Thermo Fisher Scientific). The RNA was reverse transcribed to cDNA using a high-capacity cDNA kit (catalog no. 4368814; Thermo Fisher Scientific).
Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and hypoxanthine phosphoribosyltransferase 1 (HPRT1) were first evaluated for variability between sample groups. Because their expression did not differ between labored and nonlabored samples in uterine or placental tissues, these genes were used for normalization in the present studies.
The primers used were as follows: CNR1 5′-AGCAGACCAGGTGAACATTAC-3′ and 3′-GACCATGAAACACTCTATG-5′; CRIP1a 5′-CCAGTTGTTCTCGGTCATACTT-3′ and 3′-AAAGAGCGGAGCTGTTTATAGG-5′; GAPDH 5′-ACATCGCTCAGACACCATG-3′ and 3′-TGTAGTTGAGGTCAATGAAGGG-5′; HPRT1 5′-TTGTTGTAGGATATGCCCTTGA-3′ and 3′-GCGATGTCAATAGGACTCCAG-5′ (Integrated DNA Technologies, Coralville, IA). cDNA was diluted to 5 ng/mL. Quantitative real-time polymerase chain reaction (qPCR) was performed using SYBR Green Master Mix (catalog no. QP001-01; GeneCopoeia, Rockville, MD) in triplicate with the Applied Biosystems StepOne Real-Time PCR System (Applied Biosystems, Foster City, CA). The comparative threshold cycle method was used to calculate relative gene expression. 38
Immunohistochemistry
Tissue was embedded in optimal cutting temperature compound and sectioned (5 μm) using a Leica Cryostat (Leica Biosystems, Buffalo Grove, IL). Tissue was fixed with cold (−20°C) methanol for 5 min at room temperature. Sections were washed in PBS and incubated in 3% hydrogen peroxide at room temperature for 5 min to block endogenous peroxidase. Sections were washed, and nonspecific background staining was blocked by normal horse serum (Vector Laboratories, Burlingame, CA) or normal goat serum (Vector Laboratories).
Tissue sections were incubated with primary antibody for CB1 cannabinoid receptor (1:100) (catalog no. EB10961; Everest Biotech), CRIP1a (1:100) (catalog no. sc-515504; Santa Cruz Biotechnology), and alpha-smooth muscle Actin (1:50) (catalog no. ab5694; Abcam, Cambridge, United Kingdom) overnight at 4°C. Negative controls were incubated with normal goat, mouse, or rabbit serum rather than primary antibody. Sections were washed and incubated with biotinylated rabbit anti-goat, goat anti-rabbit, or goat anti-mouse secondary antibodies (1:500) (Vector Laboratories).
Sections were washed and incubated with avidin–biotin complex (Vector Laboratories) for 30 min at room temperature. Sections were incubated with 3,3′-diaminobenzidine until brown staining was visible (1 min). Sections were counterstained with hematoxylin. The slides were dehydrated in a graded alcohol series, cleared in xylene, and mounted in permanent mounting medium. Sections were imaged with an Axioplan 2 upright microscope with attached camera (Zeiss, Oberkochen, Germany). Sheep hippocampus was used as a positive control.
For the fluorescent staining, tissue sections were fixed with cold (−20°C) methanol for 5 min at room temperature and then washed in PBS. Nonspecific background staining was blocked by normal horse serum (Vector Laboratories). Tissue sections were incubated with primary antibody for CB1 cannabinoid receptor (1:100) (catalog no. EB10961; Everest Biotech) overnight at 4°C. Negative controls were incubated with normal goat serum rather than primary antibody. Sections were washed and incubated with fluorescent donkey anti-goat secondary antibody (1:500) (catalog no. A11056; Thermo Fisher Scientific) for an hour at room temperature. Sections were washed, then dehydrated in a graded alcohol series, cleared in xylene, and mounted in permanent mounting medium. Sections were imaged with an Axioplan 2 upright microscope with attached camera (Zeiss).
Statistical analysis
Statistical comparisons for uterine and placental tissue were made using a Mann–Whitney test, looking at differences in receptor concentration among tissues exposed to labor compared with those not exposed to labor. Significant difference was considered to be at p<0.05. All analyses were performed using GraphPad Prism version 8.3.0 for Windows (GraphPad Software, San Diego, CA). Data are presented as mean±standard error of the mean. Sample size calculation was not performed as there was no published literature to determine what effect size would be significant.
Results
Twenty healthy subjects were enrolled in the study. The median gestational age was 38 weeks, 5 days in the labor group (n=9) and 39 weeks, 0 days in the nonlabor group (n=11). Additional characteristics are listed in Table 1. There was no significant difference in age, body mass index, race, gestational age, tobacco use, or birth weight. There was a significant difference in parity with the labor group having more nulliparous subjects (p=0.02). Cesarean deliveries were performed in the nonlabor group for history of cesarean section with desire for repeat cesarean, previous shoulder dystocia, breech presentation, and fetal growth restriction with non-reassuring antenatal testing. Cesarean deliveries were performed in the labor group for non-reassuring fetal heart rate tracing, history of cesarean section with desire for repeat, and arrest of descent. All subjects received either epidural or spinal anesthesia.
Subject and Delivery Characteristics
Statistical comparisons were made using an unpaired Student's t-test or Fisher exact test as appropriate with significant difference (*) considered to be p<0.05.
Uterus
CB1 cannabinoid receptor and CRIP1a were both identified in uterine samples. Immunohistochemistry demonstrated positive staining for CB1 cannabinoid receptors and CRIP1a in uterine tissue (Fig. 1). Immunostaining for alpha-smooth muscle actin (B) highlights the cytoplasm of the myofibers within the smooth muscle bundle, and the CB1 cannabinoid receptor staining correlated with the cell membranes of both the myocytes and vessel wall (A). Higher magnification of CB1 cannabinoid receptor staining (C) better demonstrates membrane localization. CRIP1a, localized at the cell membrane, has a more nonuniform distribution (D) and may also be visualized intracellularly. With fluorescent staining, CB1 cannabinoid receptor appears qualitatively less abundant in the smooth muscle bundles of labored uterine tissue (E) compared with nonlabored uterine tissue (F).

Immunostaining of CB1 cannabinoid receptor and CRIP1a in uterine tissue. Representative images from four or more sections of uterine tissue taken from laboring and nonlaboring subjects (n=4). Immunostaining specific for CB1 cannabinoid receptor
Quantitative evaluation using western blot indicated the protein abundance of CB1 cannabinoid receptor in uterine tissues was significantly lower in laboring subjects compared with nonlaboring subjects (Figs. 2 and 3). Band intensity of CB1 cannabinoid receptor normalized to β-actin was 0.79±0.118 (n=9) for all laboring subjects and 1.19±0.093 (n=11) for all nonlaboring subjects (p=0.01). Relative band intensity of CRIP1a was 0.76±0.125 (n=9) for all laboring subjects and 1.17±0.126 (n=11) for all nonlaboring subjects (p=0.06). Subgroup analysis of term laboring subjects (n=6) compared with term nonlaboring subjects (n=9) identified less CB1 cannabinoid receptor protein in tissue exposed to labor compared with tissue not exposed to labor (p=0.02).

Western blot band intensities of CB1 cannabinoid receptor and CRIP in laboring and nonlaboring uterus and placenta.
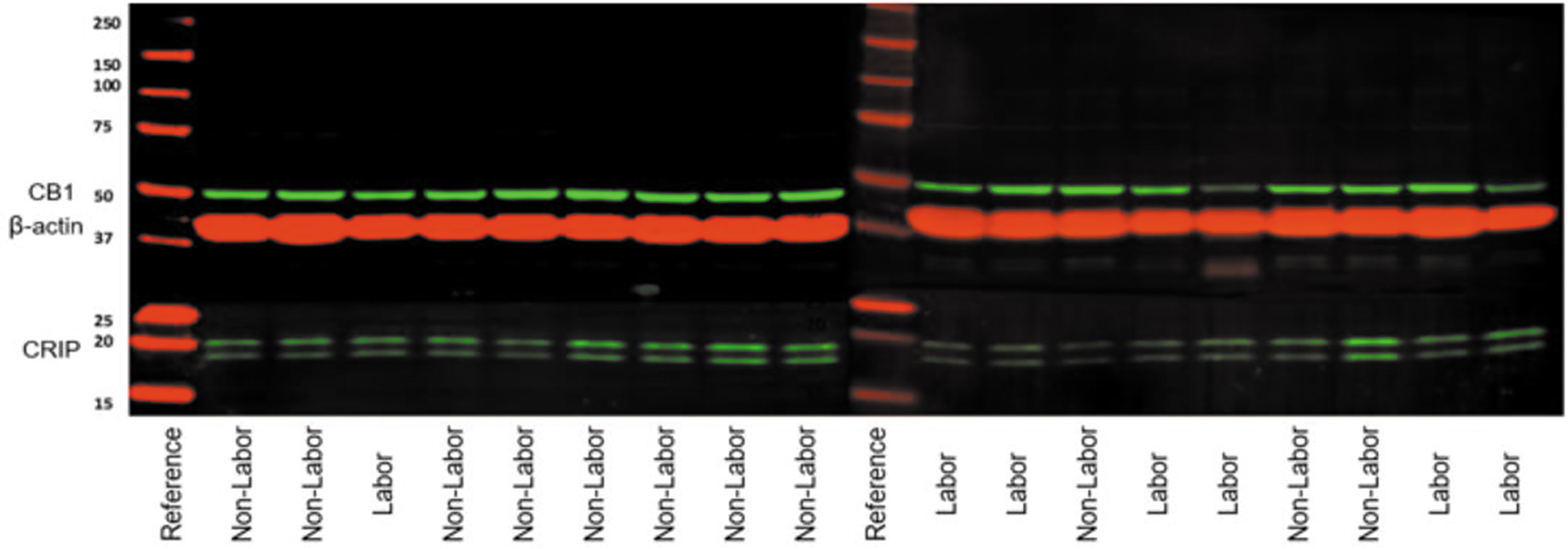
Western blot images. CB1 and CRIP1a immunoblots were visualized using the LI-COR Odyssey system. Bands were normalized to β-actin loading control.
CRIP1a protein abundance was also significantly lower in labored uterine tissue in the term subgroup (p=0.03). Although there were significantly more nulliparous subjects in the laboring group, there was no significant difference in the CB1 cannabinoid receptor (p=0.17) or CRIP1a (p=0.38) protein density between nulliparous and multiparous laboring subjects in uterine tissue. Additional subgroup analyses showed no significant difference between all term (n=15) and preterm (n=5) subjects for CB1 cannabinoid receptor (p=0.24) or CRIP1a (p=0.74), and no significant difference between subjects in spontaneous labor (n=7) and induced labor (n=2) for CB1 cannabinoid receptor (p=0.11) or CRIP1a (p=0.50).
There was no significant difference in mRNA expression of CB1 cannabinoid receptor (p=0.20) or CRIP1a (p=0.63) in labored compared with nonlabored uterine tissues (Fig. 4). Subgroup analyses including only term subjects and term subjects in spontaneous labor also showed no significant difference in mRNA expression of CB1 cannabinoid receptor or CRIP1a.

mRNA expression of CB1 cannabinoid receptor and CRIP1a using quantitative real-time polymerase chain reaction did not differ in uterine or placental tissues exposed to labor.
Placenta
CB1 cannabinoid receptor and CRIP1a were both identified in human placental samples. There was no significant difference in band density of the western blots when comparing the labor and non-labor groups (Fig. 2). Band intensity of CB1 cannabinoid receptor normalized to β-actin was 1.13±0.306 (n=9) for all laboring patients and 0.89±0.035 (n=11) for all nonlaboring patients (p=0.40). Relative band intensity of CRIP1a was 1.56±0.549 (n=9) for all laboring patients and 0.55±0.083 (n=11) for all nonlaboring patients (p=0.06). No significant difference in CB1 cannabinoid receptor (p=0.36) or CRIP1a (p=0.30) mRNA expression was identified between the two groups (Fig. 4).
Discussion
Our data confirm that the components of the ECS are present in human uterine and placental tissues by using western blot, qPCR, and immunohistochemistry. The most prominent finding was that CB1 cannabinoid receptor levels are significantly diminished in uterine tissue exposed to labor. However, no significant difference was identified between labored and nonlabored placental tissue. These findings add to the evidence that the ECS is involved in uterine and placental function.
Previous studies have demonstrated cannabinoid receptor presence in myometrial and placental tissues.19,35,36 A study evaluating myometrial cells obtained from segments of term human myometrial tissue identified CB1 and CB2 cannabinoid receptors. 19 In the same study, the effects of cannabinoids on uterine contractility were evaluated in vitro, and a CB1 cannabinoid receptor-mediated relaxant effect of endogenous and exogenous cannabinoids on myometrial contractility was demonstrated. 19
In addition, a mouse model has previously been used to evaluate the effect of CB1 cannabinoid receptor inactivation on parturition. 20 Mice lacking the CB1 cannabinoid receptor were found to experience earlier onset of labor compared with wild-type mice. 20 In wild-type mice in the same study, CB1 cannabinoid receptor silencing in late gestation also resulted in labor. 20 Our findings of reduced CB1 cannabinoid receptor protein in laboring uterine tissue are concordant with these previously published findings.
Despite decades of research, preterm labor remains a significant cause of maternal–fetal morbidity and economic strain.39,40 Complications related to premature birth include neurodevelopmental disorders, retinopathy of prematurity, bronchopulmonary dysplasia, and hearing impairment among others. Physiologic and pathologic mechanisms leading to the initiation of the final labor pathway have been implicated in the etiology of term and preterm birth, but a clear understanding of the pathophysiology has not been ascertained.1–5,39–44
The major challenge in the development of treatment and preventative therapies is the lack of a clear understanding of the processes responsible for shifting the myometrium from quiescence to an active contractile state. Identifying the ECS as a system that contributes to events preceding the labor cascade may open the door to therapeutic modalities not previously studied and would help to better understand the physiology of labor.
Current treatment of preterm labor targets stimulation of fetal maturity with corticosteroids, magnesium sulfate for fetal neuroprotection, and the consideration of tocolytic medications targeting uterine contractility. 39 Various classes of medications to decrease uterine contractions have been studied including calcium channel blockers, selective beta 2-adregergic agonists, prostaglandin synthase or cyclooxygenase (COX) inhibitors, and oxytocin receptor antagonists.41–44 Despite their frequent use, no tocolytic therapy has been shown to independently improve neonatal outcomes. First-line tocolytic medications currently recommended include nifedipine (calcium channel blocker) and indomethacin (nonselective COX inhibitor). 39 Their short-term use is indicated to allow for the administration of corticosteroids, magnesium sulfate for fetal neuroprotection and for transfer to a tertiary care center with neonatology capabilities.
Indomethacin has been studied as a tocolytic therapy since the 1970s after it was noted to inhibit prostaglandin activity. 44 Of interest, a recent study has identified indomethacin as being a positive allosteric modulator of the CB1 cannabinoid receptor. 45 Modulation of CB1 cannabinoid receptor signaling pathways has been a target of investigation with significant overlap of the prostaglandin and endocannabinoid pathways noted.45,46
Our data demonstrating lower levels of CB1 cannabinoid receptor in labored uterine tissue suggest that downregulation of the ECS during labor could be important for the mechanisms allowing labor to proceed. Allosteric modulators of the CB1 cannabinoid receptor are being investigated as possible therapy for various pathologic disorders in nonreproductive organ systems.46–48 Further investigation of ECS-related pathways may elucidate targets for clinical research pertaining to parturition.
Our findings have significant research implications; specifically, they warrant further investigation of the ECS as a system intimately involved in the labor process. One must contemplate if this system contributes to the maintenance of uterine quiescence. Although we have determined there is a significant decrease in CB1 cannabinoid receptor in laboring uterine tissue, there are other components of the ECS still to be studied in this population.
The endogenous lipid ligands anandamide and 2-AG bind to the CB1 and CB2 cannabinoid receptors. 14 Plasma anandamide levels have been correlated with risk of spontaneous pregnancy loss in the first trimester.30–32 Both anandamide and 2-AG have been identified as being of critical importance in early pregnancy events.22–32 Furthermore, plasma levels of anandamide have been shown to increase at term and dramatically increase in labor.49,50 Recent research has included the endocannabinoids, their precursors and their pathways of degradation as targets for prediction of preterm labor with a primary finding of anandamide being predictive of preterm birth. 51
Anandamide is primarily made from N-arachidonyl phosphatidylethanolamine (NAPE) by NAPE-hydrolyzing phospholipase D, although other pathways can also contribute to its synthesis.9,10 It is primarily degraded by the enzyme fatty acid amide hydrolase (FAAH), although it can be metabolized by other enzymes. 52 The activity of FAAH has been shown to be an important factor in early pregnancy success.32,35 It has been shown that low levels of anandamide correlate with high levels of FAAH at uterine implantation sites suggesting this is an important factor for successful pregnancy implantation. 22
FAAH protein expression as it relates to CB1 cannabinoid receptor expression has been described using immunohistochemistry in term membranes and placentas. 36 Furthermore, mice without FAAH and thus with chronically high levels of anandamide have been shown to be more susceptible to inflammation-induced preterm labor regardless of progesterone levels. 21
Whereas anandamide predominantly binds to the CB1 cannabinoid receptor, there is evidence that it can also act on other receptors including the ligand-gated transient receptor potential vanilloid receptor type 1 (TRPV1) 53 and peroxisome proliferator-activated receptors (PPARs).54,55 The presence of TRPV1 channels in nongravid myometrial cells has been demonstrated using calcium imaging. 56
The action of anandamide on the TRPV1 receptor is tissue specific and is influenced by a variety of factors. 57 When anandamide engages this receptor in other tissue types, it results in channel activation.58,59 However, in nongravid human myometrial cells, this was not observed. 56 PPARs have been identified in human uterine and placental tissues and a role for them in reproduction has been investigated.60–62 Anandamide's action on PPARs in reproductive tissues has not clearly been elucidated.
In our study, whereas the protein levels of CB1 cannabinoid receptor differed in the uterine tissue exposed to labor compared with that not exposed to labor, mRNA expression did not vary by qPCR. Investigation of the mechanisms explaining the variation in CB1 cannabinoid receptor and CRIP1a protein abundance without a change in mRNA expression is warranted. Mechanisms of interest include receptor desensitization and protein degradation.
To our knowledge, this study is the first to quantify the CB1 cannabinoid receptor and CRIP1a in laboring uterine and placental tissues. It included a fairly homogenous population of women without significant comorbidities, and all samples were collected consistently by a single surgeon. A limitation of this study is that the laboring group did include a small number of patients being induced or augmented with oxytocin. However, the vast majority were spontaneously laboring patients. Also, the underlying factors necessitating cesarean section as the mode of delivery could have impacted our results.
Conclusions
There exists significant overlap between prostaglandins, cytokines, steroid hormones, and the ECS. A growing body of evidence indicates that appropriate interaction between these pathways is important for successful reproduction and normal labor. Our data underscore the need for additional research investigating this complex system as it pertains to labor.
Footnotes
Acknowledgments
The authors thank Christina Tulbert and Jorge Figueroa for their assistance in acquisition of equipment for the study.
Disclaimer
The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by National Institute on Drug Abuse (NIDA) grant R01-DA042157 and the National Center for Advancing Translational Sciences (NCATS), National Institutes of Health, through grant UL1-TR001420.