
Abstract
Introduction:
Cannabidiol (CBD) is a major cannabinoid extracted from Cannabis sativa with no abuse potential. Data from recent rodent studies suggest that amelioration of alcohol-motivated behaviors may be one of the numerous pharmacological effects of CBD. This study was designed to contribute to this research, assessing the effect of CBD on operant oral alcohol self-administration in selectively bred Sardinian alcohol-preferring (sP) rats, a validated animal model of excessive alcohol consumption. In addition, this study investigated the effect of CBD on operant self-administration of a highly palatable chocolate solution in Wistar rats.
Materials and Methods:
Male sP rats were trained to lever respond for alcohol (15% v/v) under the fixed ratio 4 (FR4) schedule of reinforcement. Once lever responding had stabilized, rats were exposed to test sessions under the FR4 and progressive ratio (PR) schedules of reinforcement. Test sessions were preceded by acute treatment with CBD (0, 6.25, 12.5, and 25 mg/kg or 0, 25, 50, and 100 mg/kg, i.p.; each dose range was tested in an independent experiment). Male Wistar rats were trained to lever respond for a chocolate solution (5% w/v chocolate powder) under the FR10 schedule of reinforcement. Once lever responding had stabilized, rats were exposed to test sessions under the same schedule. Test sessions were preceded by acute treatment with CBD (0, 6.25, 12.5, and 25 mg/kg or 0, 25, 50, and 100 mg/kg, i.p., in two independent experiments).
Results:
Under the FR schedule, treatment with doses of CBD ≥12.5 mg/kg markedly reduced lever responding for alcohol and amount of self-administered alcohol. Under the PR schedule, treatment with CBD produced a slight tendency toward a decrease in lever responding and breakpoint for alcohol. Finally, no dose of CBD affected lever responding for the chocolate solution and amount of self-administered chocolate solution.
Discussion:
These results extend previous data on CBD ability to affect alcohol-motivated behaviors to an animal model of genetically-determined proclivity to high alcohol consumption. Because of the predictive validity of sP rats, these results may be of relevance in view of possible future studies testing CBD in patients affected by alcohol use disorder.
Introduction
The role of the endocannabinoid (eCB) neurotransmitter system in the regulation of several alcohol-related behaviors is well documented and summarizable with cannabinoid CB1 receptor antagonists/inverse agonists inhibiting and cannabinoid CB1 receptor agonists stimulating alcohol drinking, operant alcohol self-administration, and reinstatement of alcohol seeking in rats and mice.1–4 The adverse neuropsychiatric effects of the prototypic cannabinoid CB1 receptor antagonist/inverse agonist, rimonabant, have regrettably prevented a proper translation to patients affected by alcohol use disorder (AUD) of the large number of remarkably consistent rodent studies reporting its “antialcohol” properties.
A role for the cannabinoid CB2 receptor has also been proposed, with the cannabinoid CB2 receptor agonist, JWH015, increasing alcohol drinking in mice. 5
Over recent years, research in the eCB pharmacology field has been enriched by an increasing interest in cannabidiol (CBD), a major cannabinoid extracted from Cannabis sativa with no abuse potential.6,7 CBD acts as inverse agonist 8 and negative allosteric modulator 9 at both CB1 and CB2 cannabinoid receptors. CBD also affects the eCB system by blocking uptake of the endogenous cannabinoid receptor ligand, anandamide, and inhibiting its enzymatic hydrolysis. 10
In the alcohol research field, to date, CBD has been tested in a limited number of preclinical studies.11,12 Specifically, acute and/or repeated treatment with CBD attenuated (i) context- and stress-induced reinstatement of alcohol seeking in rats, 13 (ii) alcohol drinking in mice exposed to the “alcohol versus water” choice regimen, 14 (iii) binge-like drinking in mice exposed to the “drinking-in-the-dark” procedure, 15 and (iv) operant oral alcohol self-administration in mice.14,16 It is therefore of interest to test CBD in additional animal and experimental models of AUD, with the intent of better characterizing its “antialcohol” potential before possibly moving to clinical studies.
To this end, this study was designed to investigate the effect of treatment with CBD on operant oral alcohol self-administration in selectively bred Sardinian alcohol-preferring (sP) rats. 17 Rats of the sP line appear to constitute a proper animal model for this investigation, as—in this rat line—(i) genetically determined differences in the eCB system contribute to high alcohol preference and consumption 18 ; (ii) treatment with rimonabant reversed alcohol-induced dysfunctions of eCB 18 ; and (iii) alcohol self-administration proved highly sensitive to pharmacological manipulation of the CB1 receptor, with rimonabant treatment effectively suppressing lever responding for alcohol. 19
As an additional aim, this study investigated the effect of CBD on operant self-administration of a chocolate solution in Wistar rats. Previous studies reported contrasting results on the ability of CBD to affect sucrose intake and self-administration in rodents,20–22 leaving open the question as to whether CBD may modulate the appetitive and consummatory aspects of palatable food. The relevance of this study may reside in the experimental model: self-administration of a chocolate solution possessing addictive-like properties and inducing exaggerated lever responding in rats. 23
Materials and Methods
The experimental procedures employed in this study fully complied with European Directive No. 2010/63/EU and subsequent Italian Legislative Decree No. 26, March 4, 2014, on the “Protection of animals used for scientific purposes” and have been approved by the Italian Ministry of Health (authorization no. 342/2016-PR of April 1, 2016, and the subsequent integration of February 22, 2018).
Animals
Male sP (bred in our laboratory) and Wistar (Envigo, San Pietro al Natisone, Italy) rats were used. Rats were 55 days old at the start of each experiment. Rats of the sP line were from the 101st and 108th generations and alcohol naive at the start of the study. Rats were housed 3 per cage in standard plastic cages with wood chip bedding. The animal facility was under an inverted 12:12-h light/dark cycle (lights on at 5:00 p.m.), at a constant temperature of 22°C±2°C and relative humidity of ∼60%. Standard rat chow (Envigo, San Pietro al Natisone, Italy) was always available in the homecage. Tap water was also always available, except as noted below. Rats were extensively habituated to handling and intraperitoneal injections.
Apparatus
Self-administration sessions were conducted in standard, modular chambers (Med Associates, St. Albans, VT) described in detail elsewhere. 24 Chambers used in “alcohol” experiments were equipped with two retractable response levers, connected to two syringe pumps located outside the chamber. For half of the rats, right and left levers were associated with alcohol and water, respectively; the opposite condition was applied to the other half of the rats. Chambers of “chocolate” experiments were equipped with one retractable response lever and the retractable spout of a liquid sipper bottle located outside the chamber.
In the alcohol self-administration sessions, achievement of the response requirement (RR) resulted in activation of alcohol or water pumps, delivery of 0.1 mL fluid, and illumination of the stimulus light. In the chocolate self-administration sessions, achievement of RR resulted in 5-s exposure of the sipper spout of the chocolate-containing bottle and illumination of the stimulus light.
Experimental procedures
Alcohol self-administration in sP rats
Training and maintenance phases
Rats were initially exposed to the homecage 2-bottle “alcohol (10% v/v) versus water” choice regimen with unlimited access for 24 h/day over 10 consecutive days. This initial phase was (i) part of the conventional procedure of alcohol self-administration employed in our laboratory with sP rats, and (ii) conducted to accustom rats to the taste of alcohol and to experiencing its psychopharmacological effects with the aim of potentially shortening the subsequent autoshaping phase in the operant chambers. 24
Subsequently, rats were trained to lever respond for alcohol. Daily self-administration sessions lasted 30 min (with the sole exception of the very first session, which lasted 120 min). Rats were water deprived exclusively during the 12 h before the first session in the operant chamber. Rats were initially exposed to a fixed ratio 1 (FR1) schedule of reinforcement for 10% alcohol (v/v) for 4 daily sessions. FR was then increased to FR2 and FR4 over 4 sessions. In sessions 9 and 10, the alcohol solution was presented at a final concentration of 15% (v/v). Rats were then exposed to four sessions during which the water lever alone or alcohol lever alone was available every other day; water and alcohol were available on FR1 and FR4, respectively.
From then onward, both levers were concomitantly available (maintenance phase) for a total of 20 sessions conducted with FR4 and FR1 on the alcohol and water lever, respectively. On completion of these 20 self-administration sessions, rats displaying the most stable responding behavior were selected for use in CBD experiments.
Testing under the FR schedule
Two different experiments were performed, each testing a different CBD dose range and using independent sets of rats. On the test days, rats of each set were divided into four subgroups of n=10–12, and treated acutely with either 0, 6.25, 12.5, and 25 mg/kg or 0, 25, 50, and 100 mg/kg CBD. The experiment testing the higher dose range was designed once data from the experiment testing the lower dose range were available, with the intent of clarifying whether doses of CBD >25 mg/kg would produce a reduction in alcohol self-administration of magnitude larger than that observed when testing the lower dose range.
In both experiments, CBD (THC Pharm, Frankfurt am Main, Germany) was suspended in a 5:5:7:83 mixture of DMSO, 1,2-propanediol, Cremophor EL, and saline and administered intraperitoneally (injection volume: 4 mL/kg) 30 min before the start of the test session. Test sessions were conducted immediately after completion of the maintenance phase and were identical to those of the maintenance phase.
Measured variables were number of responses on each lever and amount of self-administered alcohol (expressed in g/kg pure alcohol). The latter was estimated from the number of earned reinforcers (assuming that each reinforcer was entirely consumed). Data were statistically evaluated by one-way ANOVA, followed by Tukey's test for post hoc comparisons, or Kruskal–Wallis test.
Testing under the progressive ratio schedule
Two different experiments were performed, each testing a different CBD dose range and using independent sets of rats. On the test days, rats of each set were divided into four subgroups of n=8–14, and treated acutely with either 0, 6.25, 12.5, and 25 mg/kg or 0, 25, 50, and 100 mg/kg CBD. In both experiments, CBD was suspended and administered as described above.
Test sessions were conducted immediately after completion of the maintenance phase and lasted 60 min. In test sessions, RR on the alcohol lever was increased progressively according to a procedure slightly adapted from that described by Richardson and Roberts 25 ; namely, RR was increased as follows: 4, 9, 12, 15, 20, 25, 32, 40, 50, 62, 77, 95, 118, 145, 178, 219, etc. The water lever was inactive.
Measured variables were number of responses on each lever and breakpoint for alcohol, defined as the lowest RR not achieved by the rat. Data were statistically evaluated by one-way ANOVA.
Chocolate self-administration in Wistar rats
Training and maintenance phases
The chocolate solution was prepared by diluting powdered Nesquik® (Nestlè Italiana, Milan, Italy; main ingredients: sugar, cocoa powder, soy lecithin, and natural flavors) in tap water. Concentration of Nesquik chocolate powder was 5% (w/v) throughout the study; this concentration resulted in a highly preferred beverage with limited caloric intake (∼0.8 kJ/g). 23
Daily self-administration sessions lasted 60 min. Rats were water deprived exclusively during the 12 h before the first session in the operant chamber. Lever responses were initially shaped under FR1 for 2 sessions; FR was progressively increased from FR1 to FR10 over 10 sessions. Subsequently, 20 additional sessions with FR10 were conducted (maintenance phase). On completion of these 20 self-administration sessions, rats displaying the most stable responding behavior were selected for use in CBD experiments.
Testing under the FR schedule
Similarly to the “alcohol” study, two different experiments were performed, each testing a different CBD dose range and using independent sets of rats. On the test days, rats of each set were divided into four subgroups of n=8–10, and treated acutely with either 0, 6.25, 12.5, and 25 mg/kg or 0, 25, 50, and 100 mg/kg CBD. The experiment testing the higher dose range was designed once data from the experiment testing the lower dose range were available, with the intent of clarifying whether doses of CBD >25 mg/kg would affect chocolate self-administration. In both experiments, CBD was suspended and administered as described above.
Test sessions were conducted immediately after completion of the maintenance phase and were identical to those of the maintenance phase. Measured variables were as follows: number of lever responses for the chocolate solution and amount of self-administered chocolate solution (expressed in mL/kg). Data were statistically evaluated by one-way ANOVA, followed by Tukey's test for post hoc comparisons.
Results
Alcohol self-administration in sP rats
Testing under the FR schedule
Treatment with the low dose range of CBD (0, 6.25, 12.5, and 25 mg/kg) reduced the number of lever responses for alcohol [one-way ANOVA: F(3,36)=5.17, p<0.005] (Fig. 1A). During post hoc analysis, statistical significance was reached by treatment with 12.5 and 25 mg/kg CBD; compared with vehicle treatment, magnitude of the reducing effect averaged ∼55% and 45% in the rat groups treated with 12.5 and 25 mg/kg CBD, respectively. Reduction in number of lever responses resulted in a proportional decrease in the amount of self-administered alcohol [one-way ANOVA: F(3,36)=4.16, p<0.05] (Fig. 1B). During post hoc analysis, statistical significance was reached only by treatment with 12.5 mg/kg CBD.
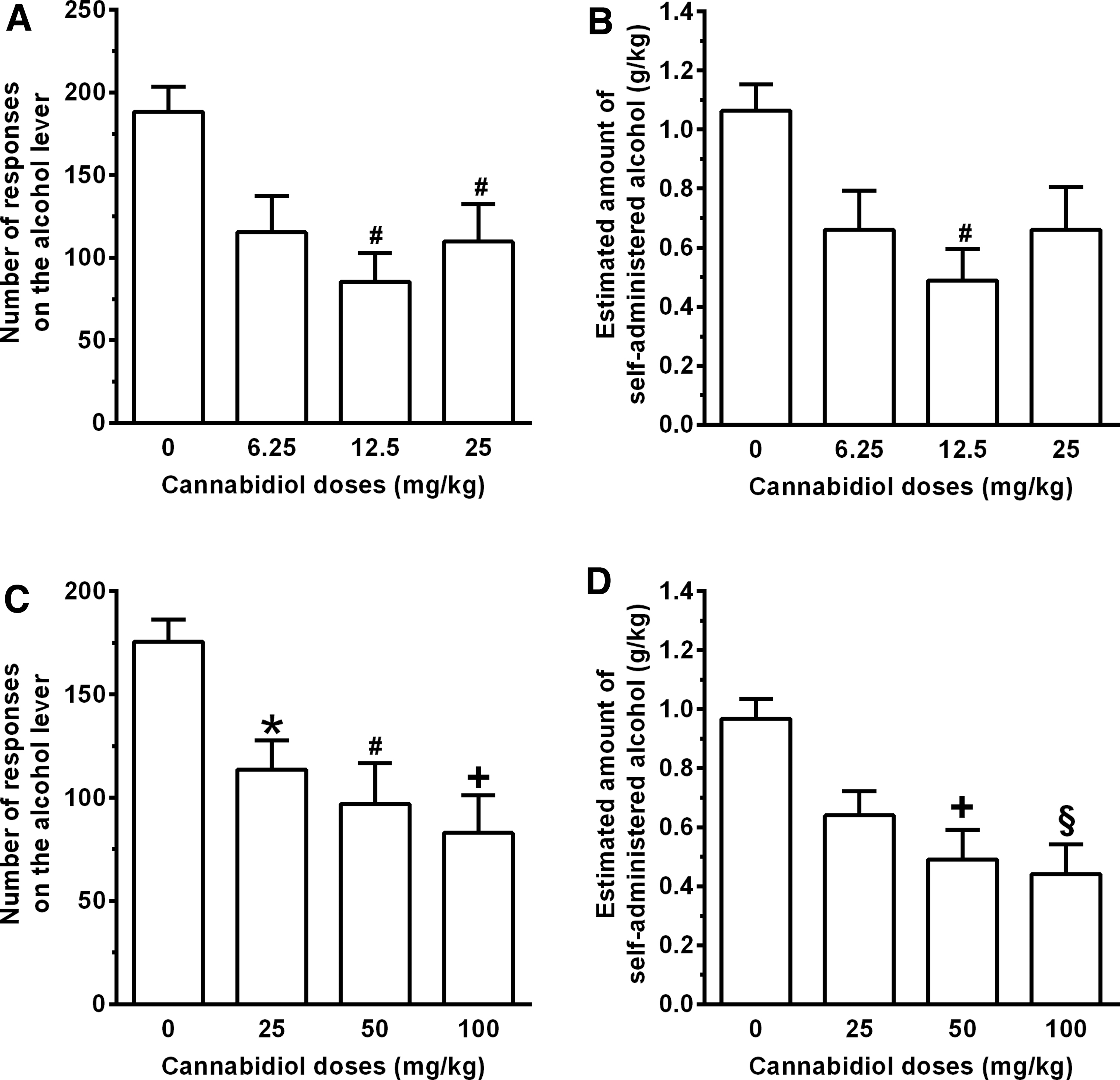
Effect of acute i.p. treatment with two different dose ranges of CBD on number of lever responses for alcohol
Lever responding for water was negligible (averaging <5 per session in all rat groups) and not altered by treatment with CBD [Kruskal–Wallis test: F(3,36)=3.93, p>0.05] (data not shown).
Treatment with the high dose range of CBD (0, 25, 50, and 100 mg/kg) reduced the number of lever responses for alcohol [one-way ANOVA: F(3,44)=6.45, p<0.001] (Fig. 1C). During post hoc analysis, all 3 doses of CBD reduced the number of lever responses for alcohol; compared with vehicle treatment, magnitude of the reducing effect averaged ∼35%, 45%, and 55% in the rat groups treated with 25, 50, and 100 mg/kg CBD, respectively. Reduction in number of lever responses resulted in a proportional decrease in the amount of self-administered alcohol [one-way ANOVA: F(3,44)=7.21, p<0.0005] (Fig. 1D). During post hoc analysis, statistical significance was reached by treatment with 50 and 100 mg/kg CBD.
Lever responding for water was negligible (averaging <4 per session in all rat groups) and not altered by treatment with CBD [Kruskal–Wallis test: F(3,44)=1.24, p>0.05] (data not shown).
Testing under the progressive ratio schedule
Treatment with the low dose range of CBD (0, 6.25, 12.5, and 25 mg/kg) failed to alter both number of lever responses for alcohol [F(3,52)=1.57, p>0.05] (Fig. 2A) and breakpoint for alcohol [F(3,52)=1.93, p>0.05] (Fig. 2B). A slight tendency toward a dose-unrelated reduction was observed in both variables. Responding on the inactive lever was modest (averaging <12 per session in all rat groups) and not altered by treatment with CBD [F(3,52)=2.67, p>0.05] (data not shown).

Effect of acute i.p. treatment with two different dose ranges of CBD on number of lever responses for alcohol
Treatment with the high dose range of CBD (0, 25, 50, and 100 mg/kg) failed to alter both number of lever responses for alcohol [F(3,31)=1.70, p>0.05] (Fig. 2C) and breakpoint for alcohol [F(3,31)=1.46, p>0.05] (Fig. 2D). A slight tendency toward a dose-unrelated reduction was observed in both variables. Responding to the inactive lever was modest (averaging <11 per session in all rat groups) and not altered by treatment with CBD [F(3,31)=1.66, p>0.05] (data not shown).
Chocolate self-administration in Wistar rats
Treatment with the low dose range of CBD (0, 6.25, 12.5, and 25 mg/kg) failed to alter both number of lever responses for the chocolate solution [F(3,28)=2.31, p>0.05] (Fig. 3A) and amount of self-administered chocolate solution [F(3,28)=0.81, p>0.05] (Fig. 3B).

Effect of acute i.p. treatment with two different dose ranges of CBD on number of lever responses for a chocolate solution
Treatment with the high dose range of CBD (0, 25, 50, and 100 mg/kg) failed to alter both number of lever responses for the chocolate solution [F(3,36)=1.04, p>0.05] (Fig. 3C) and amount of self-administered chocolate solution [F(3,36)=0.66, p>0.05] (Fig. 3D).
Discussion
Acute treatment with CBD reduced operant oral alcohol self-administration in sP rats, one of the few rat lines selectively bred worldwide for high alcohol preference and consumption and proposed, because of its predictive validity, as an animal model for testing drugs with therapeutic potential. 26 The effect of CBD on alcohol self-administration was more evident when rats were exposed to the FR schedule of reinforcement (measure of the reinforcing properties of alcohol). 27 Under this schedule, and embracing the two tested dose curves, the reducing effect of CBD ranged from the minimally effective dose of 12.5–100 mg/kg.
The effect of CBD was selective for alcohol, as none of these doses of CBD affected self-administration of the chocolate solution. The latter data indicate that the reducing effect of CBD on alcohol self-administration was not secondary to any sedative or unspecific effect.
Conversely, when rats were exposed to the PR schedule of reinforcement (measure of the motivational properties of alcohol), 27 the effect of CBD was limited to a slight tendency toward a reduction in lever responding and breakpoint for alcohol. Together, these data suggest that the reinforcing and motivational properties of alcohol are, at least in sP rats, differentially sensitive to the reducing effect of CBD.
The results of this study confirm, and extend to a model of genetically determined excessive alcohol drinking, previous experimental data indicating the ability of CBD to attenuate different alcohol-motivated behaviors in rodents, including reinstatement of alcohol seeking in rats 13 and alcohol drinking and self-administration in mice.14–16 Notably, all the studies conducted to date (including the present investigation) have used experimental procedures proved to model different aspects of human AUD,13–16 conferring translational validity to the collected results.
Anxiolysis is a major pharmacological effect of CBD, with potential clinical application.28,29 CBD-induced amelioration of anxiety-related behaviors has repeatedly been reported in rodents.13,30–34 Anxiolysis may also provide a key to understanding the mechanism underlying the reducing effect of CBD on alcohol self-administration in sP rats. Besides high alcohol preference and consumption, sP rats display inherent predisposition to anxiety-related behaviors.35–40 Voluntarily consumed alcohol markedly reduced these anxiety-related behaviors,35,41 suggesting that alcohol-induced anxiolysis is a major factor driving sP rats to seek and consume alcohol. Accordingly, it is conceivable that the anxiolytic effects of CBD may have substituted for those of alcohol, making alcohol seeking and drinking less urgent and resulting in the observed reduction in alcohol self-administration. If theoretically transposed to humans, these data suggest that CBD would represent a potentially effective therapeutic option for those patients with comorbid anxiety and AUD.
Besides the eCB system, CBD is known to interact with diverse signaling systems that contribute to mediating its multiple pharmacological effects.6,7 These systems include serotonin 5-HT1A receptor, nuclear peroxisome proliferator-activated receptor-γ, transient receptor potential of the melastatin type 8 channel, transient receptor potential of ankyrin type 1 channel, transient receptor potential of vanilloid types 1 and 2 channels, α1 and α2 glycine receptor, adenosine A2A receptor, G protein-coupled receptor-55, and μ and δ opioid receptor.6,7
Notably, several of these signaling systems are part of the neural substrate mediating the central effects of alcohol, 42 and their pharmacological activation or blockade modulates alcohol-seeking and -drinking behaviors in rodents, 43 and—at least as far as the opioid receptor system is concerned—in humans. 44 The reducing effect of CBD on alcohol self-administration observed in this study might therefore be the sum of several different actions of CBD at multiple molecular targets.
The experiments testing the effect of CBD on chocolate self-administration intended to address two different research questions: (i) to assess the selectivity of the reducing effect of CBD on alcohol self-administration, using a nondrug reinforcer as a comparison (see above); (ii) to contribute new experimental data to a relevant research issue such as the ability of CBD to modulate the appetitive and consummatory aspects of palatable food. Despite the large dose range tested (considering both experiments), data indicate that no dose of CBD altered chocolate self-administration.
These results apparently contrast with those suggesting the ability of CBD to reduce consumption or self-administration of sucrose solutions in rats and mice.20,21,22 Differences in the experimental procedures may certainly contribute to explaining these differences in CBD efficacy on consumption and self-administration of palatable food. For example, the chocolate solution used in this study generates addictive-like high levels of lever responding. Levels of lever responding as high as those recorded under this experimental procedure, averaging >2000, 23 have seldom been observed even with the most addictive drugs and are indicative of remarkably strong reinforcing and motivational properties. Vernacularly speaking, in a theoretical hierarchy of substances inducing strong reinforcing and motivational properties, this chocolate solution would equate to, or even surpass, several highly addictive drugs. It is therefore conceivable that the antiaddictive attributes of CBD are insufficient to affect reinforcing properties as strong as those generated by this chocolate solution, being conversely effective on those—much weaker—produced by sucrose solutions. 21
Finally, with the caution needed when comparing animal and human data, one can speculate that the lack of effect of CBD treatment on chocolate self-administration observed in this study resembles the inability of a CBD-enriched Cannabis preparation to modify ad libitum intake of the most preferred chocolate milk-like drink in humans 45 ; inhalation of this CBD-enriched Cannabis preparation also failed to alter sweet taste intensity perception and liking of the favorite drink. 45
Conclusion
The results of this study indicate that acutely administered CBD effectively and selectively reduced the reinforcing properties of alcohol in selectively bred alcohol-preferring sP rats. These results extend to an animal model of excessive alcohol consumption the recent lines of experimental evidence on the reducing effects of CBD on alcohol-motivated behaviors. Conversely, acute treatment with a high dose range of CBD failed to affect the reinforcing properties of a highly palatable food in rats.
Footnotes
Acknowledgments
The authors are grateful to Mrs. Carla Acciaro for animal breeding and care, and Ms. Anne Farmer for language editing of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work did not receive any specific funding.