
Abstract
Background:
Alterations in the endocannabinoid system (ES) have been described in Alzheimer's disease (AD) pathophysiology. In the past years, multiple ES biomarkers have been developed, promising to advance our understanding of ES changes in AD.
Discussion:
ES biomarkers, including positron emission tomography with cannabinoid receptors tracers and biofluid-based endocannabinoids, are associated with AD disease progression and pathological features.
Conclusion:
Although not specific enough for AD diagnosis, ES biomarkers hold promise for prognosis, drug-target engagement, and a better understanding of the disease. Here, we summarize currently available ES biomarker findings and discuss their potential applications in the AD research field.
Introduction
Alzheimer's disease (AD) is the most common cause of dementia worldwide, affecting 1 in 10 people over age 65. 1 Genetic and neuropathological evidence from clinical and experimental studies suggests a crucial role played by the deposition of amyloid-beta (Aβ), a protein derived from the amyloid precursor protein (APP), and the formation of neurofibrillary tangles (NFTs) caused by the phosphorylation of the microtubule-associated protein tau in the development of AD. 2 It has been proposed that Aβ deposition triggers a series of pathological events that lead to tau hyperphosphorylation, synaptic dysfunction, and neurodegeneration, clinically manifested by cognitive decline and, ultimately, dementia.3,4
The Aβ deposition tends to accumulate through the entire cortex while the NFTs are over the neocortex, following the Braak staging.5,6 The Braak stage is based on the hierarchical anatomical localization of NFTs,5–7 in which stages I–II correspond to tau aggregation in the transentorhinal region, stages III-IV in the limbic regions, and stages V–VI in the neocortical region. 6 At present, the clinical course of AD is understood as a cognitive continuum, encompassing (i) an asymptomatic phase where pathology begins to accumulate in the brain in the absence of symptoms (i.e., preclinical AD), (ii) a mild cognitive impairment (MCI) phase - cognitive impairment of insufficient severity to meet criteria for dementia, and (iii) a dementia phase.
In addition to Aβ and tau pathology, a sustained activation of the brain's inflammatory pathways is gaining attention as a potential core component of AD. 8 Specifically, when the acute and time-limited inflammatory response develops into a chronic inflammatory state, it is thought to exacerbate AD pathology. 8 Regarding particular cellular relationships with these processes, microglial cells seem to be persistently activated in this chronic inflammatory state, leading to a sustained release of cytokines and cytotoxic mediators. This environment is accompanied by the response of astrocytes that become reactive and contribute to homeostasis disturbances in the central nervous system (CNS).9,10
Regarding current clinical research advances in AD, the National Institute on Aging and Alzheimer's Association (NIA-AA) proposed a research framework in 2018, adopting the AT(N) classification scheme to biologically define AD, with biomarkers for Aβ (A), tau (T), and neurodegeneration (N). 4 Based on patient biomarker profiles, the AT(N) system determines not only whether an individual is within the AD biological continuum but also, more specifically, where each profile falls regarding the continuum's spectrum of severity. The NIA-AA research framework encourages the inclusion of additional biomarker categories, which culminated in the early development of ATX(N) system.
The X represents candidate biomarkers for additional pathophysiological mechanisms.11,12 It is important to emphasize that X biomarkers may serve to prognosis, drug-target engagement and to develop our understanding of additional pathophysiological mechanisms. In this context, biomarkers able to capture ES changes in AD could be envisioned as potential candidates for the X component of the expanded biomarker framework.
It has been demonstrated that the endocannabinoid system (ES) plays an important role across all of these pathological stages of AD, including the promotion of intrinsic repair mechanisms and the modulation of pathological processes, including neuroinflammation. 13 However, the field currently lacks a synthesis on where it stands regarding advances in the development of ES biomarkers for AD. Herein, we comprehensively review and critically discuss the changes in the ES observed in AD and potential novel biomarker candidates.
The Endocannabinoid System
Cannabinoid receptors
Cannabinoid 1 (CB1) receptors are encoded by CNR1 gene and represent the most abundant G protein-coupled receptors in the CNS. 14 Curiously, these receptors have a similar spatial distribution pattern between the brains of rats and humans. Specifically, CB receptors are abundantly expressed in the neocortex, hippocampus, amygdala, basal ganglia, cerebellum, and emetic centers of the brainstem.15,16 Regarding its cellular location, CB1 receptors are mainly observed in neuronal axons and presynaptic terminals, where they modulate neurotransmitter release. 17 The activation of CB1 receptors by synthetic CBs has been shown to modulate a wide range of neurotransmission systems, including GABergic, glutamatergic, 18 cholinergic, 19 and noradrenergic systems. 20
These receptors have also been described in postsynaptic neurons and glial cells. Despite its low abundance in these cells, glial expression of CB1 in glial cells has been reported to influence multiple physiological roles, such as long-term depression.21–23 Beyond the CNS, CB1 receptors have also been observed in lower concentrations in other peripheral tissues, such as those in the cardiovascular, reproductive, and gastrointestinal systems.24–26
CB2 receptors are encoded by the gene CNR2 and exhibit 48% homology with CB1 receptors. 27 They are mainly expressed peripherally in tissues and cells involved in immunological responses. 28 In the CNS, they are mainly present in microglial cells, 29 with increasing evidence that they might be expressed in some neuronal populations. 30 Moreover, CB2 expression in the CNS seems to be inducible: from a low basal level, it can display a drastic 100-fold increase following injury or during inflammation. 31 This increase is attributed to the augmented expression of CB2 receptors in microglial cells, or to the infiltration of peripheral immune cells expressing this receptor.
Regarding potential physiological roles of CB2 receptors in the CNS, considerably less is understood. However, they are known to interact with the neuro-immune system and with synaptic function. CB2 receptors regulate immune cells in several ways, including: (i) inducing immune cell apoptosis, (ii) suppressing proliferation of immune cells and the production of proinflammatory cytokines/chemokines, (iii) increasing secretion of anti-inflammatory cytokines, (iv) inducing regulatory T cells, and (v) promoting the trafficking and migration of immune cells.32,33
CB neurotransmitters
In 1992, the search for endogenous ligands of these receptors resulted in the discovery of the first endocannabinoid (eCB): anandamide (arachidonoyl ethanolamide [AEA], from the Sanskrit word ananda for “bliss”). 34 A second endogenous ligand was described right afterward, the 2-arachidonoylglycerol (2-AG).35,36 Although 2-AG and AEA are the most abundant eCBs in the brain,37,38 other molecules chemically similar to eCBS have also been identified such as the 2-arachidonyl-glycerol ether (noladin ether), oleoylethanolamide, n-arachidonoyl-dopamine, and virodhamine. The eCBs are lipid messengers, which allows them to diffuse across membranes.37,38
As the natural agonists of CB receptors, eCBs affect neurotransmitter release and synaptic plasticity.39–42 The eCBs are considered retrograde messengers due to the localization of the CB1 receptors in the presynaptic neurons and their capability to inhibit synaptic transmission. Moreover, postsynaptic neurons have the ability to increase eCBs production when activated, partly via the increase of intracellular Ca2+ levels.43–46
AEA synthesis, transportation, and metabolization
The N-arachidonoyl phosphatidyl ethanol constitutes the main substrate for AEA synthesis (Fig. 1). Its synthesis has five different brain metabolic pathways, involving different phospholipases and possibly arachidonic acid and ethanolamine.14,42,43 The AEA's half-life is very short because when released in the synaptic cleft it is quickly taken up by a high-affinity AEA membrane transporter expressed in neurons and glia.47,48 The second step of AEA metabolism includes the hydrolyzation though fatty acid amide hydrolase (FAAH). 49 The AEA is predominantly metabolized into arachidonic acid (AA) and ethanolamine.44,50 This process is selective, saturable, temperature-dependent and does not require ATP or sodium ions. 44
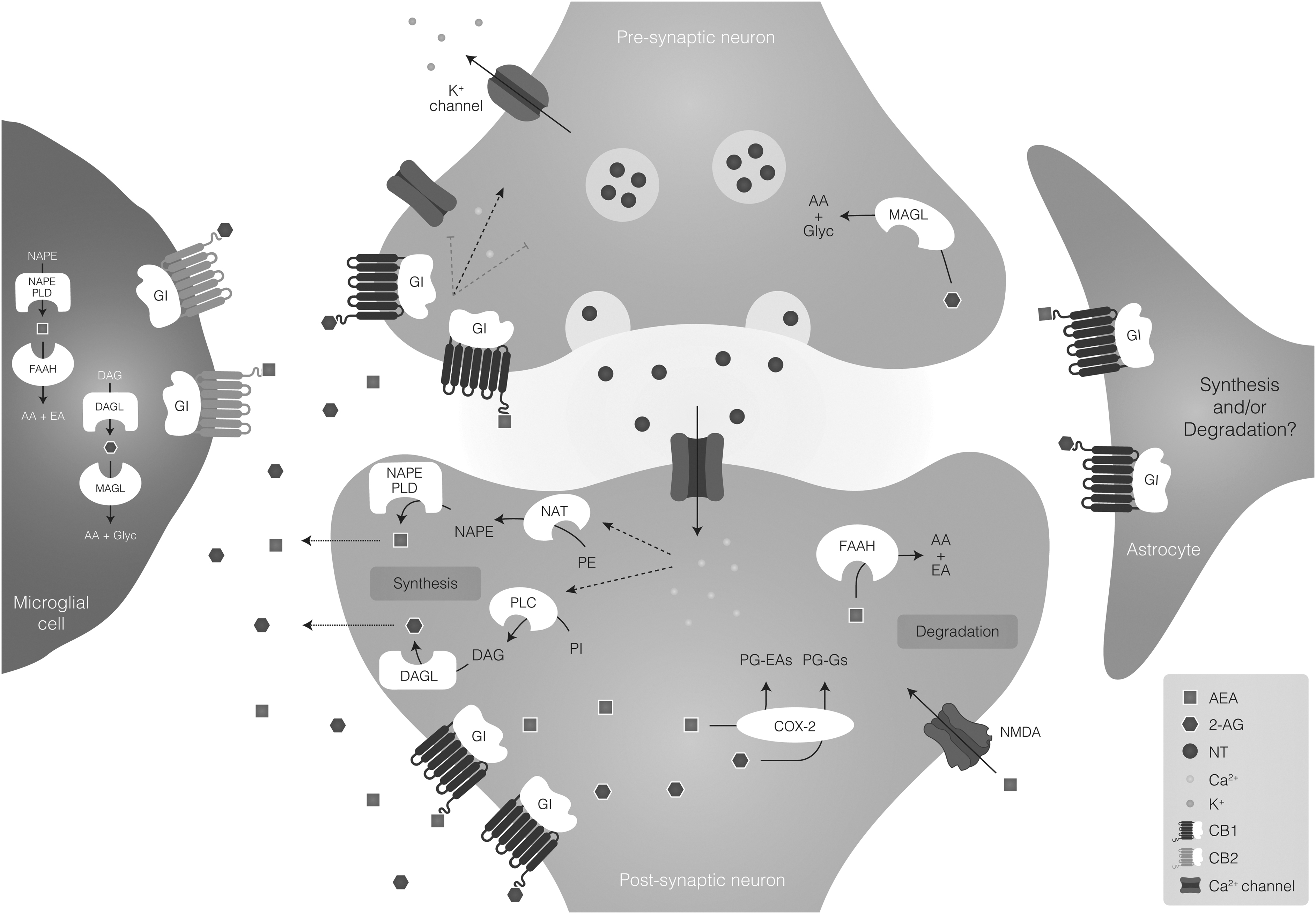
Endocannabinoid signaling on the central nervous system. In the ES, the two main receptors are CB1 and CB2. CB1 receptor is enriched in neurons, and at a lower density in the astrocytes, whereas CB2 receptor is enriched in microglia. The endocannabinoids anandamide (AEA) and 2-AG are the most widely recognized ligands of the ES, and the signaling is characteristically related to the modulation of synaptic activity. NT release from presynaptic neurons activates N-methyl-D-aspartate receptors in the postsynaptic neurons. In response to the increase of cytoplasmic calcium (Ca2+) levels, the enzyme DAGL catalyzes the synthesis of 2-AG from DAG. The NAPE-PLD catalyzes the synthesis of AEA from the precursor NAPE. Subsequently, 2-AG and AEA are released into the synaptic cleft, traveling in a retrograde way to activate the CB1 receptor located in the presynaptic terminals to inhibit further NT release through the suppression of Ca2+ influx. 2-AG is also able to activate CB1 receptors located in astrocytes. The AEA also binds and activates transient receptor potential cation channel subfamily V member 1. 2-AG is mainly degraded by the enzyme MAGL, whereas AEA is degraded by FAAH. 2-AG, 2-arachidonoylglycerol; AEA, arachidonoyl ethanolamide; CB, cannabinoid; DAG, diacylglycerol; DAGL, diacylglycerol lipase; ES, endocannabinoid system; FAAH, fatty-acid amide hydrolase; MAGL, monoacylglycerol lipase; NAPE-PLD, N-acyl-phosphatidylethanolamine phospholipase D; NT, neurotransmitter.
2-AG synthesis, transportation and metabolization
The 2-AG's main substrate for synthesis is arachidonoyl-containing phosphatidyl inositol bis-phosphate (PIP2). 14 Its main route of synthesis occurs through sequential hydrolysis of an arachidonoyl-containing-PIP2 by PLCβ followed by the hydrolysis of the resulting diacylglycerol by diacylglycerol lipase (DAGL) (Fig. 1).14,43 2-AG is then released in the synaptic cleft, and it is thought that 2-AG is taken up by brain cells through passive diffusion.47,51
The metabolism of 2-AG follows the same pathway as AEA: (i) rapid uptake into the brain cells followed by (ii) hydrolysis mediated by monoacylglycerol lipase (MAGL). 52 The major products of this hydrolysis are AA and glycerol. 53 In addition, 2-AG can also be metabolized by FAAH, into arachidonic acid (AA), ethanolamine, and glycerol, and by cyclooxygenase 2, into prostaglandin ethanolamine's and prostaglandin glycerol esters. 40 Metabolization of 2-AG occurs on presynaptic axon terminals, predominantly in terminals of GABAergic interneurons. 54
The ES in Alzheimer's Disease
CB receptors: CB1 and CB2
Since the early 1990s, the association between alterations in the ES and AD pathology has been investigated 55 (Fig. 1). So far, it is known that the expression of CB receptors in AD varies across brain regions and disease stage (Supplementary Table S1). A postmortem autoradiography study was the first to investigate the ES changes in AD brain, finding that CB1 receptors were downregulated in the multiple brain regions, including the hippocampus, neocortex, basal ganglia, and brainstem. 55
However, a comparison with postmortem brain tissue from cognitively unimpaired (CU) individuals indicated that this decrease was likely related to the aging process itself rather than specifically to AD. 55 Almost two decades later, Farkas et al. demonstrated that CB1 expression, indexed by [125I]SD-7015 autoradiography, varied inversely with Braak staging for tau pathology in AD brain. 56
AD-like transgenic models also suggested a biphasic course of CB1 receptor expression. Using the APP/PS1 mice, double transgenic mice model overexpressing a human APP with the Swedish (APPswe) mutation and a human presenilin 1 (PS1) with the dE9 mutation, a decrease in CB1 receptor density was paralleled by an elevated expression of proinflammatory cytokines, astrocyte reactivity, and an exacerbation in neuropathological features of AD. 57
A related study using the 3xTg-AD mice model, which harbors three human mutant an genes APPswe, PS1M146V, and TauP301L, demonstrated that, 58 although the whole brain CB1 receptor density was unchanged at ages 2 and 6 months, it was downregulated in the basolateral amygdala complex and dorsal hippocampus at 12 months. Aβ deposition was also found to correlate inversely with CB1 receptor expression. 58
Additional work using the 5xFAD mouse model, a transgenic model that overexpresses human APP (Swedish, Florida and London) and PS1 (M146L and L286V) transgenes, Florida (I716V), and London (V717I) mutations in APP, and the M146L and L286V mutations in PS1, showed that pharmacological blockade of the CB1 receptor by the inverse agonist SR141716 led to an exacerbated inflammatory response, but with no further impairment in behavior or spatial memory. 59 Indeed, CB1 receptors have important physiological functions, and the fine regulation of their levels/activation seems to be crucial for maintaining proper brain function.18,19,20,24–26
Postmortem brain tissue from AD patients has been shown to express CB2 receptors at high levels in the microglial cells surrounding Aβ plaques, being potential markers of microglial activation. 60 Indeed, CB2 receptor expression levels have been shown to correlate positively with Aβ1-42 levels and senile plaque scores in the frontal cortex of AD patients. 61 The lack of CB2 receptor expression has also been associated with a decrease in Aβ plaque deposition, being more prominent at 14 months, in the neocortex and hippocampus.62,63
Further, even though CB2 receptor depletion was associated with improved spatial learning and memory in 6-month-old transgenic and control mice, this effect was no longer seen at months 9 and 14. 62 Additional work using this transgenic model showed that the absence of CB2 receptors decreased pro-inflammatory response in mice, possibly due to impaired microglial activation. 62 Another study corroborated these findings using the same animal model, demonstrating that CB2 receptor deficiency was associated with reduced neuroinflammation as well as decreased Aβ plaque levels and neuronal loss. 63
Endocannabinoids
The effects of eCBs on the brain and their influence in AD have been investigated in several studies (described in Supplementary Table S1). Previous research conducted in primary cortical neuron cultures demonstrated that treatment with the eCBs anandamide (AEA) and 2-AG protected cells from Aβ-induced apoptosis. 64 In addition, analyses in the hippocampus of postmortem brain tissue suggested that the molecular composition of 2-AG signaling networks suffers modifications around senile plaques. 65
Specifically, it was demonstrated that there was a prolonged depolarization-induced suppression of inhibition in the synapse, which could contribute to synaptic silencing in AD. 65 However, investigation of eCBs alterations in the Tg2576 transgenic mice demonstrated no changes in 2-AG levels in the hippocampus. 66 On the other hand, AEA levels were found to be reduced, and FAAH metabolization into ethanolamine was significantly lower in the Tg2576 mice. 66
Additional research examining the effect of inhibiting the metabolization of 2-AG, via the inactivation of MAGL, revealed a significant reduction in Aβ1-42 levels in the hippocampus along with a suppression of neuroinflammatory response (Supplementary Table S1). 67 The inhibition of MAGL also resulted in preserved hippocampal synaptic structure and function and improved long-term synaptic plasticity, spatial learning, and memory. 67
CB Biomarkers in AD
In 1994, the first ES radioligand binding site was identified using in vitro autoradiography with the CB1 receptor agonist [ 3 H]CP 55.940 (Table 1). 55 The CB1 receptor binding site availability was found to be decreased in the hippocampal formation, neocortex, basal ganglia, and brainstem in postmortem AD brain tissue; however, the authors speculated that these changes could be associated with aging itself and with other cortical pathologies. In a subsequent study using the same tracer, the density of CB1 receptors in pyramidal cells in the CA1 region of the hippocampus was upregulated at earlier stages of tau pathology (Braak I/IV) and decreased at later stages (Braak V/VI), 68 whereas it was upregulated in layer IV of the frontal cortex (Braak III/IV). 68
Most Relevant In Vitro and In Vivo Studies Conducted with Endocannabinoid Tracers
In Kd=equilibrium dissociation constant; IC50=inhibitory concentration and Ki=inhibitory constant.
[11C]OMAR=[11C]-JHU75528.
Only studied regions were mentioned.
AD, Alzheimer's disease individuals; CB, cannabinoid; FAAH, fatty acid amide hydrolase; HC, healthy control individuals; MAGL, monoacylglycerol lipase; MMSE, mini mental state examination; PTSD, post-traumatic stress disorder individuals; SC, schizophrenia; TC, trauma-exposed controls individuals; Tg, transgenic model; WT, wild type.
Similarly, another CB1 receptor agonist, the [125I]SD7015, presented an increased binding in the prefrontal cortex of postmortem tissue from patients with early tau pathology (Braak I/II), but not in patients in more advanced stages (Braak V/VI) 56 (Fig. 2). These findings are in line with the idea that the ES might be involved in an early response to AD pathology. Unfortunately, no autoradiography studies have been conducted with tracers targeting the CB2 receptor.
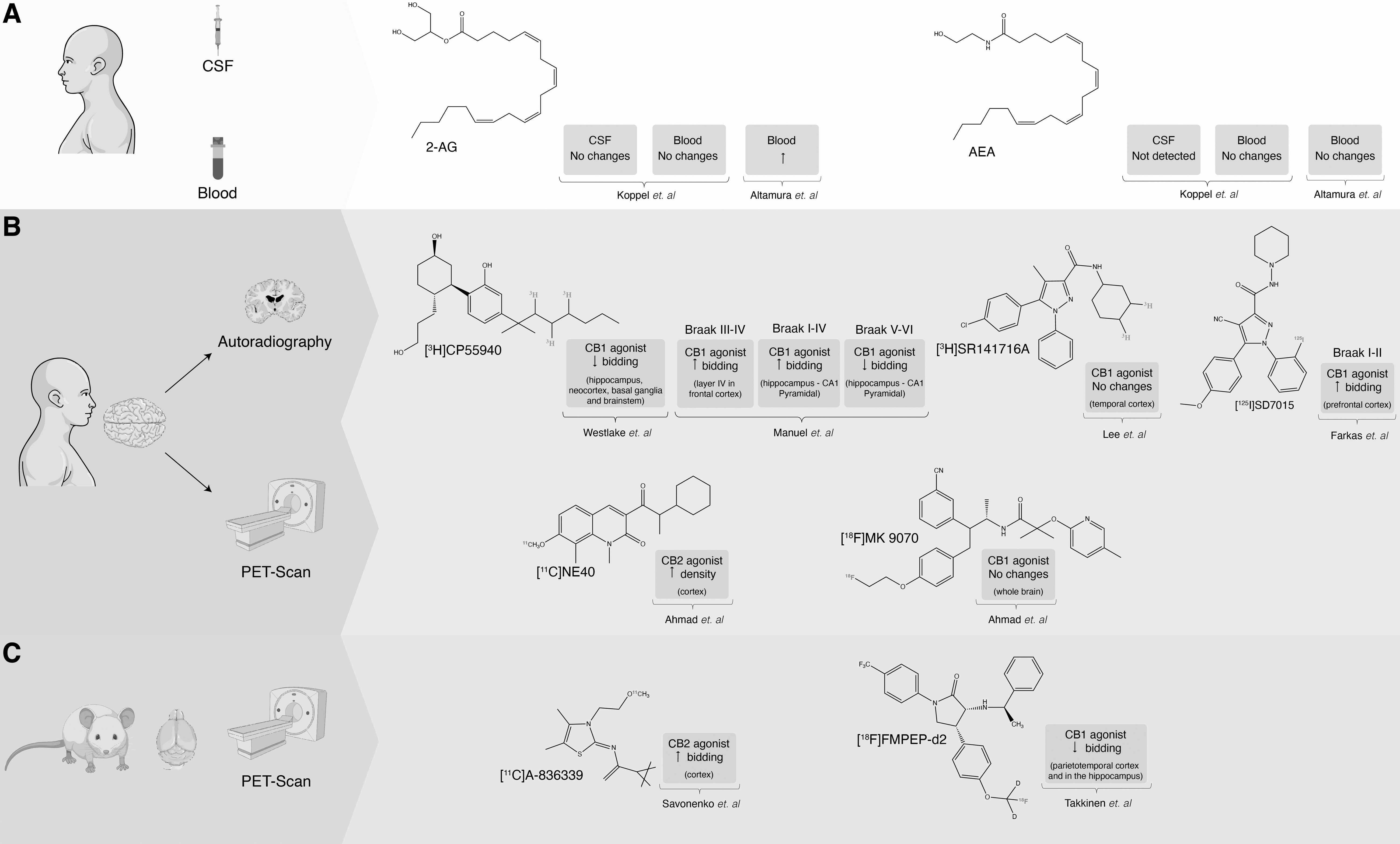
Overview of endocannabinoid biomarkers.
eCB positron emission tomography studies in AD
The positron emission tomography (PET) imaging technique allows for the non-invasive quantification of numerous biological and pharmacological targets trough the pharmacokinetic analysis of radiopharmaceuticals, which are molecules labeled with radioisotopes. In AD, PET has been used to estimate aggregated proteins (such as amyloid and tau), receptors (such as the 18 kDa translocator protein), metabolism (such as glucose brain metabolism), and others. 69
Despite promising results for ES-AD autoradiographic studies, in vivo human PET studies have only recently been performed. However, other CB1 receptor tracers have been tested in AD patients. First, it was demonstrated that the binding of [18F]MK-9470, a CB1 receptor inverse agonist, did not change in AD patients as compared with CU individuals. 70 In addition, no significant correlation was found between retention of the Aβ-PET [11C]PIB tracer and the CB1 receptor availability. Moreover, [18F]MK-9470 binding was not associated with cognitive function (measured by mini mental state examination and auditory verbal learning test) nor with CB1 receptor availability and apolipoprotein (APOE) ɛ4 carriership. 70
Although no further human studies have been conducted to date, lower [18F]FMPEP-d2 binding has been found in the parietotemporal cortex and hippocampus in the APP/PS1-21 mouse model. 71 Finally, although PET imaging of the CB1 receptor seems an interesting biological target in AD, so far, there are no promising results using CB1 tracers.
Regarding CB2 tracers, Horti et al. showed an increase in the binding of [11C]A-836339, a CB2 agonist, in the cortex of an AD-like APP/PS1 mouse model, and also in response to neuroinflammation induced by lipopolysaccharide. A significant increase in CB2 receptor availability (around 30%) in regions with high Aβ plaque load was also observed, suggesting that CB2 receptor upregulation may be directly associated with Aβ pathology. 72 Using another CB2 agonist, [11C]NE40, lower CB2 receptor availability was seen in cortical and subcortical regions in AD patients. 73
These divergent findings between mice and human data might be attributed to heterogeneous presentations of AD in humans that may not be found in the animal model.63,74 In addition, murine microglia seem to present a different response to Aβ species if compared with human microglia. 75
Overall, these findings appear to be consistent with a biphasic course of the ES in AD, similar to the trajectory described for microglial activation in AD, where an initial anti-inflammatory (protective) peak during the preclinical phase is followed by a second pro-inflammatory (detrimental) microglial phenotype during the symptomatic (MCI/dementia) phase. 76 Nevertheless, further studies with larger sample sizes and novel ES biomarkers are needed to evaluate the validity of this hypothesis.
eCB PET studies in other brain conditions
Besides the ES tracers already tested in AD, in vivo studies have shown promising results for other ES ligands (Table 1). Though most of these studies have focused on CB1 and CB2 receptors,72,77–83 some have targeted the main enzymes involved in eCB metabolism to address changes in the ES.84–86 These PET tracers have not been tested yet in AD but demonstrate favorable properties to capture changes in the ES in other brain conditions.
CB receptor tracers (CB1 and CB2)
In autoradiography studies, CB1 tracers have been the most investigated up to date, particularly [ 3 H]MePPEP and [11C]CB-119 due their high affinity to the binding site.77,79 Using both tracers, CB1 receptor binding was elevated in cortical and hippocampal regions and even enhanced in cerebellar regions.
An additional tracer of interest is [11C]OMAR, which shows a high volume of distribution (Vt) in the cortex, hippocampus, and lentiform nucleolus.78,80,82,87 Studies using [11C]OMAR have been performed in animal models (mice and baboons) and humans. The specificity of [11C]OMAR for CB1 receptor was evidenced in baboons by a blocking study with rimonabant, a selective CB1 inverse agonist. 87 In addition, another study performed in mice demonstrated that [11C]OMAR has a rapid uptake into the brain followed by a relatively slow clearance. 88
Regarding other neuropsychiatric conditions, Wong et al. investigated [11C]OMAR uptake in schizophrenia. Interestingly, this study highlighted an important gender difference in receptor availability. 78 Specifically, Vt in women was 23% higher than in men, whereas cerebral uptake was 17% higher in men. 82 Further studies demonstrated a lower CB1 receptor availability in schizophrenia patients, compared with healthy individuals. Interestingly, the use of antipsychotic drugs and tobacco seems to increase CB1 receptor availability, being an important co-factor in the analysis. 89
Among the tracers discussed in this section, [11C]OMAR seems to have the greatest potential to be tested in AD individuals as it shows favorable kinetics and increased Vt in regions associated with AD pathology. This tracer would be useful for understanding early changes in AD pathology, when CB1 receptor density is supposedly increased.56,68 In addition, [11C]OMAR may be used to investigate fluctuations in the ES signaling at different stages of AD.
ES metabolism tracers (FAAH and MAGL)
Autoradiographic work has demonstrated that tracers targeting the main enzymes involved in eCB metabolism, FAAH and MAGL, have great potential to evaluate its activity in the brain. One of the first to be investigated was [11C]CURB that binds selectively, specifically, and irreversibly to FAAH, with a higher cortical uptake in a dose-dependent manner. 84 In studies using PET, [11C]CURB was defined as highly sensitive and selective for imaging of brain FAAH activity with an irreversible two-tissue compartment kinetic model.90,91 Further findings demonstrated that FAAH activity in the brain is dependent on a common gene variant, rs324420, and genotyping individuals during PET studies allowed for a more accurate interpretation of the data. 92
Among other ES metabolic tracer candidates proposed for PET, an irreversible inhibitor of FAAH, [18F]PF-9811, should also be mentioned. Studies in rodents have shown that this tracer exhibits a high specificity for the FAAH enzyme. Compared with CB1 and CB2 receptor tracers, however, it presents a lower binding affinity. 85 The metabolism of eCBs has also been explored using [11C]TZPU and [11C]SAR127303, both irreversible inhibitors of MAGL. 86 To date, [11C]SAR127303 is thought to be the most promising based on studies conducted in rats and monkeys that demonstrated high brain penetrance, specificity, and regional brain distribution. 86
Despite its lower affinity compared with other tracers, [11C]SAR127303 is a potential lead for clinical translation and may facilitate studies aimed at improving our understanding of the ES signaling in neurodegenerative diseases.
Due to the promising results observed in other neuropsychiatric disorders, PET tracers for ES metabolism could be used to better understand the ES signaling in AD. It is likely that studies targeting early stages of AD would present higher activity of enzymes FAAH and MAGL enzymes, as an attempt to compensate for the upregulation of CB receptors. These findings would also help to better understand the ES signaling in AD, as there is to date little information on the metabolism of the eCBs.
Fluid biomarkers
It is well established that AEA and 2-AG levels are altered in postmortem AD brain tissue (Supplementary Table S1). Koppel et al. investigated for the first time whether blood or cerebrospinal fluid (CSF) eCBs levels, measured by liquid chromatography/tandem mass spectrometry, could be used as fluid biomarkers in AD. 93 However, the authors observed similar levels of these markers in the blood of CU and AD patients. In a separate cohort composed of CU individuals at risk for AD, AEA was not detected in the CSF—possibly due to its very rapid metabolism—whereas 2-AG levels were not significantly correlated with any measures of cognition. 93
In contrast with these results, Altamura et al. demonstrated that 2-AG plasma concentrations were significantly higher in AD individuals compared with CU individuals and observed a positive correlation with memory and attention in AD individuals. 94 However, as in the first study, no significant difference in AEA levels was detected, even though studies using human brain tissue have shown changes in its metabolism.
In Hillard's review of circulating eCB, it has been highlighted that many variables might alter its measurement, including sex, stress, circadian rhythm, sleeping time, and food intake. 95 Thus, the collection procedure must follow a strict and clearly designed protocol to avoid study bias that might compromise reproducibility. In addition, the discrepancies observed between studies might be attributed to patient selection, as both studies used only clinical criteria for the diagnosis of AD (i.e., no biomarkers). In addition, it would be important to evaluate whether fluctuations of eCBs associate with microglia states present in the AD continuum. Clinical and biomarker evidence of AD should be taken into consideration in future studies investigating eCB fluid biomarkers in AD.
ES Biomarkers in the NIA-AA Research Framework
To biologically define AD, the 2018 NIA-AA Research Framework recommends characterizing individuals according to the AT(N) classification, using fluid and neuroimaging biomarkers. 4 Despite focusing on the core/classical AD biomarkers, the AT(N) is encouraging the incorporation of new biomarkers inside existing categories or even the creation of additional biomarker groups, the ATX(N). 12 Though additional research is needed, CB1 receptor density could be a surrogate marker of ES-linked neurodegeneration. By contrast, CB2 receptor is overexpressed in activated microglia and could be part of a new composite for defining immunophenotype (part of the “X” array of biomarkers).11,96 It is important to emphasize that ES biomarkers may serve to prognostic evaluation but not as AD-specific diagnostic tools.
Even though it is still unknown whether ES abnormalities are AD-specific, they may provide important information about biologically heterogeneous AD presentations. However, studies using fluid biomarkers presented mixed results, limiting their current inclusion in AD clinical studies. On the other hand, ES PET tracers, binding to CB receptors, seem to be the most promising ES candidates. More specifically, PET CB receptors ligands could be tested in individuals classified according to the AT(N) framework since they seem to reflect changes across AD stages (Fig. 2).
Longitudinal studies with biological-AD inclusion criteria and evaluation must be carried out to address whether the ES responds to the pre-amyloid phase and how it could be detected.
Finally, whether ES biomarkers are associated with cognitive changes across the AD continuum is an unanswered question. Their potential role for the prediction of conversion from asymptomatic to the symptomatic phase of AD should be evaluated.
ES Drugs Targeting AD
The ES biomarkers may serve as drug-target engagement tools, since multiple ES drugs targeting in AD are under development.66,97–99 Several in vitro and in vivo studies have demonstrated that certain CB compounds confer neuroprotective effects against Aβ accumulation. The evaluation of the impact of synthetic CB drugs using AD animal models is being investigated; of which the most used in AD studies are WIN 55,212-2 (a mixed CB1/CB2 receptor agonist), JWH analogs (a CB2 receptor agonist), and HU-308 (a CB1 receptor selective agonist). All these compounds present in common anti-inflammatory activity, preventing events associated to microglial activation (Supplementary Table S1).100,101
Clinical studies have been conducted to evaluate the effect of CBs drugs in AD. 102 However, no significant correlations between CBs and cognitive status have been reported so far in the few clinical studies investigating this topic. More studies, with a higher number of subjects, are needed for drawing conclusions. In addition, drugs targeting CB1 and CB2 receptors could take full advantage of CB tracer to therapeutic decision making.
Concluding Remarks and Future Perspectives
The ES is involved in the regulation of physiological brain functions, ranging from synaptic regulation to neuroinflammatory responses. Based on postmortem autoradiography studies in humans and rodents, it is thought that CB1 receptors are upregulated in the early stages of AD, possibly as a response to neuroinflammatory processes. By contrast, it was observed that CB2 receptors are positively correlated with high levels of Aβ load.
However, these findings have not been replicated in the two human PET studies conducted to date and further investigation is required. Currently, PET-based biomarkers appear more promising than CSF or blood-based measures for detecting alterations in the ES across the AD spectrum. Specifically, ES tracers were able to identify a relationship between AD-ES among different stages of the disease, whereas the only two studies using fluids biomarker report conflicting results. Further, the results concerning receptor density throughout AD progression remain controversial. One of the reasons may be that AD patients were diagnosed using clinical criteria only, without the use of biomarkers.
Another critical factor is that some studies have shown that PET tracers' uptake present significant sex differences that should be considered in future ES studies. Present clinical research on drugs targeting the ES in AD has not shown promising results. However, it is important to note that there are only a few studies with small sample sizes. Novel trials need to be implemented with the promising targets such as the CB1/CB2 receptor ligands.
As the ES is altered in AD, identifying biomarkers that reflect its functioning may help to characterize AD as a biological construct better, particularly if combined with established AT(N) biomarkers. Also, future longitudinal studies using subjects along the AD clinical continuum may help elucidate the role of the ES in AD and its dynamics across the trajectory of AD.
Footnotes
Authors' Contributions
All authors confirmed they have contributed to the intellectual content of this article and have met the following four requirements: (i) significant contributions to the conception and design, acquisition of data, or analysis and interpretation of data; (ii) drafting or revising the article for intellectual content; (iii) final approval of the published article; and (iv) agreement to be accountable for all aspects of the article, thus ensuring that questions related to the accuracy or integrity of any part of the article are appropriately investigated and resolved.
Author Disclosure Statement
E.R.Z. serves in the scientific advisory board of Next innovative Therapeutics (Nintx). A.L. serves on the scientific advisory board of Cerveau/Enigma Biomedical Group. The other authors declare no conflict of interest.
Funding Information
E.R.Z. receives financial support from CNPq (no. 435642/2018-9 and no. 312410/2018-2), Instituto Serrapilheira (no. Serra-1912-31365), Brazilian National Institute of Science and Technology in Excitotoxicity and Neuroprotection (no. 465671/2014-4), FAPERGS/MS/CNPq/SESRS-PPSUS (no. 30786.434.24734.231120170), and ARD/FAPERGS (no. 54392.632.30451.05032021) and Alzheimer's Association [AARGD-21-850670] and Alzheimer's Association and National Academy of Neuropsychology [ALZ-NAN-22-928381]. P.C.L.F. is supported by Alzheimer's Association (AARFD-22-923814). P.R.-N. receives financial support from Weston Brain Institute, Canadian Institutes of Health Research (no. MOP-11-51-31; RFN 152985, 159815, 162303), Canadian Consortium of Neurodegeneration and Aging (MOP-11-51-31—team 1), the Alzheimer's Association (no. NIRG-12-92090, no. NIRP-12-259245), Brain Canada Foundation (CFI Project 34874; 33397), and the Fonds de Recherche du Québec–Santé (Chercheur Boursier, no. 2020-VICO-279314). T.A.P. is supported by the NIH (no. R01AG075336 and no. R01AG073267) and the Alzheimer's Association (no. AACSF-20-648075).
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.