
Abstract
Purpose:
Recent research has suggested that chronic alcohol exposure induces changes in the endocannabinoid system within the central nervous system and therefore could be an attractive target for better understanding and treating alcohol use disorder (AUD). Much of this research has centered around the CB1 receptor and its endogenous partial agonist, the endocannabinoid anandamide, as the CB1 receptor is densely expressed in brain regions involved in development and maintenance of addictive behaviors. In addition, recent evidence has suggested that chronic alcohol exposure induces changes in the modulation of endocannabinoid concentration and suggests that these changes may contribute to the motivation to abuse alcohol. Therefore, we performed a systematic literature review to evaluate how fatty acid amide hydrolase (FAAH), an enzyme that degrades anandamide, relates to the characteristics and biology of AUD, as well as how modulating FAAH through pharmacologic inhibition or genetic manipulation affects outcomes related to alcohol use and consumption.
Method:
A search strategy was developed using the terms “endocannabinoids” or “drug delivery systems” and “alcohol dependence” or “alcohol use disorder” or “alcoholism” and “Fatty Acid Amide Hydrolase” and “FAAH” as text words and Medical Subject Headings (i.e., MeSH and EMTREE). We then used this search strategy on the electronic databases PubMed, Embase, and Web of Science.
Results:
We found 224 records; after removing repeated records (37%), articles that did not fit the topic question (47%), or were not primary research (4%), we included 26 for qualitative synthesis (12%).
Discussion:
The literature clearly suggests that FAAH has a role in the biology and characteristics of AUD. FAAH inhibition seems especially promising as a target for alcohol withdrawal as it may lead to a reduction in symptoms, including anxiety and a reduction of alcohol intake reinstatement. However, decreased FAAH may also lead to reduced sensitivity to alcohol along with increased preference and intake.
Conclusions:
Modulation of FAAH is promising for therapeutic intervention of AUD, but requires more research. Pre-clinical studies have indicated that FAAH inhibition may reduce withdrawal characteristics, but may also exacerbate other characteristics of AUD outside of that period.
Introduction
Alcohol use disorder (AUD) is a chronic relapsing disorder that has significant public health burdens. According to the 2018 National Survey on Drug Use and Health, 14 million adults aged 18 years and over and 401,000 adolescents aged 12–17 years have AUD. 1 In addition, a recently published study found that the rate of alcohol related deaths per year among people aged 16 years and older doubled from 1999 to 2017, and nearly half of these deaths were from liver disease or overdoses. 2 However, only 7.9% of adults and 5.0% of youths who had AUD in 2018 received treatment. 1 Furthermore, there are few effective treatment methods currently available for treating AUD. The U.S. Food and Drug Administration (FDA) has approved only three medications for treating alcohol dependence, including naltrexone, acamprosate, and disulfiram, each of which have relative contraindications that can limit their widespread use. Therefore, research is needed to identify additional targets for the treatment of AUD.
Promising targets for novel pharmacotherapy of AUD include the endocannabinoid system in the central nervous system (CNS). The endocannabinoid system includes endogenous cannabinoids (endocannabinoids), the enzymes that synthesize and degrade the endocannabinoids, and the receptors to which the endocannabinoids bind. 3 The discovery of the endocannabinoid system's role in modulating neurotransmitter release and neuroadaptations in the past few decades have led to increasing evidence that it is involved in the initiation and progression of many psychiatric disorders, including substance use disorders. 4 Many of these therapies targeted the cannabinoid CB1 receptor, which is densely expressed in brain regions involved in development and maintenance of addictive behaviors, and modulation of these receptors has downstream behavioral effects. 5 For instance, there is evidence that CB1 agonists increase, and CB1 antagonists or inverse agonists suppress, alcohol consumption in rats.6,7 Based on pre-clinical evidence, the CB1 receptor antagonist/inverse agonist rimonabant was investigated in human trials for its possible treatment of AUD. Unfortunately, two human clinical trials found no effect of rimonabant on relapse to drinking or alcohol self-administration.8,9
More recently, research has suggested that chronic alcohol exposure also induces changes in the modulation of endocannabinoid concentration and suggests that these changes may contribute to the motivation to abuse alcohol. 10 Therefore, other targets in the endocannabinoid system are being explored, including the enzyme that degrades the endocannabinoid anandamide (AEA). AEA is a partial agonist for the CB1 receptor and is hydrolyzed by the membrane bound enzyme fatty acid amide hydrolase (FAAH) following its release into the synapse and reentry into the neuronal membrane. FAAH, therefore, effectively regulates the concentration of AEA available for stimulating CB1 receptors. With advent of more selective FAAH inhibitors, some of which have entered into clinical trials, we were curious to see if sufficient evidence exists demonstrating that FAAH expression or activity is associated with AUD and if FAAH inhibitors could be a potential treatment of AUD.
The scope of this systematic review is focused on how FAAH relates to the characteristics and biology of AUD, as well as how modulating FAAH through pharmacology or genetic manipulation affects outcomes related to alcohol use and consumption. We conducted a literature search to answer the questions: What does current literature report about how FAAH is related to the characteristics and biology of AUD? How does modulating FAAH through pharmacology or genetic manipulation affect outcomes related to alcohol use and consumption? For a more comprehensive review on the endocannabinoid system and AUD or the endocannabinoid system and addiction more broadly see Basavarajappa et al., and Sloan et al.10,11
Methods
A literature review was conducted on June 26, 2020. To provide wide coverage of available studies, we searched PubMed (endocannabinoids OR drug delivery systems AND alcohol dependence OR alcohol use disorder AND Fatty Acid Amide Hydrolase OR FAAH OR “fatty-acid amide hydrolase”), Embase (Endocannabinoid/exp AND alcoholism/exp AND FAAH AND EMBASE), and Web of Science (ALL=endocannabinoid OR endocannabinoids OR drug delivery system AND alcohol use disorder OR alcoholism OR alcohol dependence AND FAAH OR fatty acid amide hydrolase).
Inclusion criteria
Primary research in which the investigation included FAAH and AUD or salient characteristics of AUD for which results of our search were available, including, but not limited to, susceptibility to AUD, sensitivity to alcohol, withdrawal, relapse, and/or the preference and consumption of alcohol.
Exclusion criteria
Results that included FAAH and aspects of AUD not related to CNS or behavioral effects. Reviews were utilized for background information but excluded from the analysis.
Results
Search results
The PubMed search identified 52 articles, Embase identified 83 articles, and Web of Science identified 87 articles for a total of 222. Eighty-three records were repeated, leaving a total of 139 records to be screened.
The 139 remaining article abstracts were reviewed; 76 were eliminated as they did not fit topic question, and 10 were eliminated because they were reviews and not primary research articles. Twenty-five of these results were considered “Grey Literature,” they were evaluated for relevant content that did not make it to publication (e.g., conference abstracts), and all were ultimately eliminated from inclusion in this review. The 28 remaining articles went to full-text review. Four were eliminated due to not being related to topic question. Two additional sources were identified through screening of included article references and included. In total, 26 articles were included in this literature review (Fig. 1).
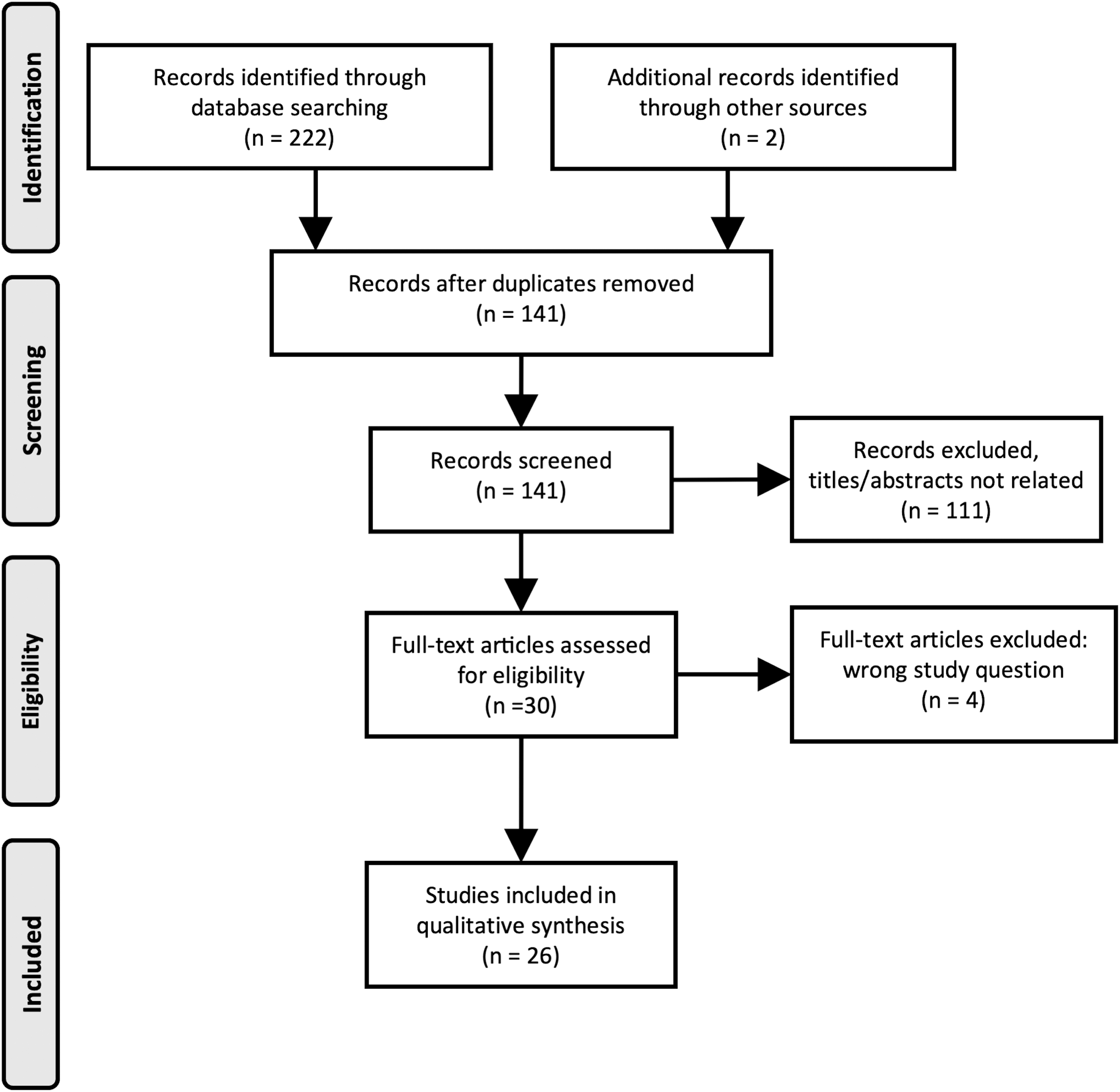
Flow diagram of the search strategy. Original research articles were identified and selected for inclusion using the preferred reporting items for systematic reviews or meta-analyses (PRISMA).
The data extracted from the included articles were organized into subsections based on a conception of the clinical course of AUD development and characteristic of AUD that was studied. They were further synthesized into groups based on the species studied, method of FAAH manipulation, and outcome to more clearly compare results.
Of the 26 citations, ∼70% of these were pre-clinical studies involving rodents and 30% of these were clinical studies. There have been no clinical trials using FAAH inhibitors in humans in relation to AUD to date. In addition, 70% included data using genetic manipulation or analysis, 20% included use of a FAAH inhibitor, while 10% included data using both (Table 1).
Characteristics of Citations Reviewed
Discussion
Our literature search found that all reported research to date conducted on the biological connection of FAAH to AUD, and potential treatment of FAAH inhibitors for AUD, have been pre-clinical, and the majority of it was in rodents. As no clinical trials in humans have been conducted, no direct conclusions about the utility of FAAH inhibitors in patients with AUD can be made. However, this literature review can serve to provide inferences about the prospects of such a novel treatment. The following subsections are organized based on available data regarding FAAH and corresponding to the time course of AUD pathology starting from premorbid risk followed by initial exposure, repeated consumption, withdrawal, and other consequences of alcohol consumption, with studies in humans, when available, described first. As sex-based differences in the endocannabinoid system and AUD have been previously described, we also reviewed the available data on sex-related differences of FAAH and alcohol consumption.
Expression and function of FAAH and association to AUD susceptibility
Human genetic studies have suggested that a common single nucleotide polymorphism in FAAH, the gene encoding FAAH, is associated with substance use disorders. This polymorphism is a missense mutation C385A (rs324420), which converts a proline residue to threonine (Pro129Thr). The missense mutation results in a FAAH variant that has normal catalytic properties but is more susceptible to proteolytic degradation. 12 Therefore, the C385A variant leads to reduced FAAH activity leading to increased anandamide levels in humans with the A/A or A/C genotype. 13
This genotype has been associated with characteristics that contribute to AUD. The C385A/A genotype is associated with a trending effect of lower FAAH levels in the brain and is strongly associated with self-reported problem drug and alcohol use and current AUD in Caucasians.14–17 However, this genotype does not appear to be associated with AUD in African American or Japanese study populations.15,18 These differences may be explained by differences in study design. The lack of association in African Americans may be attributed to socioeconomic factors as the African American study participants had a markedly lower income bracket than the European American participants. In addition, the Iwasaki et al. study in the Japanese population required AUD diagnosis based on the DSM IV criteria, whereas the other studies looked at self-reported problem alcohol use. In contrast to all previous listed results, Bühler et al. observed an association between the wild-type genotype (C/C) and an increase in risky drinking behavior in Caucasian college students. 19 Although the results have been inconsistent, the majority of the sources reviewed here have hypothesized that the C385A genotype is associated with worse outcome in AUD, likely due to increased anandamide. However, these associations have not clearly addressed if FAAH dysregulation confers risk of heavy or chronic alcohol use or if alcohol use leads to FAAH dysregulation.
In part to address this, corresponding studies in alcohol preferring rats have supported an association between dysregulated FAAH before any alcohol exposure. FAAH expression and activity is significantly decreased in the prefrontal cortex in alcohol preferring (AA) rats and in the hippocampus of alcohol preferring (sP) rats.20,21 However, FAAH expression and activity in these rats are not significantly altered in the striatum or cerebellum. 20
Therefore, the human and animal studies listed above suggest that innate differences in FAAH gene expression, protein levels, and activity might be associated to susceptibility and development of risky alcohol use in some populations. However, these differences are region dependent and could indicate that some aspects of AUD are more likely to be associated than others.
Changes in FAAH after alcohol exposure
Several lines of evidence suggest that alcohol leads to lower FAAH activity. FAAH activity is significantly lower in the ventral striatum of patients with alcohol dependence compared to that of controls, as determined from postmortem samples and independent of suicide as the cause of death. 22 This is in agreement with an animal study which found that FAAH enzyme levels and activity are significantly decreased in the striatum of alcohol preferring rats following 60 days of alcohol exposure. 21 In addition, FAAH activity is significantly decreased in the hippocampus of rats following acute alcohol administration, in the cortex of rats who were forced to drink alcohol for 24 h, and in the cortex of mice that were made alcohol tolerant by exposure to alcohol vapors for 72 h.23–25 Furthermore, FAAH activity is significantly decreased in cerebellar granular neurons that had been harvested from rats and then exposed to alcohol for 72 h. 26
In contrast, a postmortem examination with a very similar design to Vinod et al., observed that FAAH levels and activity in the prefrontal cortex were not significantly altered in patients with alcohol abuse, whether or not suicide was the manner of death. 27 In animal models it was also found that FAAH levels were not significantly altered in the amygdala, caudate, putamen, nucleus accumbens, or cerebellum in rats following acute alcohol exposure or in alcohol dependence models (Fig. 2 and Table 2).24,25,28
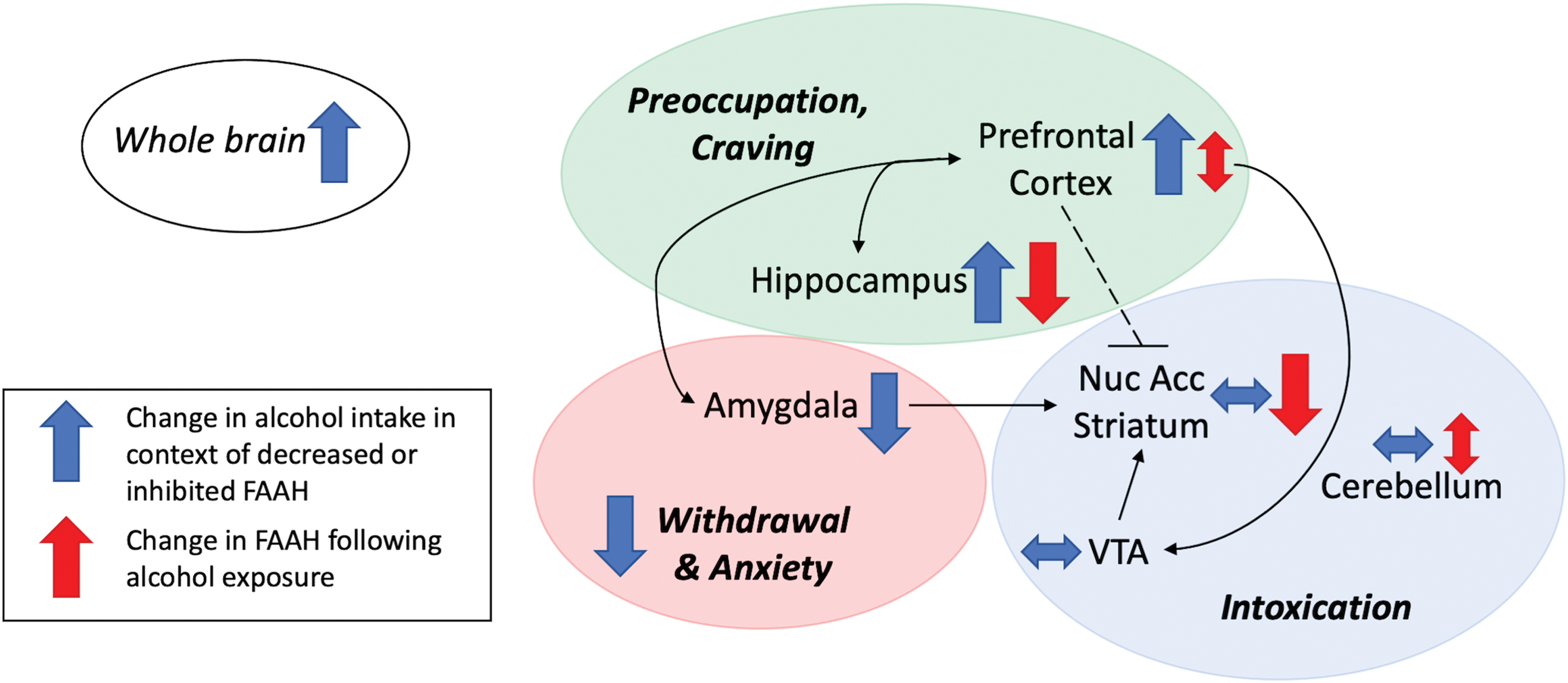
Schematic overview of interrelationships between FAAH activity and alcohol exposure, consumption, or associated risk. Brain regions and circuitry (black arrows; dashed line identifies inhibitory control) are highlighted for their role in an established model of reward pathways. Shaded circles overlap brain regions associated with salient features of AUD: green=regions associated with alcohol preoccupation and craving; blue=regions associated with alcohol intoxication and reward salience; red=region associated with alcohol withdrawal and associated anxiety. Blue arrows indicate resulting change in alcohol consumption or associated risks with AUD when FAAH is either decreased (by genetic manipulation or C385A genotype) or pharmacologically inhibited in the targeted area. Red arrows indicate regional change in FAAH expression and/or activity following alcohol exposure. Double-headed horizontal arrows indicate no change in results; double-headed vertical arrows indicate mixed results retrieved in the literature review. Whole brain FAAH inhibition or reduction by genetic deletion or mutation leads to an increase of alcohol consumption and AUD associated risk. Nuc Acc, nucleus accumbens; VTA, ventral tegmental area; FAAH, fatty acid amide hydrolase; AUD, alcohol use disorder. Color images are available online.
Fatty Acid Amide Hydrolase Activity Following Alcohol Exposure
Therefore, it appears that alcohol exposure may lead to decreases in FAAH levels and activity; however, the changes appear to be region dependent.
Effect of FAAH on alcohol preference and consumption
Reduced FAAH levels in the brain are associated with risky alcohol use in human and animal studies. Whole brain FAAH levels are inversely correlated with the number of self-reported drinks per week. 16 This is in agreement with the finding that the C385A/A genotype, which leads to lower FAAH levels in the brain, is associated with a higher number of drinking and binge drinking days. 15 However, although the genotype may be associated with risky drinking or increased alcohol use, it was not found to be associated with clinical features of AUD as diagnosed in the DSM IV. 16
To investigate FAAH's effects on alcohol preference and consumption, FAAH activity has been decreased by insertion of genetic missense mutation (C385A), genetic ablation, and pharmacological manipulation in rodents with various models of alcohol exposure. For a review of the various models of alcohol exposure in rodents see Jeanblanc et al. and Planeta et al.29,30 Genetically modified mice with lower FAAH activity due to missense mutation had increased preference for and consumed significantly more alcohol at baseline during a drinking in the dark paradigm than wild-type mice. 31 In addition, male and female FAAH knockout (KO) mice had increased preference for, and consumed significantly more, alcohol in a two-bottle choice paradigm compared to their wild-type counterparts.32,33 However, Basavarajappa et al., found that the results were significant only for female mice, while Blednov et al., found that the results were significant only for male mice with a trending effect in female mice. The difference could be attributed to study design. The wild-type mice Basavarajappa et al., used had higher baseline alcohol consumption than those used by Blednov et al., which may have limited the potential increase of drinking driven by the mutation or the sensitivity to detect a further increase of alcohol intake. In addition, Blednov et al., found that the mice had increased preference and consumed more alcohol at lower alcohol concentrations (6, 9%) with no significance at the higher concentrations (12, 20%) used by Basavarajappa et al. Male FAAH KO mice also showed increased preference for alcohol in an unlimited access paradigm. 34
The association between decreased FAAH activity and alcohol consumption was also found pharmacologically. Male wild-type mice that received the FAAH inhibitor URB597 (0.5 mg/kg i.p.) had increased preference for, and consumed significantly more alcohol, in the two-bottle choice paradigm compared to vehicle injected controls. 33 In addition, male Wistar rats that received high dose URB597 (4 μg) into the prefrontal cortex consumed significantly more alcohol after the animals had developed a stable baseline of alcohol intake in saccharin-fading self-administration protocol compared to low dose URB597 (0.4 μg) or vehicle treated controls. 20 Male mice who received URB597 (0.1 mg/kg i.p.) had a significantly increased preference for alcohol when using the unlimited access protocol compared to vehicle treated mice. 34 Therefore, there is evidence that reduced FAAH, whether through genetic means or pharmacological inhibition, increases alcohol preference and consumption.
However, additional studies suggest that the neuroanatomical location of FAAH inhibition differentially affects alcohol consumption. In contrast to rats injected with URB597 in the prefrontal cortex, male Wistar rats similarly injected in the central and basolateral amygdala (0.1 or 1.0 μg) consumed significantly less alcohol after the animals had developed a stable baseline of alcohol intake through the saccharin-fading self-administration protocol. 35 Yet, in other models, FAAH inhibition did not alter alcohol intake at all. Male Wistar or msP rats that received URB597 peripherally (0.1, 0.3, or 1 mg/kg) or into the ventral tegmental area (1.0 μg) did not alter alcohol intake after the animals had developed a stable baseline of alcohol self-administration through the saccharin-fading self-administration protocol (Fig. 2 and Table 3).35,36
Fatty Acid Amide Hydrolase and Sensitivity to Alcohol
Therefore, while evidence in human and mice studies consistently point to increased alcohol preference and consumption in reduced FAAH settings, the rat models suggest that local activity of FAAH can confer differential outcomes on alcohol consumption.
FAAH and sensitivity to alcohol
Reduction in FAAH may decrease sensitivity to alcohol in mice, as assessed by blood alcohol content, alcohol induced hypothermia, duration of loss of righting reflex (LORR), and alcohol-induced ambulatory movement. There does not appear to be any difference in blood alcohol content or difference in alcohol clearance between FAAH KO and wild-type littermates.32,33,37
However, results on alcohol induced hypothermia seem to be dependent on gender and time after alcohol exposure. One group found that alcohol-induced hypothermia was significantly decreased in male FAAH KO mice at 15 min, but not 30 min, after alcohol challenge (2 g/kg). 34 In addition, alcohol-induced hypothermia was significantly decreased in female FAAH KO mice 30 min after alcohol challenge (1.5 g/kg), but was not significantly altered in male FAAH KO mice at 30 min. 32
In a similar manner, duration of LORR was significantly decreased in male FAAH KO mice after an acute dose of alcohol (3, 3.2, or 3.4 g/kg) compared to wild type.33,34 In addition, Basavarajappa et al., observed that duration of LORR was significantly decreased in FAAH KO mice after an acute dose of alcohol (4 g/kg); however, this decrease was limited to female mice and there were no significant changes between male FAAH KO and wild-type mice. 32
Results of alcohol-induced ambulatory movement following FAAH manipulations were inconsistent. One group found that male and female FAAH KO mice recovered more quickly from the loss of coordination than wild-type mice, 33 while another group did not observe any significant differences based on genotype (Table 4). 34
Fatty Acid Amide Hydrolase and Alcohol Consumption
i.p., intraperitoneal; PFC, prefrontal cortex; CeA, central amygdala; BLA, basolateral amygdala; VTA, ventral tegmental area.
There is evidence that individuals with increased sensitivity to alcohol, such as increased flushing, are less likely to develop AUD. 38 Therefore, as the connection between sensitivity to alcohol and alcohol consumption has already been established, this evidence from rodent studies may indicate a mechanism where decreased FAAH leads to decreased sensitivity to alcohol as measured by the decreased alcohol induced hyperthermia and duration of LORR, which may increase the risk of development of AUD.
The role of FAAH in withdrawal and relapse
FAAH activity and levels may be decreased in early withdrawal. Best et al., found that patients with AUD had significantly lower FAAH activity globally in early abstinence (3–7 days) compared to healthy controls, as assessed by positron emission tomography. 16 However, they did not find that the lower FAAH levels correlated with craving, withdrawal severity, or days of abstinence. In addition, by weeks 2–4 of abstinence, FAAH activity was no longer significantly different from healthy controls. Furthermore, FAAH levels in early abstinence were not predictive of relapse before the second scan in weeks 2–4 of abstinence, as FAAH levels were not significantly different in nonabstinent patients compared with patients who had maintained abstinence.
In rodents, levels of FAAH protein and FAAH mRNA after chronic alcohol exposure are dependent on model of alcohol exposure and length of withdrawal. There is a significant difference in FAAH mRNA in the amygdala of rats in early abstinence (24 h) after 3 weeks of continuous alcohol exposure. 39 However, there is no significant difference in gene expression or FAAH levels in the amygdala or prefrontal cortex of rats in early (24 h) or longer term (1–2 weeks) abstinence after 3 weeks of intermittent alcohol exposure.39,40 Therefore, it seems that FAAH levels in male rodents may be affected at 24 h in the first withdrawal but not following intermittent exposure or repeated withdrawals.
As the negative effects of withdrawal are a likely drive to relapse drinking, it is important to consider whether FAAH is associated with withdrawal severity or anxiety. Male FAAH KO mice had a significant reduction in severity of handling-induced convulsions following a 6–9 h withdrawal from chronic alcohol exposure through alcohol vapors. 34 However, FAAH KO mice did not differ in severity of handling-induced convulsions during a 5–13 h withdrawal from acute alcohol exposure. 33 FAAH inhibition may ameliorate the anxiogenic effects of alcohol withdrawal both in early withdrawal (12 h) and late withdrawal (7 days). Injection of URB597 (0.1, 0.3, or 1.0 mg/kg i.p) into male rats that had been given a single large dose of alcohol followed by 12 h of withdrawal prevented the increase in anxiety-like response that vehicle treated rats displayed. 36 In addition, in a model where alcohol dependence was established, followed by 7 days of abstinence, there was significantly less anxiety-like behavior in rats and mice in alcohol withdrawal that had been treated with the FAAH inhibitor PF-3845 (3 or 10 mg/kg i.p.) than rats and mice that had been treated with vehicle. 28
FAAH inhibition reduced alcohol consumption in mice in early withdrawal. Male and female mice that received a single dose of URB597 (0.125, 0.25, 0.5, or 1.0 mg/kg i.p.) had decreased preference and consumed significantly less alcohol after intermittent access to alcohol for 3 weeks followed by a 1-day withdrawal or drinking in the dark paradigm for 3 weeks followed by a 1-day withdrawal. 40 FAAH inhibition reduced alcohol consumption in mice with the alcohol deprivation effect, a model used to mimic the relapse-like drinking effect. Male and female mice are given intermittent access to alcohol for 3 weeks, the alcohol is removed for 7 days, then alcohol is reintroduced, and consumption is measured. Alcohol intake upon reintroduction was significantly reduced in mice who had received either a single dose of URB597 (0.5 mg/kg) 30 min before reintroduction of alcohol or repeated doses of URB597 over 5 days, with the last dose 1 day before reintroduction of alcohol. 40 In addition, male Wistar rats and C57BL/6J mice that received PF-3845 (3 or 10 mg/kg) consumed significantly less alcohol after the animals had become alcohol dependent through the alcohol inhalation method followed by a 7-day withdrawal compared to vehicle treated controls. 28
However, in rats, FAAH inhibition had no effect on cue-induced or stress-induced reinstatement of alcohol use. Male Wistar rats were trained to self-administer alcohol and discriminate alcohol from water based on differing sensory cues. They were then subjected to 2 weeks of extinction where sensory cues were presented but no alcohol was dispensed. Finally, rats were treated with URB597 (0.1, 0.3, or 1.0 mg/kg i.p.) and then given access to alcohol. However, URB597 did not modify cue-induced reinstatement of alcohol seeking behavior. 36 In addition, URB597 did not have an effect on stress-induced reinstatement of alcohol seeking, whether through foot shock or yohimbine treatment (Table 5). 36
Fatty Acid Amide Hydrolase and Withdrawal
One significant factor contributing to alcohol use reinstatement is drive to reduce the negatively reinforcing effects of alcohol, primarily expressed in alcohol withdrawal. 41 There is robust evidence that FAAH inhibition may ameliorate the anxiety that accompanies withdrawal from alcohol and withdrawal symptom severity. In addition, models consistently show that FAAH is dysregulated in early withdrawal, and FAAH inhibition in early withdrawal may reduce reinstatement of alcohol consumption. Therefore, FAAH inhibition could be an attractive target to prevent relapse during initial withdrawal by reducing severity of withdrawal symptoms.
The role of FAAH in alcohol-related neurodegeneration and neuroinflammation
FAAH inhibition may reduce alcohol induced memory loss. Male Wistar rats subjected to injections of alcohol for 3 days in an acute alcohol binge model had an increase in recognition index in long-term memory if they received URB597 (0.3 mg/kg i.p.). 42 However, URB597 did not alter long-term memory in a model of chronic alcohol binging, in which rats were subjected to injections of alcohol 3 days each week for 4 weeks. URB597 did not alter short-term memory in either acute or chronic alcohol binge models. Of note, this study did not observe memory impairment induced by alcohol administration, although there was a trend of long-term memory impairment in both groups.
Furthermore, the same study found that FAAH inhibition decreases inflammatory mediators. Male Wistar rats in the chronic alcohol binge model had increased pro-inflammatory cytokines interferon gamma and TNF alpha in the prefrontal cortex and hippocampus, but these cytokines were reduced in rats that received URB597. 42 In addition, rats that were given URB597 had reduced interferon gamma in the prefrontal cortex in the acute binge model, but this effect was not observed in the hippocampus. Administration of URB597 in chronic alcohol binge model did not result in an increase in brain-derived neurotrophic factor in the hippocampus.
However, FAAH inhibition during an alcohol binge-like model was not neuroprotective. Administration of URB597 (0.3 mg/kg i.p.) every 12 h during the binge alcohol model, in which rats were injected with alcohol every 8 h for 4 days, did not reduce number of necrotic cells (assessed by FJB+ cells) in the ventral dentate gyrus or entorhinal cortex compared to alcohol alone or alcohol + vehicle groups (Table 6). 43
Fatty Acid Amide Hydrolase and Protection from Neuroinflammation/Degeneration in Alcohol Binge
FJB, Fluoro-Jade B; BDNF, brain derived neurotrophic factor.
FAAH and sex differences
There are significant sex differences in several aspects of alcohol consumption, consequences of alcohol use, and development of AUD. 44 Therefore, as FAAH is influenced by gonadal hormones, it is important to examine potential sex differences as it relates to AUD. 45
When evaluating FAAH levels in patients with AUD and healthy controls, Best et al. reported a significant difference in sex proportions between groups. 16 They observed a nonsignificant trend for higher FAAH levels in female participants, whether in AUD or controls.
Basavarajappa et al., found that female FAAH KO mice consumed more of both 12% and 20% alcohol and had higher alcohol preference when offered a choice between water and alcohol compared to male FAAH KO mice and wild-type counterparts of either sex. 32 Meanwhile, although male FAAH KO mice had increased preference for 20% alcohol compared to their wild-type counterparts, they did not show increased preference for 12% alcohol or increased intake of 12% or 20% alcohol compared to their wild-type counterparts. As an aside, female FAAH KO mice also had a significant increase in overall liquid consumption of 12% alcohol compared to any of the other groups. However, there was no difference between genotypes or sexes when it came to taste preference for saccharin or quinine, suggesting that increased alcohol preference was not due to altered taste preference. In addition, female FAAH KO mice were also less sensitive to the alcohol hypothermic and sedative effects after an acute 1.5 g/kg dose of alcohol. Finally, although there was no difference in alcohol levels between KO and wild-type mice, female mice of both genotypes had lower alcohol levels.
In contrast, Blednov et al., did not find such clear sex-based differences. 33 In their study, there were no significant differences when examining genetic manipulation of FAAH on alcohol intake and preference, although the results for male mice were significant compared to wild type and the results for female mice were not. Furthermore, Zhou et al. found no significant differences based on sex in mice when examining pharmacological FAAH inhibition on alcohol intake and preference. 40
Conclusion
Our systematic review demonstrates that research in humans and rodents clearly suggest that FAAH has a role in the biology and characteristics of AUD. FAAH inhibition seems to be promising as a possible target for alcohol withdrawal. During withdrawal, FAAH inhibition may lead to a reduction of symptoms and a reduction of reinstatement alcohol intake. In addition, FAAH inhibition also seems to be especially promising for its anxiolytic effects after exposure to alcohol. Whether FAAH inhibition could be helpful for treating comorbid anxiety disorders in the context of AUD remains uncertain and may benefit from additional research.
However, several results point toward the fact that chronic or acute alcohol exposure may lead to reduced FAAH activity in the brain. Furthermore, the research indicates that decreased FAAH, whether through genetic or pharmacological inhibition, may result in reduced sensitivity to alcohol and lead to increased intake and increased preference for alcohol. Thus, we might speculate that alcohol exposure leads to neuroadaptations of reduced FAAH expression to reduce sensitivity to alcohol. While the mechanism remains unknown, it is conceivable that reduced FAAH, leading to increased anandamide, contribute to neuroprotective effects countering the neurotoxic effects of alcohol and, during withdrawal, excessive glutamate release. Conversely, that lower FAAH activity is associated with worse outcomes in chronic AUD suggests that FAAH inhibition might actually worsen the course of AUD outside of the period of withdrawal.
Alcohol consumption has increased in the past two decades, particularly among women. There has been a 10.1% increase in prevalence of drinking and 23.3% increase in binge drinking among women, while overall prevalence of drinking and binge drinking did not change for men. 2 Surprisingly, few studies attempted to clarify the role of FAAH in AUD in female animal models. Therefore, we suggest that further research include both sexes and be appropriately powered to analyze effect of sex in the results. With regard to research in females, future studies may need to additionally control or adjust for estrous cycle phase or estrogen/progesterone levels, which have been largely absent from the literature in our review.
Ultimately, targeting FAAH is complicated and AUD is a complex multifaceted issue. To date, there have only been a few clinical trials testing FAAH inhibitors in humans, and none has examined AUD. Interestingly, there is new evidence which shows that FAAH is an allosteric enzyme. 46 This new evidence opens up the possibility to discover and design new nonsubstrate molecules that can potentiate or attenuate the efficacy of FAAH inhibitors, opening up additional avenues to FAAH targeted therapies.
This review expands upon recent reviews but has limitations of its own. Recent reviews by Basavarajappa et al., and Kunos examined a more expanded approach of the ECS and its interplay in AUD examining both pre-clinical and clinical data.10,47 By narrowing our focus, we aimed to have a more thorough investigation to evaluate the efficacy of targeting FAAH as a possible treatment of AUD. However, by limiting our review to FAAH, we have not closely examined interplay between the use of FAAH inhibitors or FAAH expression on the other elements of the ECS, including CB1 receptors, 2-arachidonoylglycerol (2-AG), and monoacylglycerol lipase (MAG-L), or the indirect effects of AEA catabolic inhibition, including altered levels of arachidonic acid or secondary metabolism of AEA through cyclooxygenase. Furthermore, this review did not cover effects that FAAH inhibition may have in other areas outside of the endocannabinoid system; it is important to understand that anandamide acts at receptors other than CB1. For example, anandamide also acts at peroxisome proliferator-activated receptors, which are ligand-activated transcription factors, and therefore modulate gene transcription. 48 Finally, by limiting our investigation to AUD, we minimized or ignored common comorbidities, such as anxiety, depression, and post-traumatic stress disorder, and how the characteristics of these psychiatric disorders may be jointly influenced by FAAH inhibition.
Our understanding of the role of FAAH in AUD remains incomplete, and more research is needed. The effects of FAAH on the biology and characteristics of AUD appear to be dependent on alcohol dose and method of exposure, time course of alcohol exposure or withdrawal, species and strain of rodent, brain region, and biological sex and therefore require further study.
Footnotes
Acknowledgment
The authors thank Dr. Kevin Measor for review of an earlier draft of this article and helpful comments.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by a Career Development Award #CX-001787 from the United States (U.S.) Department of Veterans Affairs Clinical Sciences Research and Development Service (GET). The contents do not represent the views of the U.S. Department of Veterans Affairs or the United States Government.