
Abstract
Objective:
Osteoarthritis (OA) is common degenerative joint disease, mostly characterized by gradual cartilage breakdown. Currently there are no disease-modifying drugs available, therefore, there is an increasing need for basic research to focus on cartilage function in OA. Changes in cannabinoid receptor 2 (CB2) expression were observed in the OA-affected joints, although its action on cartilage chondrocytes remain unclear. We studied the action of dimethylbutyl-deoxy-delta-8-THC (JWH-133), selective CB2 agonist, on chondrocytes metabolism using both in vitro and in vivo studies.
Design:
Intraarticular (i.a.) injection of monoiodoacetate (MIA) was used to induce OA in rats. OA-related pain symptoms were assessed by pressure application measurements (PAMs). Primary human chondrocytes treated with MIA were used to investigate action of JWH-133 on chondrocytes viability, proliferation, and motility. Cannabinoid system components, inflammatory cytokines and metalloproteinases (MMPs) expression was measured on messenger RNA and protein levels in chondrocytes and animal cartilage.
Results:
Repeated, i.a. administration of JWH-133 showed antinociceptive potential in PAM, as well as decreased levels of MMPs, which suggests that CB2 agonism may modify degradation of cartilage. JWH-133 administration partially reduced toxicity, increased proliferation, and chondrocytes' migration. Moreover, our data suggest that CB2 agonism leads to alleviation of MMPs expression both in vitro and in vivo.
Conclusions:
In this study, we demonstrate modifying effect of JWH-133 local administration on cartilage metabolism and MMP13 expression that was shown to be involved in cartilage degradation. CB2 receptors' activation may be of benefit for chondrocytes' proliferation, therefore delaying disease progression. Our results propose direction of studies on OA-modifying treatment that can benefit in management of human OA.
Introduction
Osteoarthritis (OA) represents the most common condition that leads to joint pain and is characterized by a loss of joint cartilage, which occurs due to an imbalance between anabolic and catabolic processes in chondrocytes. 1 In the course of the disease, subchondral bone become exposed, causing nerve irritation and pain during joint loading. Currently, there are no therapeutics available that can modify disease development, and pharmacological therapy focuses on alleviation of OA pain symptoms.2–4 Regardless of treatment, the disease progresses or even gets promoted, facilitating a strong need to develop treatment strategies that affect the development of OA.
For many years, cannabinoids, which are derived from Cannabis sativa, have found therapeutic applications because of their anti-inflammatory and analgesics properties. Cannabinoids produce these effects by activating cannabinoid receptors, which are classically divided into two types, namely cannabinoid receptor 1 (CB1) and cannabinoid receptor 2 (CB2). Both are expressed in different types of cells in joints that are involved in OA development, including subchondral osteoclasts, synovial fibroblasts, and chondrocytes. 5 Our previous studies uncovered changes in cannabinoid receptor expression in OA-affected joints. 6 Moreover, recent studies suggest that the activation of CB receptors may be beneficial in the treatment of different types of arthritis, highlighting the system as an interesting target in OA research. 7
Our studies focused on the CB2 receptor, as it is expressed in peripheral neuronal tissue and in cartilage, making it a candidate for possible synergistic therapy for both OA symptoms and disease reversal. The activation of this receptor leads to reduced inflammation and a decrease in pain symptoms in different types of OA. 8 Furthermore, studies in a CB2-deficient animal model of OA have shown the involvement of this receptor in cartilage degradation, suggesting its importance in chondroprotection. 9 The expression of CB2 is also higher in chondrocytes than in osteoblasts, indicating that chondrocytes may be more responsive than osteoblasts to pharmacological treatment. 10 Moreover, the relationship between the cannabinoid system and proteolytic enzymes is interesting in the aspect of diseases whose pathomechanism is based on tissue degeneration. 11
In the case of OA, the enzymes that can respond to the action of cannabinoids may be metalloproteinases (MMPs) and others related to the degradation of the joint cartilage. 12 There is a lack of research connecting CB2 pathway with cell proliferation and motility that influence cartilage regeneration. Studies showed that cartilage oligomeric matrix protein (COMP), tissue inhibitors of MMP (TIMP1), and MMPs are potential indicators of OA progression. 13 Moreover, those enzymes are strictly connected to cell migration in vivo.14–17
Therefore in our studies, we looked for the relationship between these enzymes and the activation of the CB2 receptor. The problem with plant-derived cannabinoids is lack of selectivity; therefore, an increasing number of studies use chemical analogs to prove the involvement of certain cannabinoid receptors in the examined processes. 11 In the present study, we used dimethylbutyl-deoxy-delta-8-THC (JWH-133), a potent CB2 agonist, in a rat model of OA after chronic intraarticular (i.a.) injections. To induce OA-related changes in animal joints, we use a model of a single injection of monoiodoacetate (MIA; 1 mg). After performing behavioral studies, we collected cartilage tissue to analyze the level of MMPs and components of the endocannabinoid system (ECS).
To discover the mechanism of action of JWH-133, further studies used cultures of human chondrocytes (HC) treated with MIA. We performed messenger RNA expression analysis of selected components of ECS, as well as proinflammatory cytokines to assess the nature of changes induced by MIA. Furthermore, we examined the effect of MIA and JWH-133 on HC viability and proliferation. Finally, through scratch-wound assays, we assessed the involvement of CB2 agonism on the proliferation and motility of chondrocytes.
Materials and Methods
Cell cultures and treatments
Experiments were performed on HC (Cat. No. 402K-05a; Cell Applications). Cells were cultured under standard conditions (37°C, 5% CO2 saturation), dedicated media purchased from Cell Applications (Cat. No. 410–500) supplemented with 10% fetal bovine serum and antibiotics (50 U/mL of penicillin and 50 μg/mL of streptomycin). Cell line used in the study is a primary culture; therefore, experiments were performed at p4–p9. Cells before experiments were cultured at least 24 h without antibiotics. Cells were treated with MIA (3 μM, based on separate set of experiments; Supplementary Figs. S1A and S2A), JWH-133 (10 μM, based on separate set of experiments; Supplementary Fig. S1B), and/or 6-Iodopravadoline (AM-630) (10 μM, based on separate set of experiments; Supplementary Fig. S1C).
Animals and induction of OA model
Male Wistar rats (Charles River, Hamburg, Germany) initially weighing between 225 and 250 g were used for all the experiments. The rats were housed in groups of 5 animals per cage under a 12:12 h light/dark cycle and had free access to food and water. All the animals could acclimatize to their holding cages for 3–4 days before any behavioral or surgical procedures were conducted. The rats were deeply anesthetized with 5% isoflurane (Forane®; Baxter Healthcare Corporation) and 27-gauge needle was introduced into the joint cavity through the patellar ligament, and 50 μL containing 1 mg MIA in 0.9% saline was injected i.a. to induce OA-like lesions. The MIA model was chosen due to its usefulness for studying joint pain and multiple similarities in the mechanisms of cartilage degeneration to those observed in human OA cartilage. 18
The experiments were performed following the guidelines of the International Association for the Study of Pain and with the approval no. 938/2012 of the Local Bioethics Committee of the Maj Institute of Pharmacology (Cracow, Poland). Care was taken to implement the “3 R” rule (replacement, reduction, and refinement) to reduce the number of animals used and their suffering during the experiments. Altogether, data obtained from 58 animals were presented in the study—to see exact number of animals in each experiment; Figure legends.
Drugs and reagents
MIA was obtained from Sigma-Aldrich (Darmstadt, Germany), AM-630, and JWH-133 from Tocris Bioscience (Bristol, UK). All the reagents were dissolved in a vehicle solution. The vehicle for the compounds used in in vitro studies was culture medium with 0.5% dimethyl sulfoxide (DMSO). As the concentration of JWH-133 used in behavioral studies was higher than compound solubility in 0.9% saline (due to small volume of i.a. injections, 50 μL), 10% Cremophore was added to the solvent. We chose not to use DMSO in animal studies due to the possible effects of this compound on articular cartilage cells during long-term exposure.
Pressure application measurement
The pressure application measurement (PAM) device (PAM; UgoBasile, Italy) has been used for the mechanical stimulation and assessment of joint pain. A quantifiable force was applied for direct stimulation of the joint, and the automatic readout of the response was recorded as described previously. 19 The peak gram force applied immediately before the limb base unit recorded withdrawal, and this value was designated as the limb withdrawal threshold (LWT). Two measurements were performed and mean LWT was calculated for each animal. The experimenter was blinded to the treatments' group.
Treatment paradigm
In an initial behavioral study, pain symptoms were measured every 2nd day starting at day 0 (induction of the model) till day 28 (the timing of the study was dictated by approval of the Ethics Committee for animal testing). In subsequent behavioral studies, responses to stimuli were measured starting 14 days after MIA injection (after pain phenotype was fully developed) and finished by the day 28. JWH-133 was administered every 2nd day at the dose of 100 ng per injection. The dose was chosen based on separate set of experiments (Supplementary Fig. S4). Changes in joint hypersensitivity were compared to the baseline control (osteoarthritic rats treated with vehicle). Cartilage was collected at day 28. Treatment paradigm is presented in Figure 1.
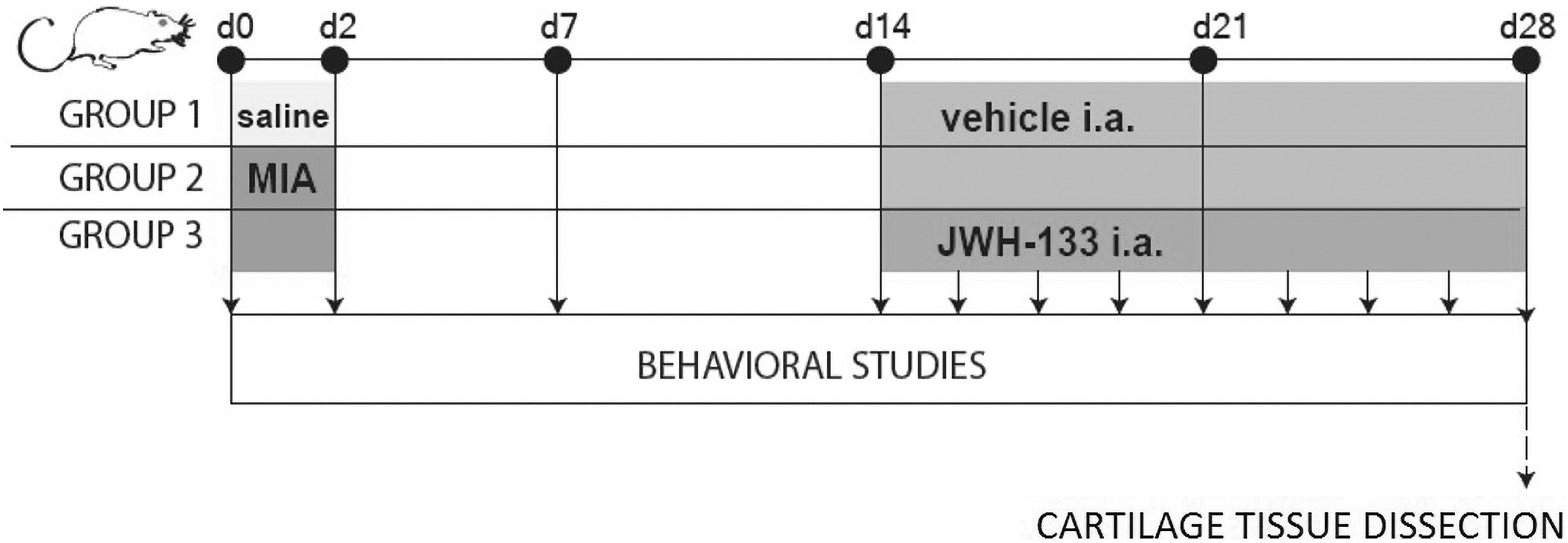
Experimental design. Animals were monitored for OA-related pain symptoms using PAM before MIA injection and every 2nd day after the procedure till the day 28. Chronic intraarticular treatment with JWH-133 (100 ng) was performed during chronic pain state (starting at day 14). Analysis of protein expression was conducted on joint tissue samples collected at the end of experiment. One symbol stands for p<0.05, two symbols stand for p<0.01, three symbols stand for p<0.001. i.a., intraarticular; JWH-133, dimethylbutyl-deoxy-delta-8-THC; MIA, monoiodoacetate; OA, osteoarthritis; PAM, pressure application measurement.
Biochemical assays
Compounds cytotoxicity was measured by the mean of Lactate Dehydrogenase (LDH) Cytotoxicity Detection Kit (Roche). LDH is stable cytoplasmic enzyme present in all cells, released to cell medium during plasma membrane damage. Cells at confluency 80–90% were treated with compounds of interest. Twenty-four hours after stimulation medium from cells was collected, placed on 96-well plates and tetrazolium salt was added for 30 min. Samples' absorbance was measured at a wavelength of λ=492 using a spectrophotometer (Multiscan, Labsystem). Results were expressed as a percentage of the absorbance in negative control cells (cells treated with vehicle). To assess the effect of test compounds on proliferation in cell cultures in vitro, 5-bromo-2’-deoxy-uridine (BrdU) incorporation assay was performed.
Detection of BrdU in proliferating cells is based on color response, and color intensity and absorbance value correlate directly with the amount of DNA containing BrdU molecules, which is related to the number of proliferating cells. Cells at confluency around 50% were treated with compounds of interest in BrdU assay. The test was performed 24 h after cells treatment. Cells were incubated with BrdU for 24 h at 37°C, 5% CO2, followed by anti-BrdU-peroxidase antibody for 90 min. Colorimetric measurements at λ=370 were made using a spectrophotometer.
RNA preparation and quantitative real-time PCR
Cell samples were collected in TRIzol Reagent (Invitrogen, Carlsbad, CA) and homogenized by pipetting and vortexing. RNA was isolated according to Chomczynski's method. 20 The total RNA concentration was measured using a NanoDrop ND-1000 Spectrometer (Nano-Drop Technologies, Wilmington, DE). Reverse transcription of total RNA (500 ng per sample) was performed using iScript reverse transcriptase (Bio-Rad, Hercules, CA) according to protocol.
Quantitative PCR (qPCR) reactions were performed using SYBR Green dye and matching primers: 5′-TGGCAGCGTGACTATGACCTT-3′ and 3′-CCACGGGTGAGCAGAGCTT-5′ (Cnr2), 5′-CACTCACAGACCTGACTCGGTT-3′ and 3′-AAGCAGGATCACAGTTGGCTGG-5′ (Mmp3), 5′-GCCACTACTGTGCCTTTGAGTC-3′ and 3′-CCCTCAGAGAATCGCCAGTACT-5′ (Mmp9), 5′-CCTTGATGCCATTACCAGTCTCC-3′ and 3′-AAACAGCTCCGCATCAACCTGC-5′ (Mmp13), 5′-GAGTAGTGAGGAACAAGCCAG-3′ and 3′-TGCATCTAGATTCTTTGCCTTTT-5′ (Il6), 5′-CAGCCATACAGCAAATCCTTG-3′ and 3′-ATTCCGGTGTTGAGCAGTTT-5′ (Cox2), 5′-GGAGATGCTTGTGACAGCGATC-3′ and 5′-TGAGTCCTCCTGGGCACTGTTA-3′ (Comp), 5′-GGAGAGTGTCTGCGGATACTTC-3′ and 5′-GCAGGTAGTGATGTGCAAGAGTC-3′ (Timp1), 5′-GATGAGTATGCCTGCCGTGTG-3′, and 3′-CAATCCAAATGCGGCATCT-5′ (B2m).
Reactions were run on a Real-Time PCR iCycler IQ (Bio-Rad) with the 3.0 software version. Cycle threshold values (Ct) were calculated automatically. Expression of the B2m was quantified to control for variation in complementary DNA amounts. The abundance of RNA was calculated as 2−(normalized threshold cycle).
Protein isolation and western blot analysis
For proteomic assessment, joint tissue samples of ipsilateral knees were collected. Cartilage samples were frozen in liquid nitrogen and stored at −80°C until protein isolation. Isolation and western blot analysis was performed as described before. 12 Used antibodies are detailed in Table 1. Glyceraldehyde-3-phosphate dehydrogenase was used as a reference protein in studies.
List of Antibodies Used for Western Blot Experiments
CB1, cannabinoid receptor 1; CB2, cannabinoid receptor 2; COMP, cartilage oligomeric matrix protein; FAAH, fatty acid amide hydrolase; HRP, Horseradish peroxidase; IgG, immunoglobulin G; MMP, metalloproteinase; TIMP1, tissue inhibitors of MMP.
Scratch healing assay: collective cell migration assay
The plated HC were grown to confluence on noncoated surface. The cells were incubated with the tested compounds and after 24 h, a scratch (so-called wound) was made in the cell layer with the tip of an automatic pipette. The cells prepared in this way were placed in an Olympus CK-40 inverted microscope chamber coupled with a video microscopy system, where they were recorded for 24 h. A 48-h experimental system was used to observe changes using the lowest possible dose of the MIA compound that did not show toxic effect in HC cells (for details see Supplementary Fig. S1). The wound area after 24 h was compared to the original wound area and the percentage of overgrowth was calculated. The experiment was performed three times, and the data were collected from at least 24 reading frames for each experimental setup.
Statistical analysis
All statistical analyses were performed using Prism V.5 (GraphPad Software, La Jolla, CA). The results are plotted as the mean±standard error of the mean. Differences in the behavioral tests (PAM test, n=8–10), transcriptomic (qPCR assay, n=4–6), and proteomic (western blot analysis, n=6–8) results between the groups were assessed by one-way analysis of variance followed by Bonferroni post hoc test. The number of animals was minimized to comply with 3R policy, but with respect to preserving statistical significance. Cell culture experiments were conducted in three biological repetitions, each at different passage level. The threshold of significance was set at p<0.05. (*) denotes significant differences between sham-operated animals and cells treated with vehicle; (#) denotes MIA-treated animals or cells.
Results
Chronic JWH-133 i.a. injections increased withdrawal threshold in the chronic pain state of MIA-treated animals
We used the PAM test to assess pain-like behavior in osteoarthritic rats. Our results revealed significant increases in joint hypersensitivity after MIA injection that was characterized by a biphasic profile of pain-like behavior (Fig. 2A). An immediate development of pain behavior lasted for 6 days, followed by an alleviation of symptoms from days 8 to 12, then further progressive, chronic pain development from day 14 until the experimental endpoint. As described previously, the first pain phase relates to on-site inflammation, while the second phase develops due to cartilage degeneration and exposure of nerve endings within the affected joint. 21 Sham-treated animals (saline injection) experienced development of pain related to the procedure but did not develop chronic pain behavior as MIA-treated group did (Supplementary Fig. S3).

PAM after MIA injection (1 mg)
To observe the effect of CB2 agonism on cartilage condition in OA, the treatment paradigm, consisting of 3 doses of JHW-133 (100 ng per knee joint), began at day 14 and continued until day 28 (Fig. 2B). We observed alleviation of pain symptoms at day 20 after MIA administration. This dose of JWH-133 was optimized in separate studies (Supplementary Fig. S4). The treatment with the CB2 agonist was effective until the end of the behavioral experiment, showing a significant increase in withdrawal threshold in the JWH-133-treated group of animals. JWH-133 showed no effect in sham-operated animals (data not shown).
Protein expression of CB2, selected MMPs, and related proteins in OA-affected cartilage is affected by treatment with JWH-133
Both cannabinoid receptors, CB1 and CB2, were examined in cartilage samples collected at day 28 from animals subjected to JWH-133 treatment. We observed elevated levels of CB2 protein (46 kDa) in OA-affected cartilage that was reduced by chronic administration of JWH-133 (Fig. 3A). This demonstrates that the receptor for this compound was present in the i.a. space, while the response of protein expression to the treatment suggested an effective dose of JWH-133. The level of CB1 protein (52 kDa) was augmented in OA-affected cartilage, but JWH-133 treatment did not reverse that effect (Supplementary Fig. S5A). The expression of selected MMPs, TIMPs and COMP is particularly important for OA pathogenesis in the aspect of chondrocytes' degeneration; therefore, their levels were evaluated in the cartilage of MIA-injected rat knees.
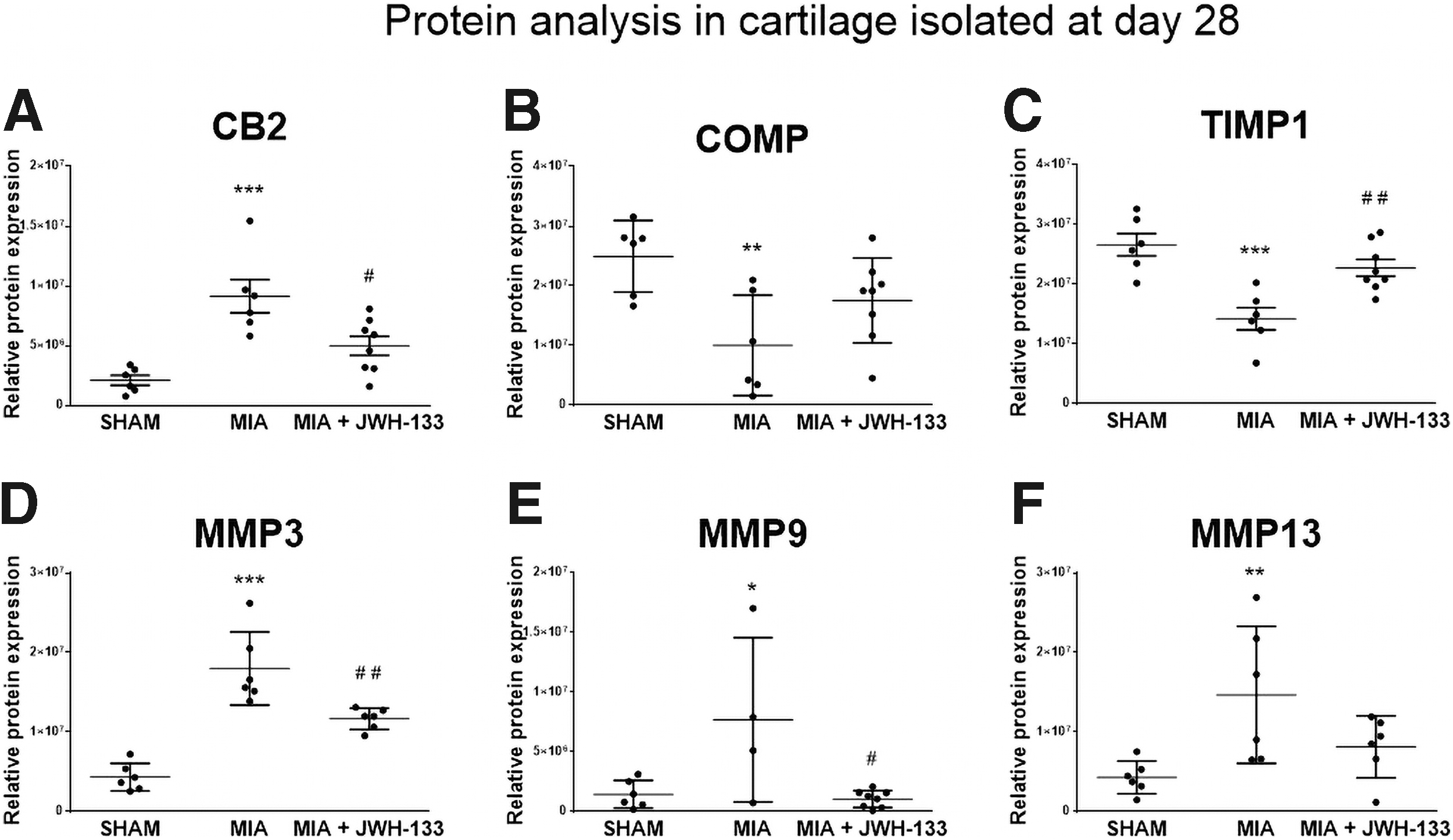
Protein expression of CB2 receptor
We found decreased protein level of TIMP1 (26 kDa) and COMP (100 kDa) in OA-affected cartilage that were restored by JWH-133 treatment (Fig. 3B, C). Furthermore, we performed proteomic analysis of selected MMPs and TIMPs. Our data revealed that the expression of MMP3, MMP9, and MMP13 (54 kDa proform, 98 kDa proform, and 48 kDa proform) was elevated in the cartilage of MIA-treated animals (Fig. 3D–F). Moreover, there was significant alleviation of expression for MMP3 and MMP9 observed after treatment with JWH-133.
Expression of MMP13 was alleviated to the initial level after treatment with JWH-133
To further investigate the action of JWH-133 on chondrocytes in the development of OA, we used primary HC cultures. Cells were subjected to MIA (3 μM), JWH-133 (10 μM), and AM-630 (potent CB2 agonist used to block molecular effects of JWH-133; 10 μM). Doses were optimized based on a separate set of experiments (Supplementary Fig. S1). We measured level of CB2 transcript in HC to elucidate effect of MIA and JWH-133 on those cells (in vivo samples can be obtained from animals, which can include some of the synovial fibroblasts or macrophages that infiltrated the joint tissue during development of OA). The molecular analysis revealed upregulation of Cnr2 transcript (encoding CB2 receptor) after MIA treatment that tended to decrease after additional incubation with JWH-133 (Fig. 4A).
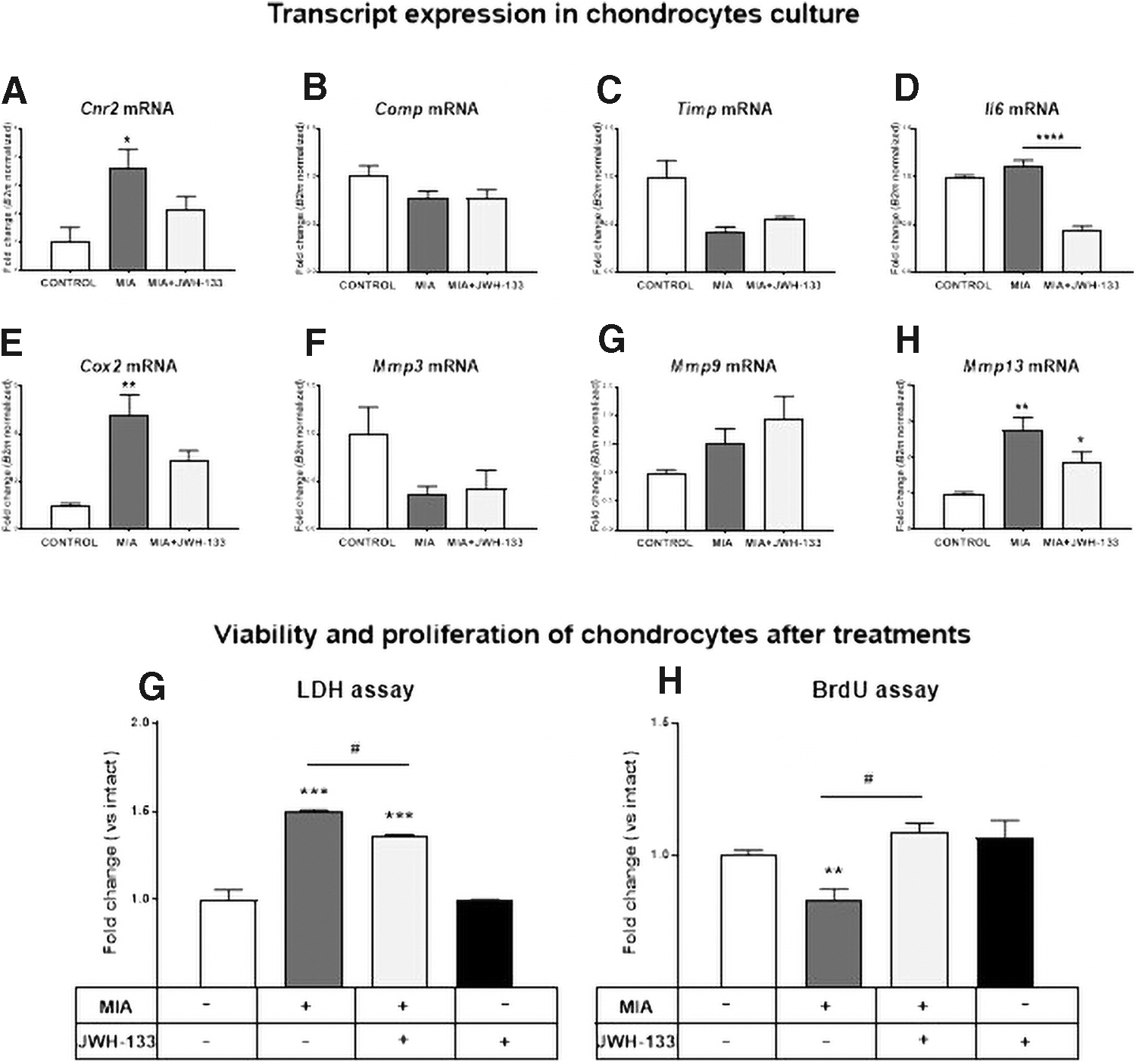
Molecular changes in HC after treatment with MIA (3 μM) and JWH-133 (10 μM). qRT-PCR analysis of Cnr2 receptor
The changes in the expression of Comp and Timp1 transcripts were not as robust as in the case of animal tissue (Fig. 4B, C), although it may be due to differences in treatment paradigm. Nonetheless it has to be taken under consideration that changes in the protein levels present in the animal model may be due to the presence of other cell types in collected tissue. We observed a decrease of Il6 and Cox2 (inflammatory markers) transcript levels after treatment with JWH-133 (Fig. 4D, E).
In addition, we examined expression levels of selected MMPs' transcripts, namely Mmp3, Mmp9, and Mmp13 (Fig. 4F–H). The expression of Mmp3 tended to decrease in MIA-treated HC, regardless of JWH-133 treatment (Fig. 4F). There were no significant differences in Mmp9 transcript levels in any of the performed experiments (Fig. 4G). The most interesting expression pattern was found for Mmp13, which resembled the corresponding protein expression in OA-affected animal cartilage (Fig. 4H).
JWH-133 reverses proliferation arrest caused by MIA treatment in HCs
After unveiling the mechanism of JWH-133 action in HC, it was necessary to determine whether MIA affects cell survival, proliferation, or motility. To examine the effect of tested compounds on chondrocyte viability, we performed an LDH assay, observing a decreased survival rate after MIA treatment that was only partially reversed by JWH-133 treatment (Fig. 4G). We also performed a BrdU assay to determine the effect of JWH-133 on the proliferation of MIA-treated HC. We observed decreased cell proliferation caused by MIA that could be completely reversed by JWH-133 treatment (Fig. 4H).
JWH-133 reversal reduced motility of MIA-treated HC in a manner dependent on CB2 activation
To evaluate whether the effect of JWH-133 is CB2-dependent, a scratch-wound test was performed using an antagonist of this receptor, AM-630, at a concentration of 10 μM. The compound was coadministered with JWH-133 (10 μM) and MIA (3 μM). The CB2 receptor antagonist was administered 15 min prior JWH-133 to saturate the CB2 receptors in the chondrocytes. The scratch-wound assay was conducted for 24 h. Migration of HCs significantly decreased after treatment with 3 μM MIA (Fig. 5A–C). This inhibition was abolished by administering JWH-133 at a concentration of 10 μM (Fig. 5A, D). Blocking the CB2 receptor impaired the protective effect of JWH-133, leading to the conclusion that the effects induced by JWH-133 are dependent on CB2 receptor activation (Fig. 5A, E).
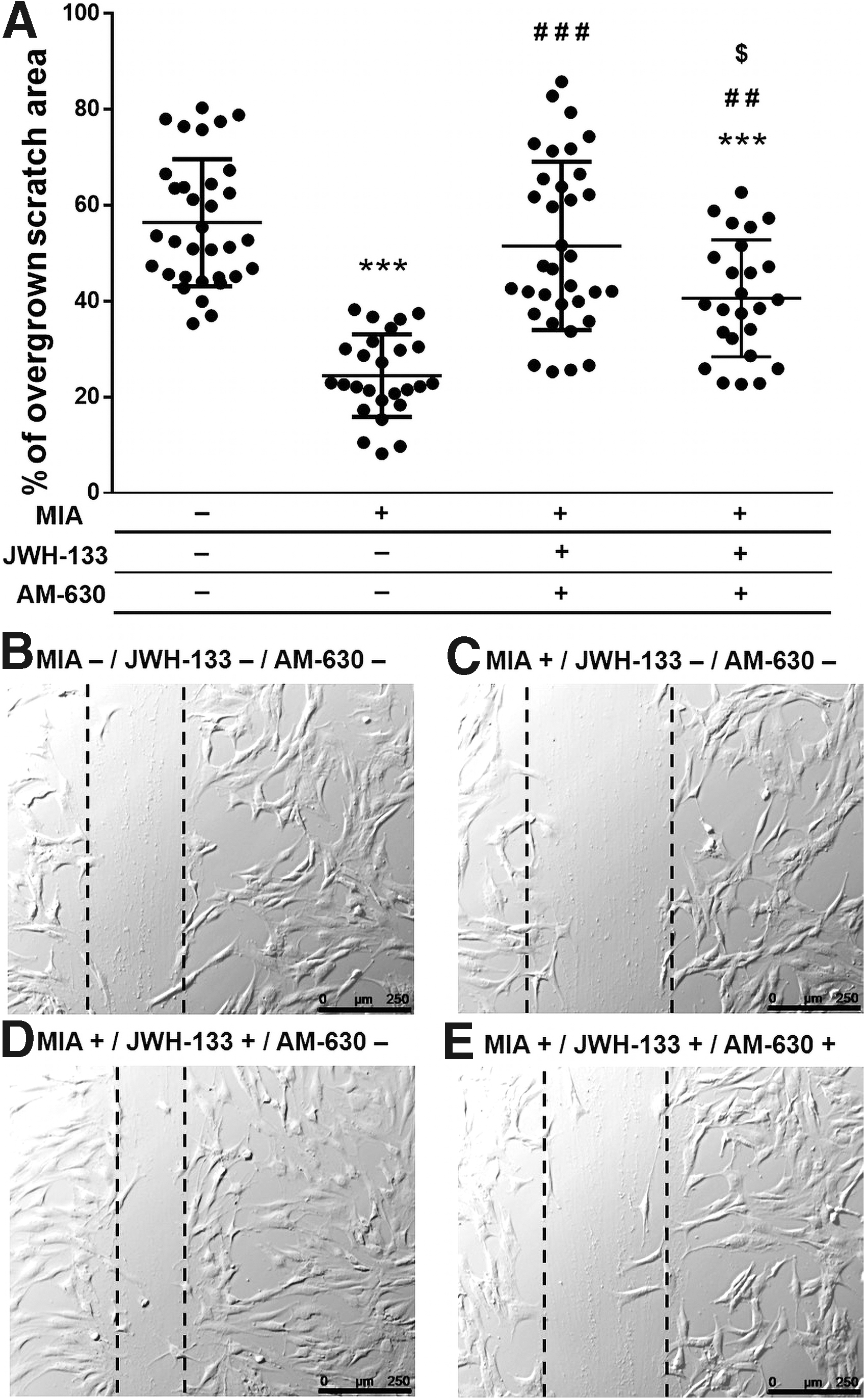
Effects of JWH-133 (10 μM) and AM-630 (10 μM) on HC motility in wound healing assay. The cells were seeded on noncoated surface and grown to confluence. Cells were treated with tested compounds or their mixtures for 24 h prior the experiment. After making the wound in a monolayer, the cells were recorded for 24 h. Data were presented as graphs representing scratch closure expressed in % of closed scratch
Discussion
While there are continual increases in understanding of the role of the ECS in OA, its role in cartilage tissue degeneration has been left behind. Here, for the first time, we investigate the effect of CB2 stimulation on chondrocytes migration and proliferation, and the expression of corresponding MMPs, both in vitro and in vivo, in an MIA-induced rat model of OA. We present results showing the protective action of JWH-133 on chondrocyte proliferation and mobility, which was observed as the alleviation of pain behavior in in vivo studies.
The studies were conducted in the MIA rat model, focused on the chronic pain phase. The two-phase development of pain symptoms in this model has already been described in our previous studies.12,19 Here, we also used the JWH-133 treatment paradigm starting from day 14 after model induction, due to the correlation of such treatment with the clinical situation, where patients reporting pain symptoms are characterized by changes in the articular cartilage. We observed alleviation of joint hypersensitivity after three doses of JWH-133. Furthermore, the effect had not diminished at the end of the experiment. These data are promising as they highlight a potential therapy for OA patients. The only concern might be the administration route, as frequent i.a. injection is not recommended for the long-term treatment of musculoskeletal diseases. 22
The anti-inflammatory and antinociceptive effect of CB2 agonism is well described in the literature of various pathologies and cell types. 23 Our research assumed that the analgesic effect of JWH-133 is based not only on the anti-inflammatory effect, but also on the protective effect on the articular cartilage. This was confirmed as we observed that knee joints isolated from animals subjected to JWH-133 treatment restored levels of COMP and TIMP1. The reduction in COMP protein expression observed in MIA-injected animals indicated the irreversible breakdown of cartilage structure, which may contribute to the development of chronic OA-related pain. Interestingly, tissue isolated from OA patients also displays reduced COMP expression. 24 Expression of both COMP and TIMP1 was shown to be altered in the articular cartilage of OA patients25,26 and in the MIA-induced model of OA, 12 highlighting the validity of the selected research model.
Our previous studies in an MIA-induced model of OA showed that the development of the second phase of pain symptoms is related to degradation of joint cartilage.12,19 Among the many MMPs related to cartilage damage, we chose to look at those that have been shown to be promising therapy targets or biomarkers for OA in the past.16,27 Functional linkage between MMPs and ECS in cartilage during the development of OA has been suggested by our group previously. 12 Based on those studies, we examined expression levels of MMP3, MMP9, and MMP13. All these proteins showed increased expression during the development of OA that was abolished after administration of JWH-133. Nevertheless, in animal studies, we analyzed all the cells that built up articular cartilage.
In addition, we must consider the possibility of contamination of the collected samples with cells from the subchondral layer or the synovium. This approach is beneficial when looking at all aspects of OA at once, but it makes it difficult to discover the molecular mechanisms underlying the pathogenesis or therapy of OA. Therefore, in our studies, we used primary cultures of HC to evaluate results obtained from the animal model. Our results centered on MMP13, a major enzyme that targets cartilage for degradation, showing that MIA treatment induced similar expression changes to OA, which could be reversed by JWH-133. Clinical studies revealed that patients with articular cartilage destruction have high MMP13 expression, 28 suggesting that increased MMP13 may be associated with cartilage degradation.
Studies have also shown that transgenic Mmp13-overexpressing mice develop spontaneous OA-like articular cartilage destruction. 29 Therefore, attenuation of MMP13 expression through agonism of CB2 receptors may create novel therapeutic opportunities.
Given the importance of MMP13 in migration and proliferation processes, we determined the chondrocytes' behavior. 30 Decreased chondrocyte proliferation and migration are one of the hallmarks of cartilage degradation in OA. Therefore, discovering molecular targets involved in those processes might be a first step to the development of disease modifying therapies. Our data showed that MIA treatment of chondrocytes significantly decrease their proliferative abilities and decreased motility. In addition, immunofluorescence staining showed a decrease in the number of actin fibers in chondrocytes under the influence of MIA, which demonstrates the toxicity of this compound to the cytoskeleton and its inhibition of their proliferative capacity (Supplementary Fig. S6). These observations, together with increased protein levels of MMP13 in MIA-treated chondrocytes and cartilage, suggest that an MMP13 inhibitor may have potential for the treatment of OA progression.
However, no MMP13 inhibitor is available for clinical use, predominantly because of inadequate efficacy. 31 Therefore, the idea of using CB2 receptors as mediators of MMP13 activity appears to be appealing. In our studies, both proliferation and chondrocytes motility were at least partially restored by JWH-133 administration. Moreover, we were able to show that the pharmacological effect on cell migration is CB2 dependent. Although the certainty around the mechanism could be provided by the use of the CB2 antagonist in a broader range of the experiments, especially in the case of experiments on compounds toxicity, the conclusions are made without additional evaluation of CB2 blockage. Nonetheless, further experiments allowed conclusions to be drawn regarding the CB2-dependent effect of JWH-133 in the context of cell migration.
In addition, we have to take under consideration that measured cell migration (in wound healing assay) is the result of cell mobility and its ability to proliferate. While some of our studies showed that JWH-133 increases cell proliferation (Supplementary Fig. S2B), further research is needed to distinguish between these processes. We believe that provided data will be a starting point for further more detailed studies on the involvement of ECS in cell proliferation and motility.
We concluded that provided experimental evidence is sufficient to support the potential of CB2-based drugs in novel applications aimed at preserving articular cartilage and joint function instead of simply alleviating OA symptoms as existing treatments do. Together with the anti-inflammatory and antinociceptive properties of CB2 agonists, we propose novel, innovative dual-acting therapy on OA, for which further investigations are required. Especially in the aspect of long-term CB2 agonism and its influence on cartilage as well as limitations of target-specific effects that were not studied in the presented article.
Footnotes
Authors' Contributions
N.M. designed the study, obtained funding, induced the MIA model, performed behavioral experiments, isolated knee joint samples, analyzed qPCR data, conducted the statistical analyses, interpreted the data, prepared figures, and wrote the article. J.B. performed scratch healing assay and analyzed the data. M.K. performed the qPCR experiments and analyzed the qPCR data. A.P. performed the western blot assays, analyzed the western blot results. J.D. was consulted for interpretation of the scratch healing assay data. K.S. obtained funding, participated in study design, and reviewed the article. All authors have approved the final version of the article.
Author Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
Supported by Ministry of Science and Higher Education grant: 0044/DIA/2013/42 awarded to Natalia Malek, National Science Centre grants: OPUS UMO-2014/13/B/NZ7/02311, SONATA BIS/NCN/2012/07/E/NZ7/01269 awarded to Katarzyna Starowicz and Maj Institute of Pharmacology PAS statutory funds. NM is a recipient of the scholarship for young researchers (no. 444/STYP/13/2018) funded by the Ministry of Science and Higher Education.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.