
Abstract
Introduction:
Alpha/beta-hydrolase domain 6 (ABHD6) is an enzyme that hydrolyzes 2-arachidonoylglycerol, a high-efficiency endogenous cannabinoid. Although the endocannabinoid system has been suggested to be involved in regulation of bladder function, the roles of ABHD6 in the control of micturition remain unknown. To elucidate the physiological and pathological roles of ABHD6 in vivo, we examined phenotypes of ABHD6 knockout rats (Abhd6−/−) generated by clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated proteins system.
Materials and Methods:
Age-matched knockout and wild-type (WT) rats of both sexes were used.
Results:
Expression of ABHD6, assessed by quantitative real-time polymerase chain reaction and Western blot analysis, was clearly diminished in Abhd6−/− rats compared with WT rats. Mutant rats had a normal appearance, and the body weight and food consumption were similar to those of WT rats. The interval between bladder contractions assessed by continuous cystometry was significantly shorter in ABHD6 knockout rats than in WT rats when the bladder was stimulated with acetic acid. Mechanical paw withdrawal thresholds measured by von Frey testing were significantly lowered in the knockout rats than in WT rats. The plasma levels of prostaglandin E2 (PGE2) and the stable metabolite of PGE2 in Abhd6–/– rats were twice as high as that in WT rats.
Conclusions:
Deletion of the ABHD6 gene in rats causes more frequent urination in the stimulated bladder and hyperalgesia to non-noxious mechanical stimuli along with increased plasma PGE2.
Introduction
The endocannabinoid signaling system is involved in a variety of physiological functions and in disease states, including moderation of pain and inflammation. 1 Alpha/beta-hydrolase domain containing 6 (ABHD6) is a serine hydrolase that has been recently demonstrated to regulate hydrolysis of the endogenous cannabinoid 2-arachidonoylglycerol (2-AG).2,3 Since 2-AG is the most abundant endocannabinoid and a full agonist for cannabinoid receptors (CB1/CB2), it is assumed that suppression of ABHD6 activity would lead to activation of the cannabinoid receptor-mediated endocannabinoid signaling.
The endocannabinoid system has also been suggested to be involved in regulation of normal bladder function 4 and in the pathogenesis of overactive bladder in humans 5 and rats.6,7 ABHD6-deficient mice have been utilized in several studies8–12 of the physiological and pathological roles of ABHD6 in vivo. To date, however, there is no study concerning the role of ABHD6 in regulation of lower urinary tract function. Since previous experiments in rats with a cannabinoid agonist or with an endocannabinoid-degrading enzyme inhibitor have shown to diminish irritation-induced bladder overactivity,7,13 we supposed that ABHD6-deficient rats would be insensitive to various stimuli, and could alleviate frequent urination in various kinds of experimental models of lower urinary tract symptoms.14–16
This study was conducted to display preliminary characterization and urodynamics of ABHD6 knockout rats created with the clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated proteins (Cas9) system.
Materials and Methods
Animals
Generation of founder animals lacking ABHD6 on a Sprague-Dawley (SD) outbred genetic background using CRISPR and Cas9 system was performed by KAC Co. Ltd. (Kyoto, Japan). The sequence of the oligonucleotide for guide RNA was as follows: 5′-GGAAAGAAGCTACGAAGGCCAGG-3′. A mixture of guide RNA (50 ng/μL) and Cas9 mRNA (100 ng/μL) was microinjected into SD pronuclear stage embryos to generate the Abhd6−/− genotype. Embryos were subsequently transferred to pseudopregnant female rats. The selected male founders were mated with female wild-type (WT) SD rats (sourced from CLEA Japan, Inc., Tokyo, Japan) to produce heterozygous Abhd6+/– rats. The resulting heterozygous offspring were subsequently paired to generate homozygous Abhd6−/− rats. To identify the founder and homozygous animals, genotyping was performed on tail biopsies using forward 5′-cccactacggtttgttttccaggc-3′ and reverse 5′-gggcacccaaaacccttttctggc-3′ primers flanking the target site. Knockout rat production was maintained by breeding homozygous pairs.
A total of 46 Abhd6−/− and 40 age-matched WT rats of both sexes weighing 200–500 g were used in this study. The study protocol was approved by the institutional animal care and use committee of the University of the Ryukyus.
Quantitative polymerase chain reaction
Expression of Abhd6 mRNA in the bladder, cerebrum, and liver was examined in two WT and two knockout rats. After the tissue was dissected free and weighed, total RNA was extracted from tissue by using an RNeasy Mini Kit™ (Qiagen, Hilden, Germany). After treatment with DNase I to avoid genomic contamination, 1 μg of RNA was used for synthesis of cDNA with SuperScript™ II Reverse Transcriptase (Thermo Fisher Scientific). The primer sets of Abhd6 and glyceraldehyde phosphate dehydrogenase (GAPDH) were as follows: Abhd6, 5′-tcaaggtcttgtcgacgttc-3′ (forward) and 5′-tggtcttgttttccccaaat-3′ (reverse); GAPDH, 5′-ggcattgctctcaatgacaa-3′ (forward) and 5′-atgtaggccatgaggtccac-3′ (reverse).
Relative expression of target genes was analyzed by performing quantitative real-time polymerase chain reaction (RT-qPCR) with TB Green™ Premix Ex Taq™ II (Tli RNaseH Plus) (Takara, Japan) and the ViiA 7 Real-Time PCR System (Thermo Fisher Scientific) under the following conditions: initial denaturation at 95°C for 30 sec and 40 cycles of 95°C for 5 sec and 60°C for 30 sec. Expression of the target gene was normalized for the geometric mean level of GAPDH expression.
Western blot analysis
Immediately after euthanasia, the bladder was isolated and stabilized in Allprotect Tissue Reagent™ (Qiagen). The tissue was homogenized in 0.5 mL of ice-cold homogenization buffer (20 mM Tris-HCl [pH 7.5], 2 mM ethylenediamine tetraacetic acid (EDTA), 2 mM ethylene glycol tetraacetic acid (EGTA), and 0.5% Triton X-100) containing protease inhibitor cocktail (no. 25955-11, NACALAI TESQUE, INC., Kyoto, Japan) and phosphatase inhibitor cocktail (no. 07575-51; NACALAI TESQUE, INC.). After homogenization, the tissue homogenate was centrifuged at 15,000 g for 10 min at 4°C, and the supernatant was used for Western blot analysis. We solubilized 120 μg protein in Laemmli sample buffer containing 2.5% of 2-mercaptoethanol at 98°C for 5 min, and subjected it to 8% sodium dodecyl sulfate–polyacrylamide gel electrophoresis at room temperature. The gels were transferred to polyvinylidene difluoride (PVDF) membranes, which were blocked with 5% nonfat milk. They were then incubated with anti-ABHD6 and anti-α-tubulin antibodies (1:1000) in 5% nonfat milk and Tris-buffered saline-Tween. After being washed and incubated with a horseradish peroxidase-conjugated antirabbit IgG antibody (1:5000), the membranes were developed with an enhanced chemiluminescence system (Western Lightning ECL Pro; PerkinElmer, Inc., Waltham, MA). Quantitative densitometry was performed by a lumino-image analyzer (LAS-4000 mini EPUV; Fujifilm) and commercial software (Multi Gauge version 3.0; Fujifilm).
Continuous cystometry
Female WT (n=3) and Abhd6−/− (n=5) rats were anesthetized with urethane (0.8 g/kg subcutaneously and 0.4 g/kg intraperitoneally) and body weight was measured. To enable continuous cystometry, a PE-50 polyethylene catheter (Clay Adams, Parsippany, NJ) was inserted into the bladder through the urethra. Physiological saline was infused through the catheter at 0.05 mL/min and bladder activity was monitored through the urethral catheter, which was connected to a pressure transducer and a saline infusion pump through a three-way stopcock. When bladder contractions occurred, urinary flow also occurred around the urethral catheter. Cystometry was continued for at least 1 h and the intervals between bladder contractions, baseline intravesical pressure, threshold bladder pressure, maximum bladder contraction pressure, and bladder contraction duration were measured during the final 30 min.
After continuous cystometry with physiological saline, the rats also underwent continuous cystometry with 0.25% acetic acid solution instead of physiological saline. 17 Cystometry was continued at least 1 h, and the same measurements were acquired during the final 30 min.
von Frey measurement
For the mechanical withdrawal test, rats were placed individually on a mesh cage plane in a transparent plastic box and were allowed to adapt to the testing environment for at least 15 min. Mechanical sensitivity to non-noxious stimuli was assessed using an electronic von Frey apparatus (BIO-EVF5; Bioseb Instrument, Vitrolles, France). 18 The von Frey test was applied to the plantar surface of each hind paw, through the mesh floor. Every rat was subjected to two tests with an interval of 3 min, and the mean was taken as the withdrawal threshold. To assess the effect of various analgesics including diclofenac sodium (25 mg), tramadol hydrochloride (50 mg), pregabalin (75 mg), imipramine hydrochloride (25 mg), neurotropin (8 U), and rizatriptan benzoate (10 mg), the threshold values obtained before and 2 h after oral administration were compared. Doses of these analgesics were determined by referring to the clinical doses.
Cold allodynia
To quantify cold sensitivity of the foot, brisk foot withdrawal in response to acetone application was measured in 8 male WT rats and 11 male Abhd6−/− rats, as reported previously. 19 The acetone was applied five times (once every 5 min) to each paw. The frequency of foot withdrawal was expressed as a percent: (number of trials accompanied by brisk foot withdrawal)×100/(number of total trials).
Measurement of 2-AG and arachidonic acid in cerebrospinal fluid
Cerebrospinal levels of 2-AG and arachidonic acid in three female WT rats and three female Abhd6−/− rats were determined by a liquid chromatograph/time-of-flight mass spectrometer analysis (Supplementary Data S1) carried out through a facility of service at Human Metabolome Technologies, Inc. (Tsuruoka, Japan). Under isoflurane anesthesia, cerebrospinal fluid was collected through a PE50 tube inserted into the vertebral cavity. The samples were immediately cooled on ice and stored at −80°C until analyzed.
Measurement of plasma prostaglandin E2 and PGE metabolites
Blood was collected from six female WT rats and six female Abhd6−/− rats for the measurement of plasma levels of prostaglandin E2 (PGE2) and 13,14-dihydro-15-keto prostaglandin E2 (PGEM), a stable metabolite of PGE2. Under isoflurane anesthesia, blood was drawn from the inferior vena cava into a 7 mL EDTA-2Na tube (Venoject II; Terumo Co., Tokyo, Japan). The samples were cooled on ice and then centrifuged at 1000 g for 10 min. The plasma was stored at −80°C until analyzed. Plasma samples were assayed in duplicate for PGE2 and PGEM by enzyme immunoassay using commercial kits (no. 514010 and 514531, respectively; Cayman Chemical Company, Ann Arbor, MI, ).
Data analysis
The data were analyzed by Student's t-test or analysis of variance (ANOVA) followed by Bonferroni's post hoc test. All statistical tests were performed using the computer program Prism 7.0 (GraphPad Software, Inc., CA). A value of p<0.05 was considered to be statistically significant. All results are expressed as means±standard error of the mean.
Results
General characteristics of ABHD6 knockout rats
Expression of ABHD6 was examined by RT-qPCR analysis and Western blot analysis in the Abhd6−/− rats selected by DNA genotyping (Fig. 1a). Expression of ABHD6 in bladder, brain, and liver in Abhd6–/– rats at the transcriptional level was markedly lower compared with that in WT rats (Fig. 1b). Bladder ABHD6 protein in Abhd6–/– rats was also significantly lower, 7.6% of that of WT rats as shown in Figure 1c (p<0.001).
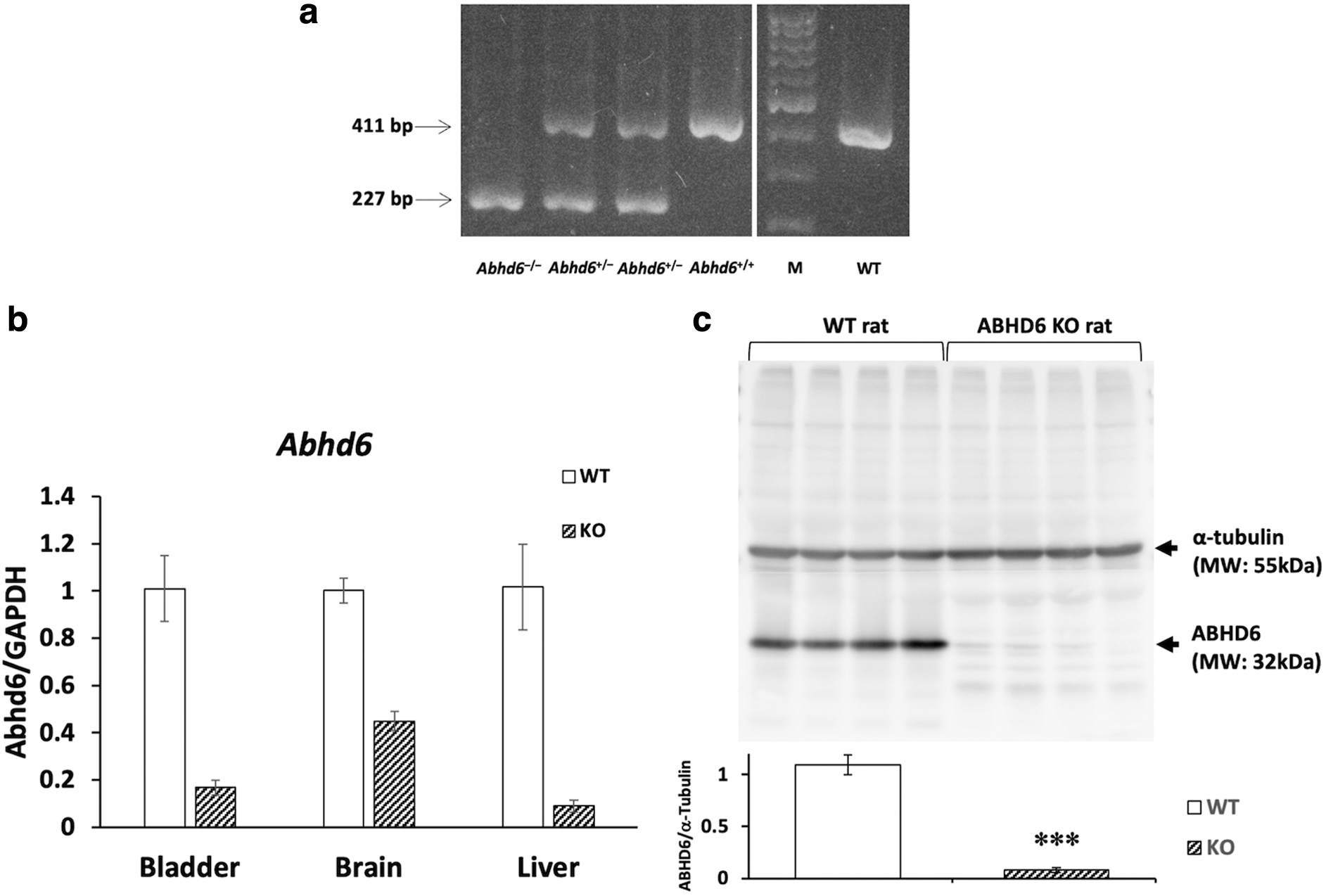
PCR analysis of DNA from tail tissues in Abhd6-184 del and WT rats
The appearance of Abhd6–/– rats was indistinguishable from that of WT rats. The body weight of the ABHD6 knockout rats of both sexes was similar to the corresponding age-matched WT rats (Fig. 2). Daily food consumption (7.1±0.4% of body weight per day, n=6) in knockout rats was also compatible with that (7.2±0.2% of body weight per day, n=4) in WT rats.
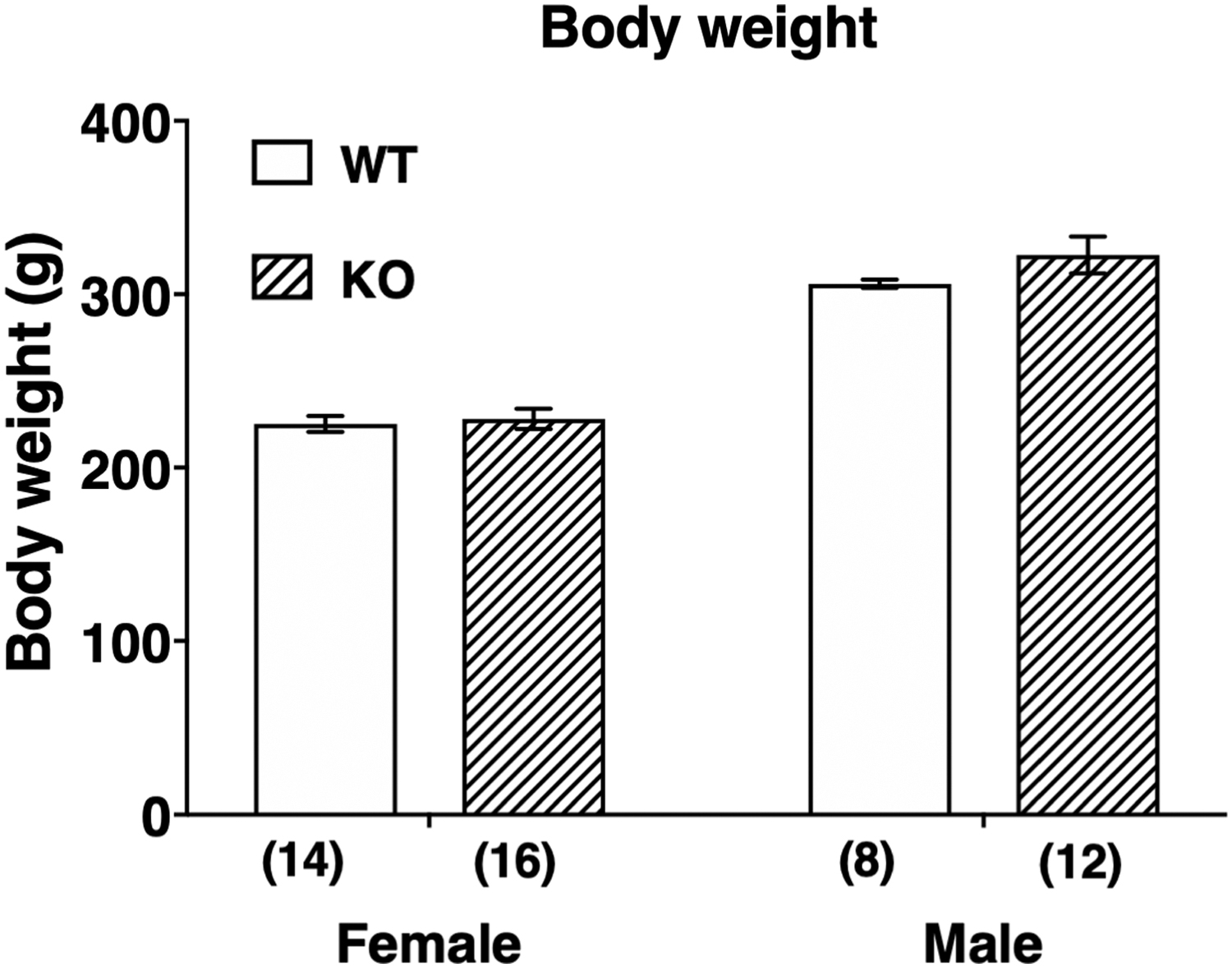
Body weight in female and male WT and ABHD6 knockout rats. Number in parentheses indicates number of animals. Data are means±SEM. There was no significant difference between WT and knockout rats as determined by two-way ANOVA followed by a Bonferroni's multiple comparison test. ANOVA, analysis of variance.
Continuous cystometry
During continuous cystometry with physiological saline, there were no significant differences of any cystometric parameters between Abhd6–/– and WT rats (Fig. 3). When continuous cystometry was done with the 0.25% acetic acid solution, the interval between bladder contractions in ABHD6 knockout rats was significantly shorter than that in the WT rats (p<0.01), indicating increased sensitivity to bladder stimulation compared with normal rats. The duration of bladder contraction was not significantly different between knockout and WT rats (data not shown).
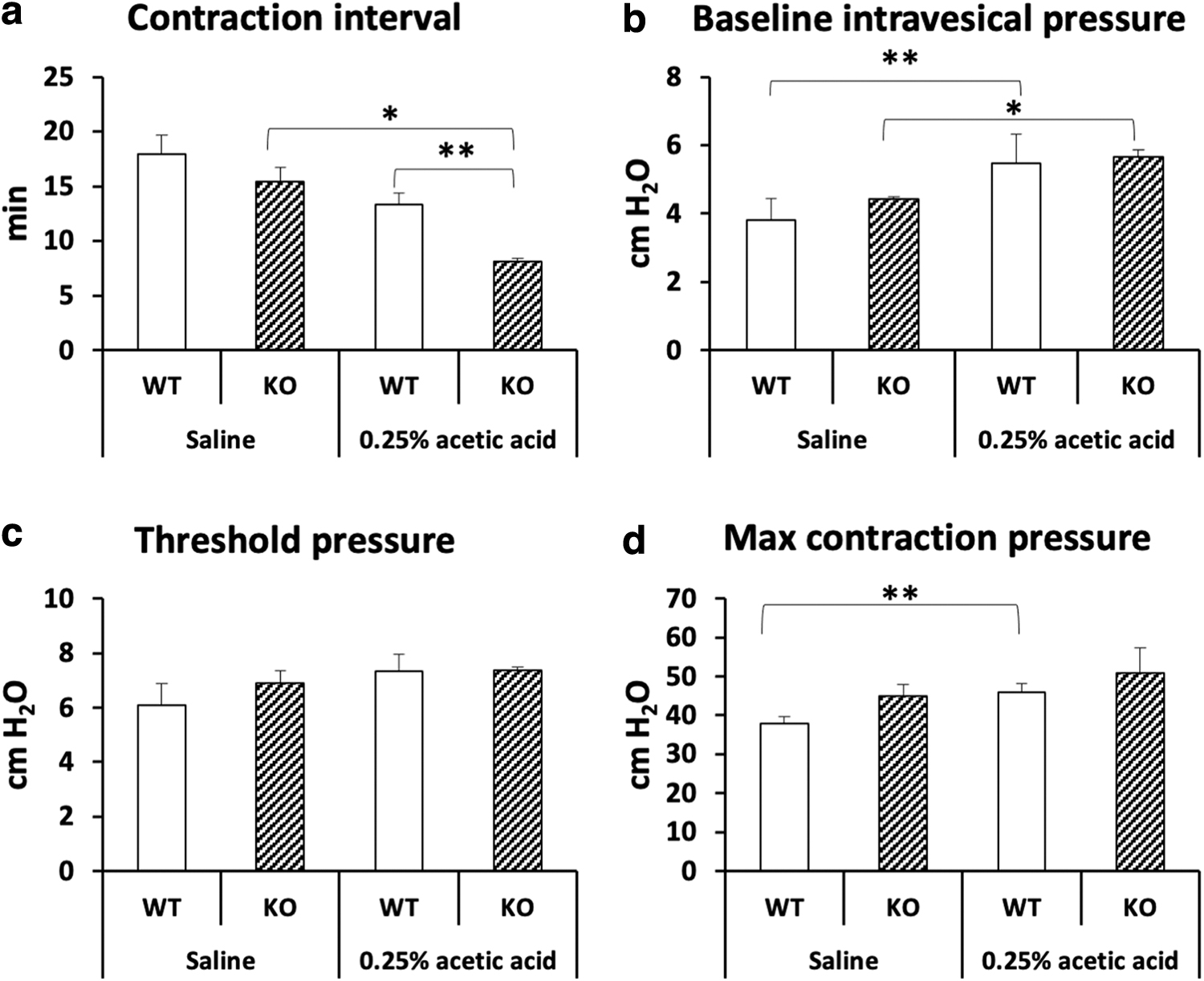
Cystomeric parameters during continuous cystometry in female WT and ABHD6 knockout rats intravesically treated with saline or 0.25% acetic acid.
von Frey test
Both female and male Abhd6–/– rats showed significant decreases in withdrawal threshold compared with the corresponding WT rats (p<0.01 each, Fig. 4), indicating a hyperalgesic response of the paw to non-noxious stimuli.
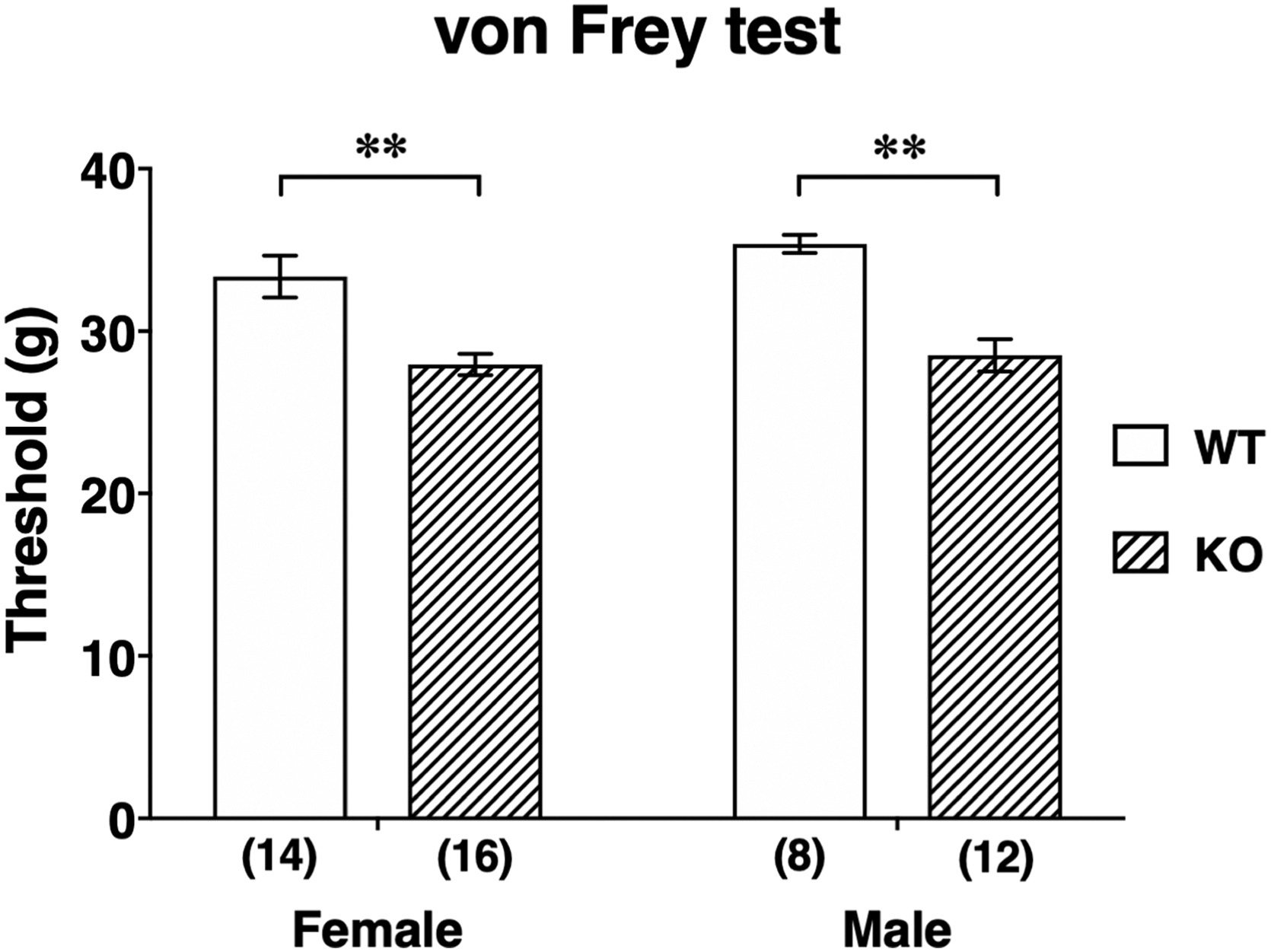
Withdrawal threshold values of von Frey test in female and male WT and ABHD6 knockout rats. Numbers in parentheses indicate number of animals. Data are means±SEM. **p<0.01 by two-way ANOVA followed by a Bonferroni's multiple comparison test.
The effect of the analgesics with mechanisms of action on the withdrawal threshold using male Abhd6–/– rats (Fig. 5) revealed that oral diclofenac (a cyclooxygenase inhibitor), tramadol (a synthetic opioid), pregabalin (a voltage-dependent Ca2+ channel blocker), imipramine (a 5-HT/NA reuptake inhibitor), and neurotropin (an activator of the descending inhibitory pathway) but not rizatriptan (a 5-HT1B/1D agonist) significantly raised the threshold values (Fig. 5a–f). However, the data must be interpreted with caution, because effects of these analgesics at the same dose were not examined in WT rats.

Effect of various analgesics on threshold values of von Frey test in male ABHD6 knockout rats.
The frequencies of brisk foot withdrawal in response to acetone application in Abhd6–/– rats (n=11) and WT rats (n=8) were 15.5±2.8% and 21.3±6.4%, respectively, which was not statistically different (p=0.373).
Measurement of 2-AG and arachidonic acid in cerebrospinal fluid
Cerebrospinal 2-AG was undetectable in WT rats, whereas the 2-AG in ABHD6 knockout rats was detected (8.4±2.5, relative area; Supplementary Fig. S1a). The cerebrospinal level of arachidonic acid, a degradation product of 2-AG, in ABHD6 knockout rats (56.6±45.1, relative area) was 6.9-fold greater than the level in WT rats (8.2±4.9, relative area; Supplementary Fig. S1b), although the difference was not statistically significant (p=0.352).
Measurement of plasma PGE2 and PGE metabolites
The plasma level of PGE2 in Abhd6–/– rats (266.8±44.4 pg/mL) was more than twice the level in WT rats (125.6±9.2 pg/mL, p<0.05; Fig. 6a). The plasma level of PGEM, a stable metabolite of PGE2, in Abhd6–/– rats (231.0±10.9 pg/mL) was also twice as high as that in WT rats (113.5±14.0 pg/mL, p<0.001; Fig. 6b).

Plasma levels of PGE2
Discussion
ABHD6 is a serine hydrolase that degrades monoacylglycerol lipids, particularly the endogenous cannabinoid 2-AG, in both central and peripheral tissues. 20 It seems likely that phenotypes of ABHD6-deficient animals would presumably be analogous to the multiple effects produced by chronic treatment of WT animals with a selective inhibitor of the enzyme. In contrast to our expectations, our results have shown that deficiency of ABHD6 in rats causes more frequent urination in the stimulated bladder and hyperalgesia to non-noxious mechanical stimuli when compared with the WT.
ABHD6 knockout rats showed urinary frequency in response to intravesical acetic acid-induced nociceptive stimuli without a difference in baseline bladder function compared with WT rats. These results were unexpected, because lack of ABHD6 would be expected to attenuate irritation-induced bladder overactivity, most likely through enhanced cannabinoid signaling in the bladder mechanoafferent function, as shown in previous experiments in rats with a cannabinoid agonist or with an endocannabinoid-degrading enzyme inhibitor.7,13 The reason for the discrepancy between the present results obtained with ABHD6 knockout rats and the mentioned findings cannot be explained, but a difference in the treatment period (acute vs. chronic) might be implicated in the conflicting results.
In line with the mentioned urodynamic study showing bladder overactivity when stimulated with acetic acid, ABHD6 knockout rats exhibited decreased withdrawal threshold to mechanical non-nociceptive stimuli assessed by the von Frey test, likely indicating mechanical allodynia or hyperalgesia. Although the mechanisms involved in the phenotype observed remain unknown at present, there may be some possible factors to be considered: first, according to a previous study, 21 genetic disruption of monoacylglycerol lipase (MAGL) causes a profound increase in brain levels of 2-AG and desensitization of CB1 receptors, and results in tolerance to the antiallodynic effects of CB1 receptor activation. If such tolerance, at least partly, occurs in the case of ABHD6 deletion, it may be possible to explain the results that analgesic effects were not observed in ABHD6 knockout rats. Second, it is plausible that prostaglandins such as PGE2 and PGI2 may play a role in the irritability of ABHD6 knockout rats, because these are known to enhance the sensitivity of sensory neurons to various modes of stimulation. Direct administration of prostaglandins to the spinal cord causes hyperalgesia and allodynia. 22 In addition, it is possible that hydrolysis of accumulated 2-AG by MAGL, a primary 2-AG hydrolytic enzyme, contributes to the brain levels of arachidonic acid, which, in turn, converts to PGE2 by catalysis of cyclooxygenase-1/2. 23 Interestingly, a recent study has shown that deletion of ABHD12, another 2-AG hydrolyzing enzyme, causes significant elevations in brain levels of not only 2-AG but also arachidonic acid and PGE2. 24 Furthermore, 2-AG can be oxygenated by cyclooxygenase-2, and the metabolite of 2-AG includes PGE2 glycerol ester, 25 which has been demonstrated to induce hyperalgesia in vivo. 26 Thus, we evaluated circulating levels of prostaglandins in ABHD6 knockout rats, and found statistically significant twofold increases in plasma levels of PGE2 and the stable metabolite of PGE2. Accordingly, it is likely that the increased sensitivity to non-noxious stimuli in ABHD6 knockout rats is attributable, at least, in part, to the increase in PGE2. Furthermore, the present findings that administration of diclofenac, a cyclooxygenase inhibitor, dampened the withdrawal response of knockout rats may support this view. Third, recent in vitro studies have unveiled that ABHD6 is part of the α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor macromolecular complex, and inactivation of ABHD6 by CRISPR/Cas9 or shRNAs increases AMPA receptor-mediated synaptic transmission and the resultant glutamatergic transmission independently of CB receptors.27,28 According to these findings, lack of ABHD6 might cause enhanced pain sensation through activation of AMPA receptor. It is obvious, however, that further experiments such as pharmacological approaches using selective inhibitors are needed to clarify roles of the receptors or enzymes concerned in the phenotypes.
In conclusion, we demonstrated that genetic disruption of Abhd6 in rats causes frequent urination in the stimulated bladder and hyperalgesia to non-noxious mechanical stimuli, and suggest possible involvement of increased PGE2 in the irritability of ABHD6 knockout rats.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.