
Abstract
Background:
Cannabidiol (CBD), one major nonintoxicating phytocannabinoid from Cannabis sativa demonstrated anti-inflammatory effects in animal models of several inflammatory conditions, including arthritis. However, it is still unknown which cell types mediate these anti-inflammatory effects of CBD, and, since CBD binds to a plethora of receptors and enzymes, it is complicated to pinpoint its mechanism of action. In this study, we elucidate the effects of CBD on B cells and peripheral blood mononuclear cells (PBMCs) in respect to survival, calcium mobilization, drug uptake, and cytokine (IL-6, IL-10, and TNF) and immunoglobulin production.
Methods:
Modulation of intracellular calcium and drug uptake in B cells was determined by using the fluorescent dyes Cal-520 and PoPo3, respectively. Cytokine and immunoglobulin production was evaluated by enzyme-linked immunosorbent assay. PBMC composition and B cell survival after CBD treatment was assessed by flow cytometry.
Results:
B cells express two major target receptors for CBD, TRPV2 (transient receptor potential vanilloid 2) and TRPA1 (transient receptor potential ankyrin 1), which are not regulated by B cell activation. CBD increased intracellular calcium levels in mouse and human B cells, which was accompanied by enhanced uptake of PoPo3. These effects were not dependent on transient receptor potential channel activation. CBD increased the number of early apoptotic B cells at the expense of viable cells and diminished interleukin (IL)-10 and tumor necrosis factor (TNF) production when activated T cell independently. In PBMCs, CBD increased IL-10 production when B cells were activated T cell dependent, while decreasing TNF levels when activated T cell independently. In PBMC/rheumatoid synovial fibroblast cocultures, CBD reduced IL-10 production when B cells were activated T cell independently. Immunoglobulin M production was augmented by CBD when B cells were activated with CpG.
Conclusion:
CBD is able to provide pro- and anti-inflammatory effects in isolated B cells and PBMCs. This is dependent on the activating stimulus (T cell dependent or independent) and concentration of CBD. Therefore, CBD might be used to dampen B cell activity in autoimmune conditions such as rheumatoid arthritis, in which B cells are activated by specific autoantigens.
Introduction
Cannabis sativa contains a plethora of bioactive compounds with phytocannabinoids and terpenes being the most prominent. Δ 9 -Tetrahydrocannabinol (THC) and cannabidiol (CBD) have been investigated in more detail since they are currently used in medical practice. 1 However, although THC and CBD are considered as anti-inflammatory, their impact on chronic inflammation in autoimmune diseases has not been addressed adequately. While THC binds to classical cannabinoid receptors (CB1 and CB2) with high affinity, CBD targets several receptors, enzymes, and transporters and has only negligible affinity for cannabinoid receptors.2,3 In contrast to THC, CBD is nonintoxicating and therefore has a better adverse event profile than THC containing drugs.
Rheumatoid arthritis (RA) is characterized by joint erosions, excessive proinflammatory cytokine, and autoantibody production. 4 In synovial tissue, resident synovial fibroblasts (SF) expand due to inhibition of apoptosis and they are activated by interactions with lymphocytes. This leads to cytokine, chemokine, and matrix metalloproteinase (MMP) production by rheumatoid arthritis synovial fibroblasts (RASF), which further enhances the recruitment of immune cells. 5 Therefore, current therapies of RA aim to reduce cytokine burden, which not only prevents excessive immune cell activation but also reduces proinflammatory effects mediated by RASF. 6
In previous studies, we already demonstrated an inhibitory effect of CBD on RASF cytokine production and proliferation, 3 but, it is yet unclear how CBD affects B lymphocytes or fibroblast/peripheral blood mononuclear cells (PBMCs) cocultures. In this study, we assessed not only the effects of CBD in isolated human and mouse B lymphocytes but also its impact on PBMCs alone and in coculture with RASF mimicking the environment in synovial tissue. This study might help to clarify the capacity of CBD to treat chronic, autoimmune inflammation and whether CBD might be used as adjunct therapy in RA.
Materials and Methods
Biochemicals
Upon arrival, all compounds were dissolved in DMSO (exception: AEA [pre-dissolved in ethanol]; ISO and RR [dissolved in ddH2O]), aliquoted and kept at −20°C in the dark. Before experiments, working solutions in Hank's balanced salt solution (HBSS) or medium were prepared. Each aliquot was only used once and was discarded after use. All compounds used in this study are listed in Table 1.
Biochemicals Used in This Study
The concentrations used in this study are based on values found in the literature.
DIDS, 4,4′-diisothiocyanatostilbene-2,2′-disulfonate; TLR9, toll-like receptor 9; TNF, tumor necrosis factor.
Patients
Six patients with long-standing RA fulfilling the American College of Rheumatology revised criteria for RA 7 were included in this study. The RA group comprised only females with a mean age of 68.8±12.0 years. C-reactive protein was 9±13 mg/dL. Two out of six patients received glucocorticoids, three out of six methotrexate, one out of six sulfasalazine, and one out of six biologicals. All patients underwent elective knee joint replacement surgery, and they were informed about the purpose of the study and gave written consent. The study was approved by the Ethics Committees of the University of Düsseldorf (Approval No. 2018-87-KFogU). We confirm that all experiments were performed in accordance with relevant guidelines and regulations.
SF and tissue preparation
Samples from RA synovial tissue were collected immediately after opening the knee joint capsule, and tissue was prepared for cell isolation thereafter. 8 Synovial tissue was cut into small fragments and treated with liberase (Roche Diagnostics, Mannheim, Germany) at 37°C overnight. The cell suspension was filtered (70 μm) and centrifuged at 300 g for 10 min. After that, the pellet was treated with erythrocyte lysis buffer (20.7 g NH4Cl, 1.97 g NH4HCO3, 0.09 g EDTA ad 1 L H2O) for 5 min, recentrifuged for 10 min and then resuspended in phenol red-free RPMI-1640 (#R7509; Sigma Aldrich, Taufkirchen, Germany) with 10% fetal bovine serum (FBS, #10500-064; Gibco/Thermo Fisher), 1% penicillin/streptomycin (#P4333-100ML; Sigma), 1% GlutaMAX (#35050-038; Thermo Fisher), 1% sodium pyruvate (#S8636; Sigma), and 25 mM HEPES (#H0887-100ML; Sigma).
Cells were maintained at 37°C and 5% CO2. After overnight incubation, RPMI medium was replaced with fresh medium to wash off dead cells and debris.
Western blotting
The following antibodies were used: transient receptor potential ankyrin 1 (TRPA1) (1 mg/mL, 1:5000, NB110-40763; Novus Biologicals, Centennial, CO), and transient receptor potential vanilloid (TRPV)2 (1:1500, NBP1-32096; Novus Biologicals). One million cells were lysed with RIPA buffer (10 mM Tris-Cl [pH 8.0], 1 mM EDTA, 0.5 mM EGTA), 1% Triton X-100, 0.1% sodium deoxycholate, 0.1% SDS, and 140 mM NaCl, and protein content was determined. RIPA buffer was supplemented with complete protease inhibitor (Roche Diagnostics). Gels (separation gel: 10% acrylamide) were loaded with 10 μg protein and run for 60 min at 20 mA (Bio-Rad, Puchheim, Germany). Gels were blotted at 80 V for 90 min on nitrocellulose membranes (Bio-Rad). Membranes were blocked in 5% milk in TBS for 1 h and incubated with primary antibodies overnight at 4 C.
After washing, membranes were incubated with the detection antibody (goat anti-rabbit IgG HRP, DAKO P0448, 1:2000) for 2 h at room temperature. Proteins complexed with horseradish peroxidase (HRP)-conjugated antibody were stained by addition of ECL Prime (GE Healthcare, Freiburg, Germany) and visualized in a V3 Western Workflow (Bio-Rad).
Intracellular calcium and PoPo3 uptake
Mouse and human B cells were incubated with 4 μM of the calcium dye Cal-520 (ab171868; Abcam, Cambridge, United Kingdom) in HBSS (1 mM Ca2+, #55037C; Sigma) or phosphate-buffered saline (PBS; no Ca2+) with 0.02% Pluoronic F127 (#P6866; Thermo Fisher Scientific, Waltham) for 60 min at 37°C followed by 30 min at room temperature. Cells were washed and resuspended in HBSS or PBS containing 1 μM PoPo3 iodide (#P3584; Thermo Fisher Scientific) with respective antagonists/ligands/inhibitors. After a 30 min incubation at room temperature, cells were transferred into black 96 (200,000 cells/well for mouse samples)- and 384 (35,000 cells/well for human samples)-well plates. A baseline reading was established, CBD was added thereafter, and the intracellular Ca2+ concentration as well as PoPo3 uptake were evaluated with a TECAN multimode reader over 90 min.
Flow cytometry and Annexin V assay (apoptosis)
B cells were analyzed using a MACS Quant 9 analyzer (Miltenyi Biotec, Bergisch Gladbach, Germany). B cell survival and apoptosis were assessed using the FITC Annexin V assay kit (BD Biosciences, Heidelberg, Germany) according to manufacturers' instructions.
Isolation of mouse and human B cells
Mouse B cells were isolated from spleens using CD19 microbeads (#130-121-301; Miltenyi Biotec) according to the suppliers' instructions. Human B cells were isolated from peripheral blood using CD19 microbeads (#130-050-301; Miltenyi Biotec).
Isolation of PBMCs from peripheral blood
PBMCs were isolated using the Greiner LeucoSep Tubes (#227290; Greiner Bio-one, Kremsmünster, Austria) according to manufacturers' instructions.
Animals
In this study, only male mice were used as estrous cycles might cause data variability. Since we investigated expression of TRPA1 in mouse B cells, we excluded female animals due to the influence of estrogen on TRPA1 expression. 9 Male DBA/1J mice (6–8 weeks old) were purchased from Janvier (France). They had unrestricted access to chow and water. Five animals lived in one cage, and they were acclimated to the environment for 1 week before commencement of experiments. The University of Düsseldorf approved all experiments according to institutional and governmental regulations for animal use (Project: O57/15). All procedures were performed in accordance with the German Animal Welfare Act and the European Directive 2010/63/EU on the protection of animals used for scientific purposes.
RASF coculture with PBMC
Coculture experiments were performed in 96-well plates (Cellstar; Greiner bio-one). In brief, 5000 RASF were seeded in 200 μL RPMI-1640 (Sigma-Aldrich) containing 10% FBS and grown for 72 h. Then, 250,000 isolated human PBMCs were added and cells were stimulated with cytokines/CBD as indicated for 7 days. After that, supernatants were collected and cytokine and immunoglobulin production were assessed by enzyme-linked immunosorbent assay (ELISA).
Enzyme-linked immunosorbent assay
ELISAs for interleukin (IL)-6 (#555220), IL-10 (#555157), and tumor necrosis factor (TNF; #555212) were obtained from BD (Franklin Lakes) and were conducted according to the manufacturers' protocol. Immunoglobulin M (IgM) and immunoglobulin G (IgG) were detected by an in-house ELISA. All samples were run in duplicates.
Statistical analysis
Statistical analysis was performed with SPSS 25 (IBM, Armonk). The statistic tests used are given in the figure legends. When data are presented as line plots, the line represents the mean. When data are presented as bar charts, the top of the bar represents the mean, and error bars depict the standard error of the mean. The level of significance was p<0.05.
Results
Mouse and human B cells express TRPV2 and TRPA1
CBD activates several ion channels of the transient receptor potential (TRP) family and it shows the highest affinity toward TRPA1, TRPV1, and TRPV2. 10 We verified the expression of TRPA1 in mouse and human B cells (Fig. 1A, C) and TRPV2 in mouse B cells (Fig. 1B). Densitometric analyses of both proteins in relationship to the reference protein (housekeeper) GAPDH revealed that expression was not modulated when B cells were activated with the toll-like receptor 9 (TLR9) agonist CpG (not shown).
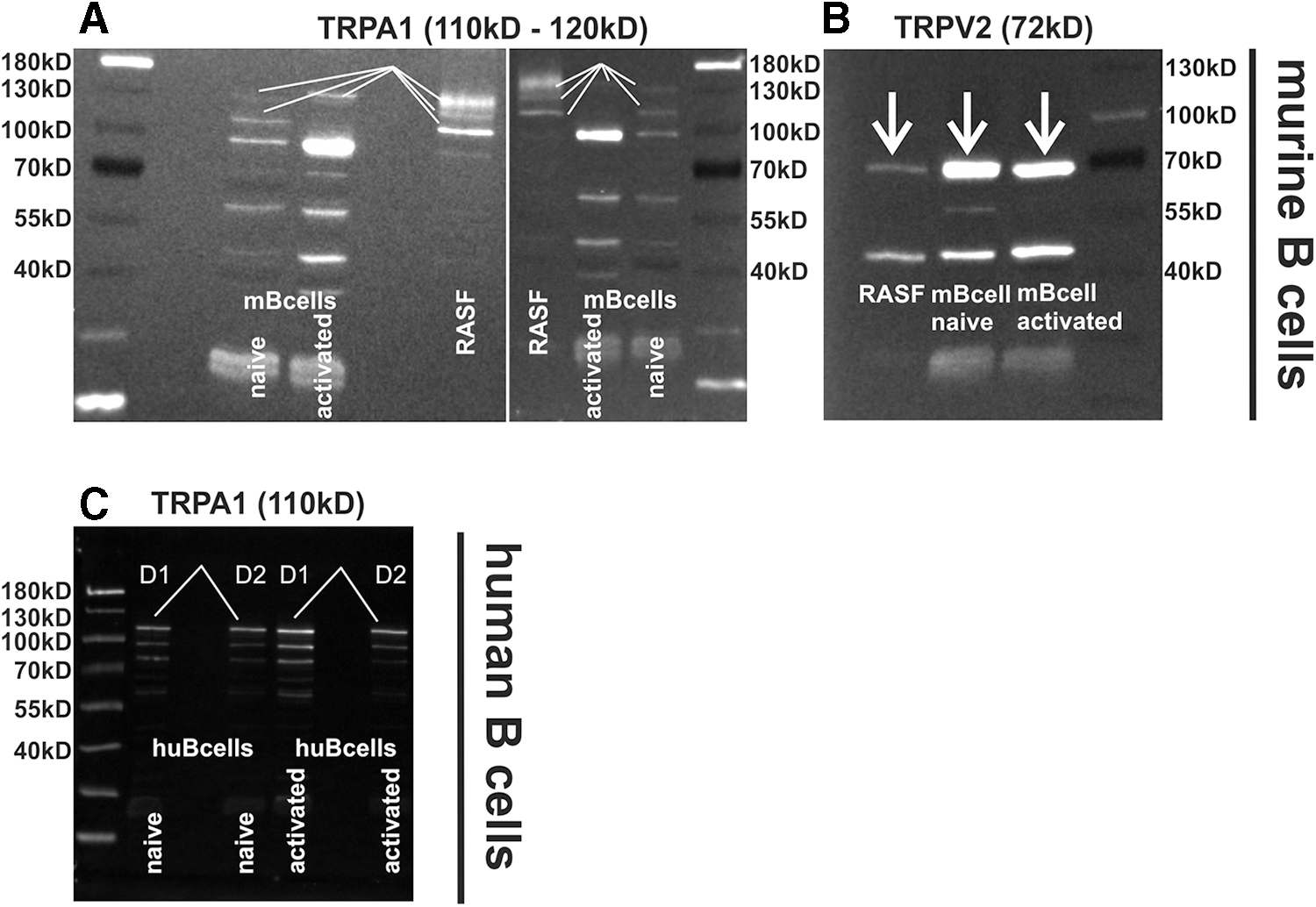
TRPA1 and TRPV2 protein levels in murine and human B cells.
CBD increases intracellular calcium in mouse and human B cells
In previous studies, we found CBD to increase intracellular calcium levels in isolated RASF. 3 Therefore, we investigated the impact of CBD on intracellular calcium levels in activated and nonactivated mouse and activated human B cells. We found that CBD dose-dependently increased intracellular calcium levels in naive and CpG-stimulated mouse B cells (Fig. 2A).
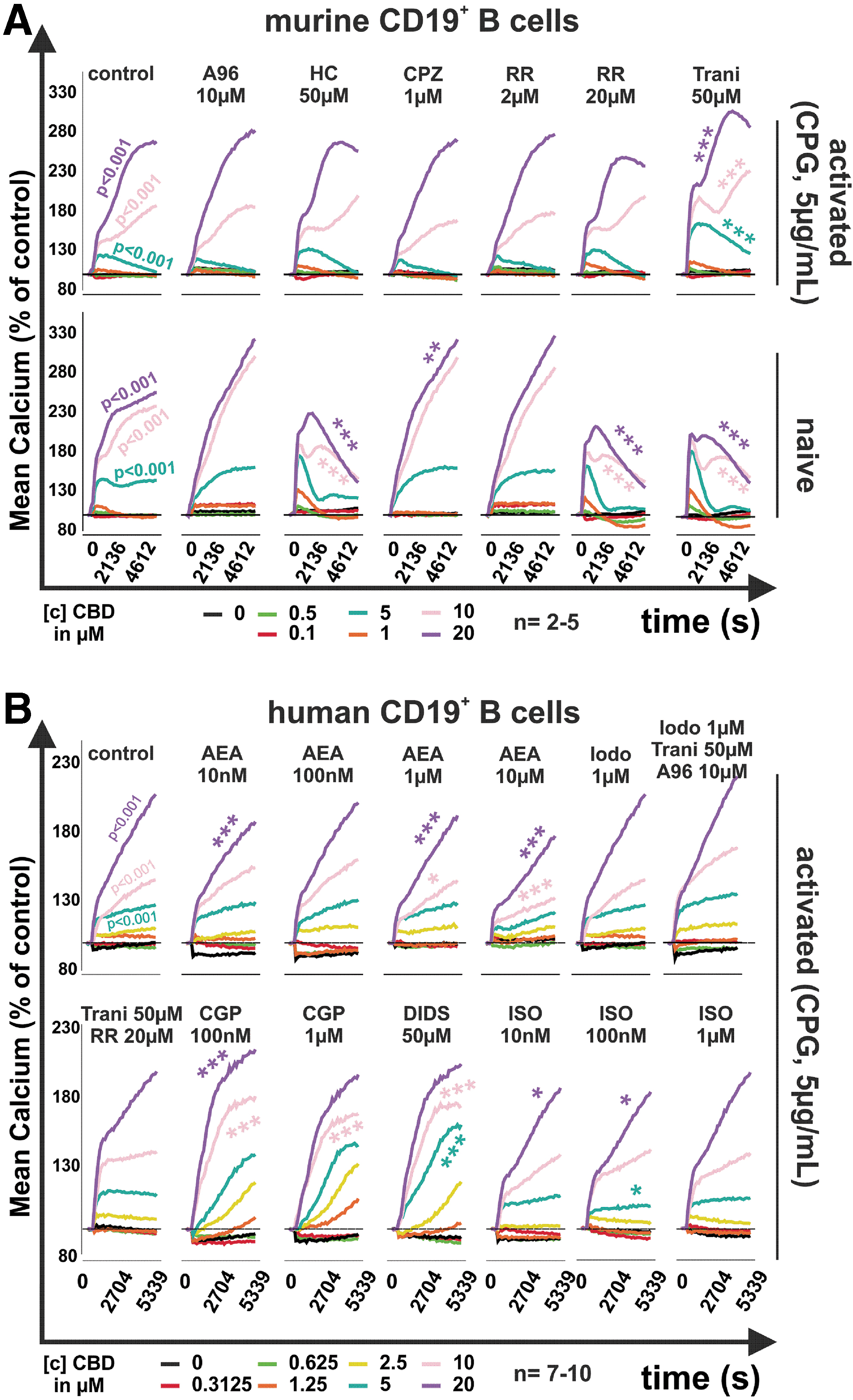
Mean intracellular calcium levels in murine and human B cells in response to CBD.
In naive B cells, the stimulating effect of CBD (10 and 20 μM) on intracellular calcium was reduced not only by the TRPA1 antagonist HC030031 (p<0.001) but also by the pan TRP inhibitor ruthenium red (RR, 20 μM; p<0.001) and the TRPV2 inhibitor tranilast (50 μM; p<0.001). The TRPV1 antagonist capsazepine (CPZ; 10 μM) further increased the effects of CBD at 20 μM. In CpG activated B cells, none of the inhibitors was able to reduce the increase in calcium induced by CBD. In contrast, tranilast further increased calcium levels when combined with CBD (5, 10, and 20 μM; p<0.001).
When analyzing human B cells (Fig. 2B), we focused on CpG-activated B cells, since activated B cells contribute to inflammation and disease activity in RA. 11 In addition, we used several modulators of TRP channels and inhibited mitochondrial proteins that have been described as target molecules for CBD.3,12 Similar to mouse B cells, human B cells increased intracellular calcium in response to CBD (≥5 μM, p<0.001) (Fig. 2B). The endocannabinoid anandamide (AEA) has been described to ligate and desensitize TRPV1 and TRPA1, 13 and therefore, we used this compound to modulate potential effects of CBD at these receptors. At 10 nM, 1 μM, and 10 μM, AEA modestly reduced the increase in calcium induced by CBD (20 μM; p<0.001). Interestingly, neither the combination of the TRPV1 antagonist iodoresiniferatoxin (1 μM), the TRPV2 antagonist tranilast (50 μM), and the TRPA1 antagonist A967079 (10 μM) nor the pan TRP inhibitor RR modulated the effects of CBD.
Since CBD interacts with the mitochondrial proteins voltage-dependent anion-selective channel 1 (VDAC1) and mitochondrial sodium/calcium exchanger (NCLX), we also used 4,4′-diisothiocyanatostilbene-2,2′-disulfonate (DIDS; VDAC inhibitor) and CGP37157 (CGP, NCLX inhibitor). 12 Both compounds further increased CBD-induced intracellular calcium (DIDS 50 μM + 5 μM or 10 μM CBD, p<0.001; CGP 100 nM +10 μM or 20 μM CBD, p<0.001; CGP 1 μM + 10 μM CBD, p<0.001). The unselective agonist of β adrenoreceptors, isoproterenol (ISO), was used since it was shown that norepinephrine is able to reduce the activation of TRPV1 channels, which was reversed by the nonselective β antagonist propranolol. 14 ISO (10 nM to 1 μM) slightly modulated the effects of CBD on intracellular calcium levels, but the magnitude was small, and peak calcium levels were not altered.
CBD enhances PoPo3 uptake in mouse and human B cells
PoPo3, a cationic fluorescent dye, is sold as a live/dead stain similar to propidium iodide that is supposed to enter necrotic cells exclusively. However, we discovered that the uptake of PoPo3 can be independent of cell death and is fostered by the activity of organic cation transporters (OCTs) at the cell membrane. 3 Therefore, PoPo3 can be used as surrogate marker for cellular drug uptake. We used PoPo3 in conjunction with the calcium dye Cal-520 and determined its uptake under the same stimulatory conditions.
In naive mouse B cells, CBD in concentrations above 5 μM significantly increased the uptake of PoPo3 (p<0.001), while in CpG-activated mouse B cells, CBD already elicited PoPo3 uptake at 0.5 μM (p<0.001). Although PoPo3 uptake was modulated positively or negatively by A967079 (10 μM), HC030031 (50 μM), CPZ (10 μM), and RR (2 μM), depending on CBD concentration in naive B cells (Fig. 3A, lower panel), none of these compounds inhibited the effects of CBD. However, RR at 20 μM significantly reduced PoPo3 uptake induced by 20 μM CBD in naive mouse B cells (Fig. 3A, lower panel). In CpG-activated mouse B cells (Fig. 3A upper panel), only CPZ (1 μM) and RR (20 μM) inhibited CBD-induced PoPo3 uptake (p<0.001). The other compounds increased or decreased PoPo3 uptake dependent on CBD concentration, but a clear pattern was absent.
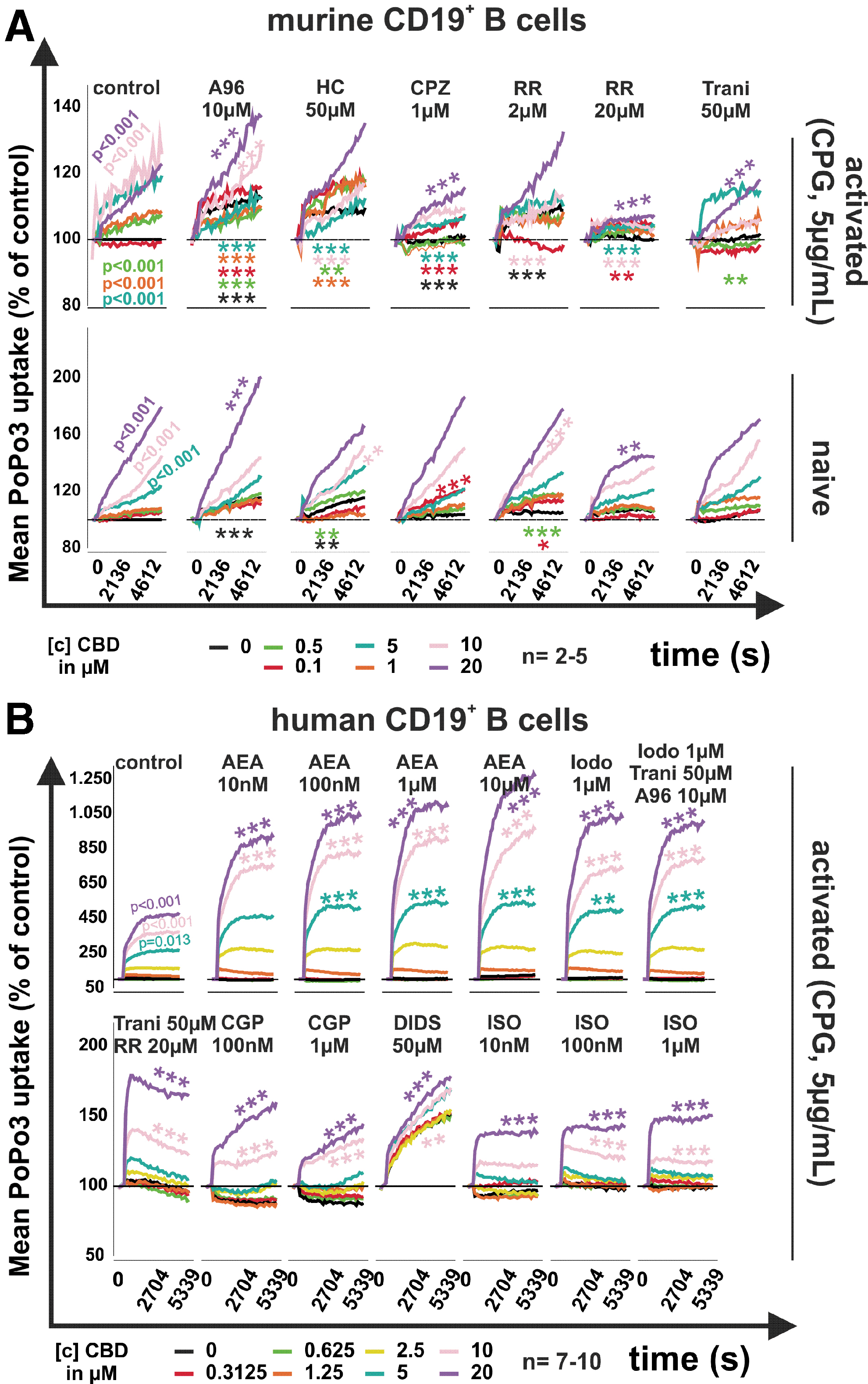
Mean intracellular PoPo3 uptake in murine and human B cells in response to CBD.
In human B cells, PoPo3 uptake was also induced by CBD (p=0.013 at 5 μM and p<0.001 at 10–20 μM), and AEA, iodoresiniferatoxin, and the combination of iodoresiniferatoxin, tranilast, and A967079 further augmented the CBD-induced (10–20 μM) PoPo3 uptake (p<0.001). Interestingly, the combination of tranilast (50 μM) and RR (20 μM), CGP (0.1, 1 μM), DIDS (50 μM), and ISO (10 nM to 1 μM) decreased CBD-induced [10 and 20 μM]) PoPo3 uptake (p<0.001, except for ISO [10 nM at CBD 10 μM]).
CBD reduces survival, IL-10, and TNF production of mouse B cells
CBD has been shown to decrease the viability of several cell types, including RASF, 3 and therefore, we investigated the impact of this phytocannabinoid on apoptosis of mouse B cells using annexin V staining followed by flow cytometry. CBD reduced the number of living cells from 16%±13% to 5.5%±4.7% at 5 μM CBD (p=0.013), to 6.2%±5.9% at 10 μM CBD (p=0.021), and to 5.5%±5.6% at 20 μM (CBD (p=0.011) (Fig. 4A). This was not inhibited by either the TRPA1 antagonist A967079 (10 μM) or the TRPV1 antagonist CPZ (10 μM).

Survival, IL-10, and TNF production of murine B cells in response to CBD.
In contrast, early apoptotic B cell numbers increased from 64.3%±12.7% to 73.2%±6.5% at 5 μM CBD (p=0.031), to 74.2%±6.7% at 10 μM (p=0.031), and to 76.2%±8.0% at 20 μM CBD (p=0.008) (Fig. 4B). The number of late apoptotic and necrotic cells was unchanged by CBD (Fig. 4C, D).
Besides its influence on cell viability, CBD has been shown to modulate cytokine production. We assessed anti-inflammatory IL-10 and proinflammatory TNF production induced by CpG. In concentrations above 5 μM, CBD completely abrogated both IL-10 and TNF production (p<0.001) by CpG stimulated mouse B cells (Fig. 4E, F). In the absence of CBD, IL-10 production was 98±26 pg/mL and TNF production was 298±348 pg/mL. In addition, CPZ alone decreased IL-10 and TNF production significantly (Fig. 4E, F; p<0.001), while A967079 alone decreased TNF production only (Fig. 4F; p=0.012).
CBD modulates IL-10 and TNF production in PBMCs and PBMC/RASF coculture
We already investigated the impact of CBD on isolated RASF and B cells, but to better mimic the environment in synovial tissue, we determined the influence of CBD on PBMCs and PBMC/RASF cocultures. In PBMC monoculture and PBMC/RASF coculture, CBD did not modulate IL-6 production regardless of stimulation (Fig. 5A, D). Of note, IL-6 production in coculture was 100-fold higher than in PBMCs alone, which suggests that RASF are the main producer of this cytokine.

Cytokine production by human PBMC monoculture and coculture with RASF in response to CBD.
IL-10 release was increased from 108±35 to 279±86 pg/mL (p<0.001) by the addition of anti-IgM and to 227±81 pg/mL (p<0.001) by the addition of CpG in coculture without the addition of CBD (Fig. 5B). Similarly, in PBMC monoculture, anti-IgM and CpG increased IL-10 production from 97±56 to 246±142 pg/mL (p=0.003) and to 410±388 pg/mL (p<0.001), respectively (Fig. 5E). In coculture, 10 μM CBD increased the anti-IgM-induced IL-10 production to 536±269 pg/mL (p=0.005), but decreased IL-10 production from 96±36 to 65±33 pg/mL (p=0.034) when cells were stimulated with interferon-gamma (IFN-γ) (Fig. 5B). In PBMC monoculture, CpG-induced IL-10 production was reduced from 410±388 to 136±49 pg/mL (p<0.001) by 10 μM CBD (Fig. 5E).
In contrast, anti-IgM-induced IL-10 production was increased from 246±142 to 349±52 pg/mL (p=0.002) (Fig. 5E). In addition to IL-10, we also determined TNF production, since it is one major cytokine in the pathogenesis of RA and target of biological therapy. 15 In coculture without CBD, TNF secretion was enhanced by anti-IgM treatment from 57±25 to 105±46 pg/mL (p=0.006), while in monoculture TNF was increased by anti-IgM from 115±149 to 580±444 pg/mL (p=0.001) and by IFN-γ to 308±257 pg/mL (p=0.02) (Fig. 5C, F).
Overall, CBD had an inhibitory effect on TNF production. In coculture, CBD (1 μM) reduced TNF production by CpG from 89±45 to 37±12 pg/mL (p=0.004) and by IFN-γ from 85±39 to 38±8 pg/mL (p=0.013) (Fig. 5C). In line with this, 10 μM CBD reduced TNF levels when untreated from 57±25 to 31±9 pg/mL (p=0.031) (Fig. 5C). In PBMC monoculture, 1 μM CBD reduced CpG- and IFN-γ-induced TNF production from 146±103 to 56±22 pg/mL (p=0.010) and from 308±257 to 71±49 pg/mL (p=0.033), respectively. At 10 μM CBD, CpG- and IFN-γ-induced TNF production was reduced to 50±21 pg/mL (p=0.002) and to 67±63 pg/mL (p=0.011), respectively (Fig. 5F).
CBD: influence on immunoglobulin (IgM and IgG) production
The impact of CBD on immunoglobulin production has only been addressed superficially. In murine models, Jan et al and Kaplan et al found CBD to reduce antibody production by reducing T cell activation and proliferation.16,17 In this study, we show whether CBD influences differentially activated human PBMCs alone or in combination with RASF in respect to antibody production. In coculture, IgM and IgG (IgG) production were significantly induced when CBD was combined with CpG, a TLR9 agonist used to activate B cells independently of T cells (Fig. 6A, C). Under these conditions, 10 μM CBD increased CpG-induced IgM production from 399±42 to 1123±363 ng/mL (p=0.041), respectively (Fig. 6A).

IgM and IgG production in PBMC monoculture and coculture with RASF in response to CBD.
Of note, 1 μM CBD also enhanced basal IgM production from 80±31 to 328±232 ng/mL (p=0.049) (Fig. 6A). IgG production was similarly modulated by CBD in coculture. CBD increased IgG levels under CpG (from 403±346 to 909±162 ng/mL at 10 μM CBD, p=0.041), and without further stimulation (p=0.049; only significant in Kruskal–Wallis test but not in Bonferroni post hoc analysis) (from 465±279 to 776±54 ng/mL [(1 μM CBD], and to 784±49 ng/mL [10 μM CBD]) (Fig. 6C).
Discussion
In this study, we showed that CBD influences intracellular calcium levels, compound uptake, survival, cytokine, and immunoglobulin production of mouse and human B cells. These effects were independent of TRPA1, TRPV1, or TRPV2 activation although CBD was described as an agonist for these ion channels.3,10,18,19 In addition, we found that CBD modulates IL-10 and TNF production by PBMCs alone or in coculture with RASF.
In a first approach, we determined the expression of potential target receptors in mouse and human B cells. We focused on TRPA1 and TRPV2 channels since we and others have shown their involvement in RA pathology.3,20 Whether TRPV1 is also expressed in B cells is still unclear, but results from Soutar et al suggest that its involvement in regulating B cell function might be negligible since piperine, an agonist at TRPA1 and TRPV1 channels, influenced B cell cytokine and IgG production also in TRPV1 knockout mice suggesting that its target is TRPA1 rather than TRPV1. 21 To our knowledge, we are the first group to reveal TRPA1 protein expression in mouse and human B cells. Mouse B cells, however, showed very low expression of TRPA1 compared with their human counterpart.
Of note, we detected up to four bands at the expected size for TRPA1, although the lowest band at around 100 kDa is not TRPA1 but another cross-reactive protein detected by the antibody used in our study. 22 The bands above 100 kDa are specific for TRPA1 as they reflect differentially glycosylated variants of TRPA1. 23 The presence of TRPV2 protein has already been demonstrated in human B cells, 24 and therefore, we only investigated its expression in mouse B cells, where this ion channel was robustly expressed.
Since TRPV2 and TRPA1 are involved in the regulation of cellular calcium levels, we assessed the effects of CBD, an agonist at both receptors, on intracellular calcium concentrations. We found CBD to increase intracellular calcium levels in naive and CpG-activated mouse and in CpG-activated human B cells. While TRP channels were involved in regulating calcium in naive mouse B cells, activated mouse B cells were unresponsive to general TRP or specific TRPA1 inhibition. In accordance with this, calcium levels in activated human B cells were also not reduced by TRPA1 antagonists, but were negatively modulated by the endocannabinoid AEA and enhanced by targeting the mitochondrial proteins voltage-dependent anion-selective channel 1 (VDAC1) with DIDS and the mitochondrial NCLX with CGP37157 (CGP).
CBD has been identified as a ligand for several mitochondrial proteins such as VDAC1 and NCLX,3,12,25 and it has been shown that CBD induces calcium overload with concomitant opening of the membrane permeability transition pore and cell death.3,12 DIDS further increased the CBD-induced elevation of intracellular calcium levels since both compounds inhibit VDAC1, resulting in reduced mitochondrial calcium uptake and therefore increased cytosolic calcium levels. This was also demonstrated in VDAC1-silenced HeLa cells that showed a 20% increase in cytosolic calcium upon VDAC1 knockdown. 26 CGP is an inhibitor of NCLX, preventing calcium efflux from mitochondria, but it also inhibits voltage-gated calcium channels at the plasma membrane, thereby reducing calcium entry from the extracellular space. 27 CGP did not reduce the effects of CBD but further augmented CBD-induced intracellular calcium levels, suggesting that CBD and CGP do not share a common cellular target.
We also used AEA to inhibit the effects of CBD since it shares many of CBDs cellular targets and is able to desensitize TRP channels. 28 AEA slightly reduced the effects of CBD on intracellular calcium, although we were unable to determine the proteins involved. However, ISO, an unselective beta adrenergic agonist also reduced CBD-induced calcium elevations, and since beta adrenergic receptors are able to decrease TRPV1-mediated responses, 14 AEA might also target TRP channels to modulate the effects of CBD.
In addition to intracellular calcium, we also determined the CBD-induced uptake of the fluorescent dye PoPo3, which we recently identified as a surrogate molecule for drug uptake. 3 In mouse and human B cells (activated and naive), CBD induced PoPo3 uptake, which was modulated positively or negatively, but was not inhibited by TRPA1 antagonists. CPZ and RR reduced CBD-induced PoPo3 uptake in activated mouse B cells, while in naive mouse B cells, only RR showed some effect. In human B cells, AEA, the TRPV1 antagonist iodoresiniferatoxin and the combination of TRPV1/TRPA1/TRPV2 antagonists enhanced CBD-induced PoPo3 uptake, while CGP, DIDS, RR, and ISO reduced the uptake.
These results seem random, but in a previous study, we found that OCTs are the most likely proteins that mediate PoPo3 uptake 3 and, for example, ISO has been identified as an inhibitor of OCT.29,30 Similarly, other compounds used in this study might enhance or diminish the activity of OCTs and thereby modulate the effects of CBD on PoPo3 uptake. PoPo3 uptake experiments revealed another important feature of CBD: It fosters the uptake of compounds into B cells. This might be used to shuttle therapeutic molecules like saracatinib, whose uptake is dependent on the human novel OCT1, 31 more efficiently into B cells.
Since intracellular calcium and cell death are connected, 32 we also assessed the ability of CBD to induce apoptosis and necrosis in mouse B cells and found that CBD reduced the number of living B cells while concomitantly increasing the number of early apoptotic cells. Although CBD has been described as antiapoptotic in several studies,33–35 others have identified proapoptotic pathways that are engaged by CBD.12,36,37 Our results are in line with the proapoptotic effects of CBD, and we also observed that high concentrations of CBD are necessary to reduce cell viability. 38 Activated B cells are a source of the proinflammatory cytokine TNF, 39 but they also produce anti-inflammatory IL-10, 40 and, therefore, we investigated the production of these cytokines in response to CBD.
Concentrations equal or greater than 5 μM CBD completely inhibited TNF and IL-10 production in mouse B cells and this is in line with previous results where we demonstrated a strong inhibitory effect of CBD on SF IL-6, IL-8, and MMP-3 production. 3 In addition, CBD also reduced cytokine levels in other cell types. 38 Although IL-10 was inhibited by CBD, which per se would be considered proinflammatory, the impact on TNF might be more relevant since mouse B cells stimulated with CpG produced around 3.5 times more of this proinflammatory cytokine compared to IL-10. Of note, the TRPV1 antagonist CPZ did not inhibit the effects of CBD, but had significant own effects independent of CBD. This might be an indicator that TRPV1 is expressed and functional in B cells, but it might also be an effect of TRPA1 since CPZ was identified as an agonist at this ion channel. 41
Besides B cells, we also analyzed the impact of CBD on PBMC and PBMC/RASF coculture cytokine and immunoglobulin production under differential stimulating conditions. IL-6 production was not modulated by CBD in PBMC and PBMC/RASF coculture. In previous studies, we detected an influence of CBD on RASF IL-6 production, but this was only evident with low FBS concentrations. 3 Interestingly, as in mouse B cell monoculture, in PBMC cultured alone, IL-10 production was induced by the TLR9 ligand CpG and decreased by CBD. However, when PBMC were cocultured with RASF, CpG also induced IL-10, but we did not observe an inhibitory effect of concomitant CBD stimulation. In addition, CBD reduced CpG-induced TNF production in PBMC/RASF coculture. CpG is an agonist at TLR9 and this pattern recognition receptor is only expressed by plasmacytoid dendritic cells (pDCs) and B cells in peripheral blood.42,43
Since pDCs only account for around 0.11% in peripheral blood 44 while B cells account for 5–10% of all PBMCs, 45 it is likely that B cells are the major target of CpG and main contributor to IL-10 production under these stimulatory conditions. CpG is considered as a T cell independent B cell activation stimulus, 46 and therefore, we also used anti-IgM as a polyclonal activator of B cell receptor signaling. Since T cells were present in the cell preparation, anti-IgM stimulation mimicked an antigen-specific activation of B cells. Under these conditions, IL-10 and TNF production was upregulated in PBMC and PBMC/RASF cocultures independent of CBD. However, in contrast to CpG, CBD enhanced the anti-IgM-induced IL-10 production in mono and coculture without regulating TNF levels.
These results demonstrate that CBD influences IL-10 and TNF production dependent on activation stimulus (antigen specific or unspecific) and whether cultured alone or together with RASF. Of note, we also used IFN-γ as an activation stimulus for PBMC and PBMC/RASF coculture. IFN-γ induces major histocompatibility complex class II expression on RASF, 47 and since we did not match HLA haplotypes, an allogeneic reaction between RASF and T cells might occur, which in turn would activate T cells. However, without CBD, IFN-γ did not modulate cytokine production compared to control, suggesting limited, if any, T cell activation.
RA is characterized by autoantibody production and, therefore, we investigated the impact of CBD on IgM and IgG production. While IgM and IgG production was not modulated by CBD, IFN-γ, CpG, or anti-IgM in PBMC alone, CBD enhanced the CpG- and anti-IgM-induced production of IgM and IgG in coculture. In contrast to our findings, it has been shown in vivo that CBD reduces the production of antigen-specific immunoglobulin. 16 However, in this aforementioned study, splenocytes from ovalbumin treated mice were investigated and, in this setting, activation of B cells and T cells was antigen specific.
In our study, we used CpG activation of B cells, which does not require T cell help and which induces the production of natural antibodies protecting against bacterial infections. 48 It is therefore possible that CBD supports less specific IgM production but inhibits specific IgG production by interfering with T cell help and, in fact, it has been shown that CBD reduces TH1 cytokine production. 49
Concluding Remarks
In this study, we investigated for the first time the impact of CBD on B cell function. CBD not only decreases B cell viability, cytokine, and antibody production but it also enhances drug uptake. These data provide evidence that CBD can be used as adjunct therapy in RA. Therefore, we propose a combination of CBD with common antirheumatic drugs and this might provide several advantages over monotherapies such as enhanced drug uptake (and thereby lowering the necessary therapeutic concentrations and possible side effects) and additional or even synergistic effects on cytokine and antibody production.
Ethics Approval
This study was approved by the local Ethics Committee of Düsseldorf (2018-87-KFogU and 2018-96-KFogU).
Footnotes
Acknowledgment
We thank Birgit Opgenoorth for excellent technical assistance.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by an unlimited grant of the Hiller Foundation.