
Abstract
Background:
The underlying pathomechanism of acute lung injury (ALI)/acute respiratory distress syndrome (ARDS) is the immune response to inflammation or infection within the pulmonary microcirculation. Systemic spread of pathogens, activated immune cells, and inflammatory mediators contributes significantly to mortality in patients with ARDS.
Objective:
The endogenous cannabinoid system is a major modulator of the immune response during inflammation and infection. Phytocannabinoids, such as cannabidiol (CBD), have shown promising anti-inflammatory effects in several pathologies. The overall objective of this study was to evaluate the effects of CBD on local and systemic inflammation in endotoxin-induced ALI in mice.
Materials and Methods:
ALI was induced by pulmonary endotoxin challenge. Four groups of male C57BL/6 mice were randomized in this study: control, ALI, ALI with CBD treatment, and control with CBD treatment. Analyses of local and systemic cytokine levels, lung histology, and leukocyte activation as visualized by intravital microscopy of the intestinal and pulmonary microcirculation were performed 6 h following intranasal endotoxin administration.
Results:
Pulmonary endotoxin challenge induced significant inflammation evidenced by local and systemic cytokine and chemokine release, lung histopathology, and leukocyte adhesion. Intraperitoneal CBD treatment resulted in a significant decrease in systemic inflammation as shown by reduced leukocyte adhesion in the intestinal microcirculation and reduced plasma cytokine and chemokine levels. Pulmonary chemokine levels were decreased, while pulmonary cytokine levels were unchanged. Surprisingly, the ALI score was slightly increased by CBD treatment in a manner driven by enhanced neutrophil infiltration of the alveoli.
Conclusion:
In this model of experimental ALI, CBD administration was associated with reduced systemic inflammation and heterogeneous effects on pulmonary inflammation. Future studies should explore the mechanisms involved as they relate to neutrophil infiltration and proinflammatory mediator production within the lungs.
Introduction
With the recent COVID-19 pandemic, acute respiratory distress syndrome (ARDS) became temporarily the most frequent reason for admission to intensive care units worldwide.1,2 Before the arrival of SARS-CoV-2, however, ARDS already represented a major indication for intensive care owing to diverse etiologies, including pneumonia, sepsis, aspiration, trauma, and others.3,4 ARDS is categorized as mild, moderate, or severe based on degree of hypoxemia, and develops from a continuum of pathological pulmonary inflammation termed acute lung injury (ALI).5,6 This phenomenon can result in the spread of pathogens, activated leukocyte subpopulations, and proinflammatory cytokines into the systemic circulation, thereby contributing to systemic complications such as multiple organ dysfunction and death. 6
The endocannabinoid system (ECS) consists of endogenous ligands (e.g., anandamide and 2-arachidonoylglycerol), canonical G protein-coupled receptors (i.e., CB1 and CB2), and specific enzymes for ligand synthesis and degradation. 7 CB1 receptors are predominantly expressed in the brain, whereas CB2 receptors are usually upregulated on immune cells during inflammation. 8 CB2 receptor activation exerts anti-inflammatory actions, 9 and selective pharmacologic targeting of CB2 pathways appears to be a promising approach to reduce systemic inflammation in ALI. For instance, our group recently demonstrated that the synthetic CB2-selective agonist HU-308 reduces peripheral cytokine release and leukocyte adhesion in lipopolysaccharide (LPS)-induced ALI. 10
While the immunomodulatory potential of CB2 has been extensively demonstrated, anti-inflammatory effects may also be achieved via other pathways related to the ECS. A prime example of this is cannabidiol (CBD), a phytocannabinoid that has been shown to reduce inflammation via actions at multiple receptor targets. For example, in various studies, the anti-inflammatory effects of CBD have been linked to activation of 5-HT1A, 11 adenosine A2A, 12 or PPARγ, 13 inhibition of anandamide breakdown, 14 and direct or indirect antioxidant effects as reviewed by Atalay et al. 15 Regarding actions at the CB2 receptor, CBD has been reported as a partial agonist, 16 negative allosteric modulator, 17 and inverse agonist or antagonist 18 under different conditions.
Irrespective of molecular mechanism, CBD administration has shown anti-inflammatory effects in several pre-clinical and clinical inflammatory conditions, which may reflect the multimodal actions of this agent at distinct targets.19–21 Our goal was to study the utility of CBD in ALI-related local and systemic inflammation.
Materials and Methods
Experimental animals
Male C57BL/6 mice (wild type, 12–14 weeks old, 20–30 g body weight) were purchased from Charles River Laboratories International, Inc. (Saint-Constant, QC, Canada) and housed in ventilated plastic cages in a pathogen-free room of the Carleton Animal Care Facility (CACF), Dalhousie University. Animals were kept on a 12-h light/dark cycle at 21°C and acclimatized for a minimum of one week before experiments. All procedures were approved under protocol number 20-075 by the University Committee on Laboratory Animals at Dalhousie University and performed following the guidelines of the Canadian Council on Animal Care.
Experimental model and groups
Experimental groups included control (CON, n=18), LPS (n=15), LPS+CBD (n=15), CON+CBD (n=9). LPS from Pseudomonas aeruginosa (Sigma-Aldrich, Oakville, ON, Canada; L8643, source No. 12180104) was diluted to 10 mg/mL in sterile saline (0.9% sodium chloride [NaCl]) and stored at −20°C. CBD (Quantum Pharmaceuticals, Montreal, QC, Canada) was dissolved in anhydrous ethanol to 40 mg/mL and stored at −20°C. On the day of experiment, a 1:1:18 solution of CBD/ethanol:Kolliphor EL:sterile saline was prepared for injection. Mice were anesthetized via sodium pentobarbital (90 mg/kg intraperitoneal [i.p.]) or isoflurane gas (1–5%; pulmonary intravital microscopy [IVM] experiments only) and secured on a small animal intubation stand (Kent Scientific, Torrington, CT, USA). The tongue was gently protracted and small droplets of LPS solution (LPS groups) or saline (CON groups) were pipetted into the left nostril to a dosage of 5 mg/kg or equivalent volume saline.
Αn intraperitoneal CBD dosage of 10 mg/kg was selected based on previous studies.13,21 CBD (CBD groups) or vehicle (non-CBD groups) was administered immediately after intranasal instillation, following which mice were returned to individual cages on heating pads for hourly monitoring. Six hours after injection, 5–8 animals per group were sacrificed for collection of blood and lung tissues. This time point was selected on the basis of a previous study by our group demonstrating an elevated systemic proinflammatory cytokine release at 6 h postinduction. 10 An additional 4–5 animals per group were anesthetized for intestinal or pulmonary IVM (see the “Intestinal IVM” and “Pulmonary IVM” sections).
Multiplex assays
Whole blood samples were taken by cardiac puncture and centrifuged at 3000 rpm at 4°C for 10 min. Plasma was extracted and stored at −80°C. For cytokine and chemokine analysis of lung tissue, a portion of left lung was removed, flash-frozen in liquid nitrogen, and stored at −80°C. Lung tissue was homogenized with T-PER buffer using a bead mill homogenizer (Bead Ruptor Elite; Omni International, Kennesaw, GA, USA) and total protein concentration of lysate was evaluated by bicinchoninic acid assay (Pierce BCA Protein Assay Kit; Thermo Fisher Scientific, Waltham, MA, USA).
Plasma and lung tissue lysate were analyzed using magnetic bead-based multiplex assays obtained from R&D Systems (Minneapolis, MN, USA). Multiplexed analytes for plasma samples consisted of interleukin 6 (IL-6), tumor necrosis factor-α (TNF-α), chemokine (C-X-C motif) ligands 1 and 2 (CXCL1, CXCL2), interferon-γ (IFN-γ), intercellular adhesion molecule 1, interleukin-10 (IL-10), interleukin-1β (IL-1β), lipopolysaccharide-induced CXC chemokine (LIX), and p-selectin. Multiplexed analytes for lung tissue lysate samples consisted of IL-6, TNF-α, CXCL1, CXCL2, IFN-γ, IL-1β, and LIX. Samples were processed as described by the manufacturer and assayed in duplicate with a sample dilution of 1:2 for plasma and 1:5 for lung tissue. Plates were read using the Bio-Rad 200 luminometer with Bio-Plex manager software (Bio-Rad, Mississauga, ON, Canada). Lung tissue sample values were normalized to total protein concentration.
Lung histology
Right lungs were fixed in 10% phosphate-buffered formalin for 3 days. Samples were cleaned to remove connective and muscle tissue and stored in 70% ethanol before embedding in paraffin. Embedded tissues were cut into 5 μm sections, mounted, and dried for a minimum of 2 days in an oven (56–76°C), and then stored at room temperature before hematoxylin and eosin staining. Finally, stained tissue sections were examined via brightfield microscopy (Optika B-290TB; Optika SRL, Ponteranica, BG, Italy) and imaging software (Optika Vision Lite 2.1).
A previously described scoring system was used to semiquantitatively analyze lung injury in tissue sections. 22 In brief, 10 high-powered fields of view per slide were selected and each was scored in a blinded manner on the basis of neutrophil presence in the alveolar space (NA), neutrophil presence in the interstitium (NI), hyaline membrane formation (HM), protein debris in the alveolar space (PD), and alveolar septal thickening (ST). For neutrophil presence, scores of 0, 1, or 2 were given for the presence of n=0, 1≤n≤5, or n>5 neutrophils, respectively. For hyaline membrane and protein debris, scores of 0, 1, or 2 were given for the presence of n=0, n=1, or n>1 instances, respectively. For septal thickening, scores of 0, 1, or 2 were given for n<2x, 2x ≤ n≤4x, n>4x, respectively.
From these five numerical scores, a composite score was generated using the formula “ALI Score=[0.2(NA) +0.14(NI) +0.07(HM) +0.07(PD) +0.02(ST)]/(number of fields).”
Intestinal IVM
Animals were anesthetized via sodium pentobarbital (90 mg/kg; Ceva Sainte Animale, Montreal, QC, Canada). Fifteen minutes before imaging, rhodamine-6G (0.05%, 1.5 mL/kg; Sigma-Aldrich) and fluorescein isothiocyanate-labeled bovine serum albumin (FITC-BSA, 5%, 1 mL/kg; Sigma-Aldrich) were administered via tail vein. A laparotomy was performed to expose the terminal ileum. The mouse was placed on its side with a loop of the terminal ileum placed on the viewing platform of a specifically designed stage fixed to a heating pad. 23 A glass slide was placed over the terminal ileum and a continuous flow of 0.9% NaCl (heated to 37°C, 7 mL/h) was applied to the region to approximate physiological conditions. An epifluorescence microscope (Leica DMLM, Wetzlar, Germany) with a mercury-arc light source (LEJ EBQ 100; Carl Zeiss, Jena, Germany) and 530–550 nm bandpass excitation filter enabled visualization of the intestinal microcirculation.
Leukocyte trafficking was visualized within both submucosal collecting venules (V1, diameter=50–100 μm) and postcapillary venules (V3, diameter=20–35 μm). For each venule type, six videos of 30 sec in length were acquired. Videos were analyzed in a blinded manual manner using Fiji. 24 Adherent leukocytes were defined as labeled cells that remained immobile for the 30-sec recording period. Endothelial surface area was estimated under the assumption of cylindrical vessel geometry. Leukocyte adhesion was quantified and reported as cells/mm2.
Pulmonary IVM
Pulmonary microcirculation was imaged in vivo at 6 h postinduction in a previously described manner. 25 Briefly, animals were anesthetized with isoflurane 1–5% (Fresenius Kabi, Bad Homburg, Germany), intubated, and mechanically ventilated. Body temperature was maintained at 37.0±0.5°C. Rhodamine-6G and FITC-BSA were applied as in intestinal IVM methods. A thoracotomy was performed to expose the left lung, and a vacuum-stabilized imaging window (Luxidea, Calgary, Canada) was applied to stabilize the lung for imaging. Imaging was performed using an epifluorescence microscope as in intestinal IVM methods. Five fields of view each of pulmonary venules, arterioles, and capillary regions of interest (ROIs) were recorded for 30 sec using a black/white CCD camera. Videos were analyzed in a blinded manual manner using Fiji.
The area of capillary ROI was estimated under the assumption of planar geometry. Adherent leukocytes were defined as labeled cells that remained immobile for the 30-sec recording period. Leukocyte adhesion was quantified and reported as cells/mm2.
Statistical analysis
All data were analyzed using the statistical software package Prism 9 (GraphPad Software, La Jolla, CA, USA). Shapiro–Wilk test was used to confirm normal distribution of data. One-way ANOVA followed by Holm–Šídák's multiple comparison test was used to analyze normally distributed data. Comparisons were assessed between the LPS group and each other group, where applicable. Unpaired t tests were used to compare means where only two groups were concerned. All data were expressed as mean±standard deviation. Significance was assumed at p≤0.05 and denoted by asterisks as *p≤0.05, **p≤0.01, ***p≤0.001, and ****p≤0.0001.
Results
Multiplex assays
To assess the impact of CBD on LPS-induced inflammation, multiplex analysis of plasma and lung tissue lysate focused on four protein mediators that drive inflammation in ALI and other pathologies: the cytokines IL-6 and TNF-α, and CXCL1 and CXCL2. As per the methods, additional analytes were included in these assays and are reported in Supplementary Tables S1 and S2.
As expected, LPS challenge induced robust elevation of plasma IL-6, CXCL1, and CXCL2 (Fig. 1). TNF-α followed a similar trend, however, seven of eight plasma samples in the control group fell below the assay's detection threshold, and thus, a statistical comparison was not made between control and LPS. CBD treatment in LPS-challenged mice significantly reduced the release of CXCL2, IL-6, and TNF-α. Also, as predicted, CBD-treated controls exhibited low cytokine and chemokine levels versus LPS-challenged mice.
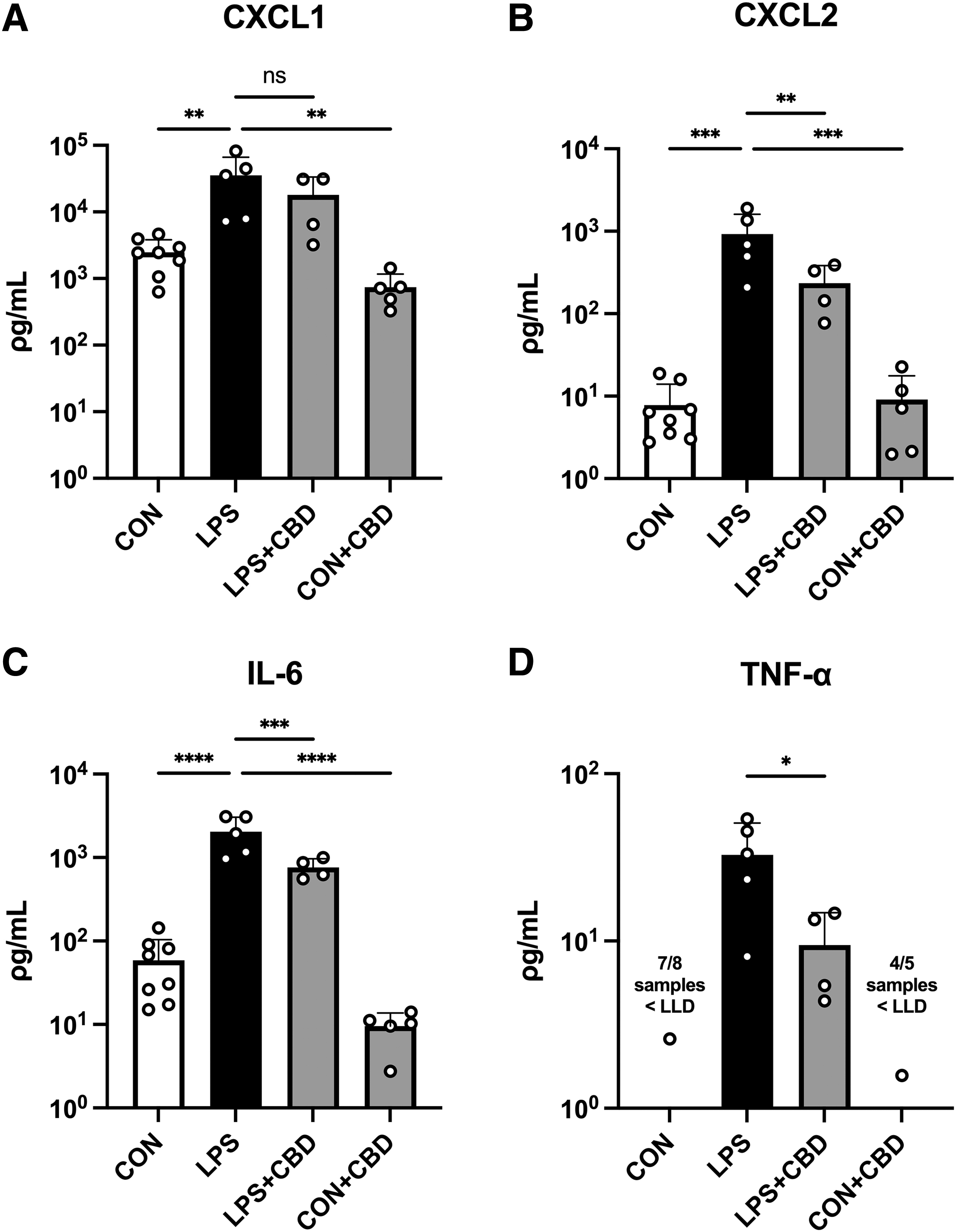
Plasma cytokine and chemokine levels measured by multiplex analysis.
In lung tissue lysate, we observed similar increases in cytokine and chemokine expression in LPS-challenged mice (Fig. 2). CBD treatment significantly reduced CXCL1 and CXCL2 release but did not significantly reduce the release of IL-6 or TNF-α. Also, as predicted, CBD-treated controls demonstrated low cytokine/chemokine release versus LPS-challenged mice, although no statistical comparison was performed for TNF-α due to detection threshold limitations.
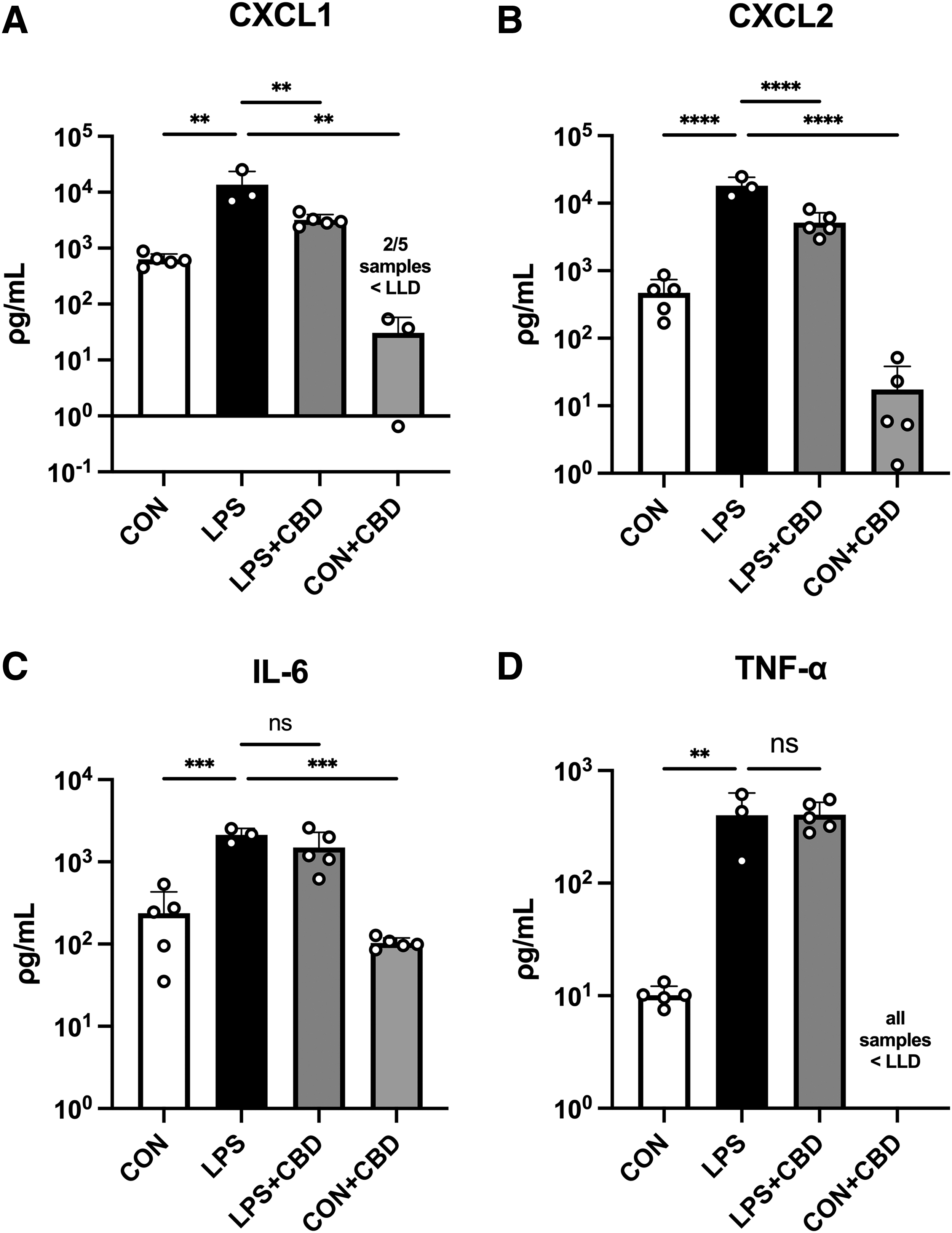
Lung tissue lysate cytokine and chemokine levels measured by multiplex analysis.
Lung histology
To assess changes in lung morphology and cell-mediated immune response, we performed a histological study of right lung samples. As predicted, LPS challenge caused a significant increase in ALI score (Fig. 3), although hyaline membranes were not observed. Surprisingly, CBD treatment modestly increased ALI score in LPS-challenged mice, a result that stood in contrast to the observed reductions in chemokine release in lung tissue. Further analysis of individual components of the composite ALI scores suggested that this increase was driven by increased alveolar neutrophilia (Supplementary Fig. S1). CBD-treated controls, however, did not display elevation in ALI score.
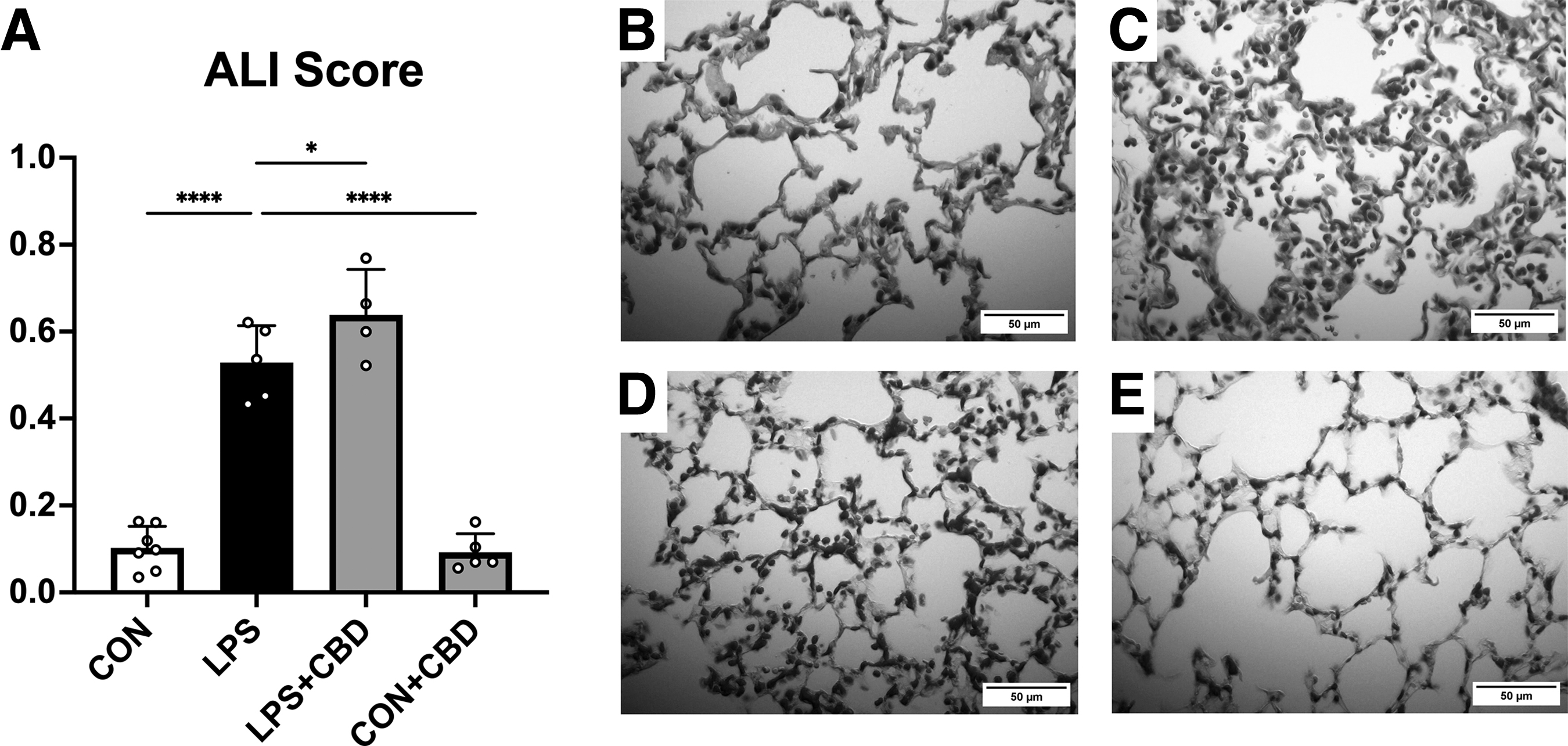
Histological analysis of hematoxylin and eosin-stained right lung sections.
Intravital microscopy
Firm adhesion to the endothelium is a key step in leukocyte transmigration and occurs at greater rates under inflammatory conditions. To assess occurrence of this phenomenon in the peripheral microcirculation, we performed IVM of collecting (V1) and postcapillary (V3) venules in the terminal ileum. Leukocyte adhesion was increased in both venule types in LPS-challenged mice (Fig. 4). CBD administration effectively attenuated this increase in LPS-challenged mice. As expected, CBD-treated controls exhibited low levels of leukocyte adhesion.
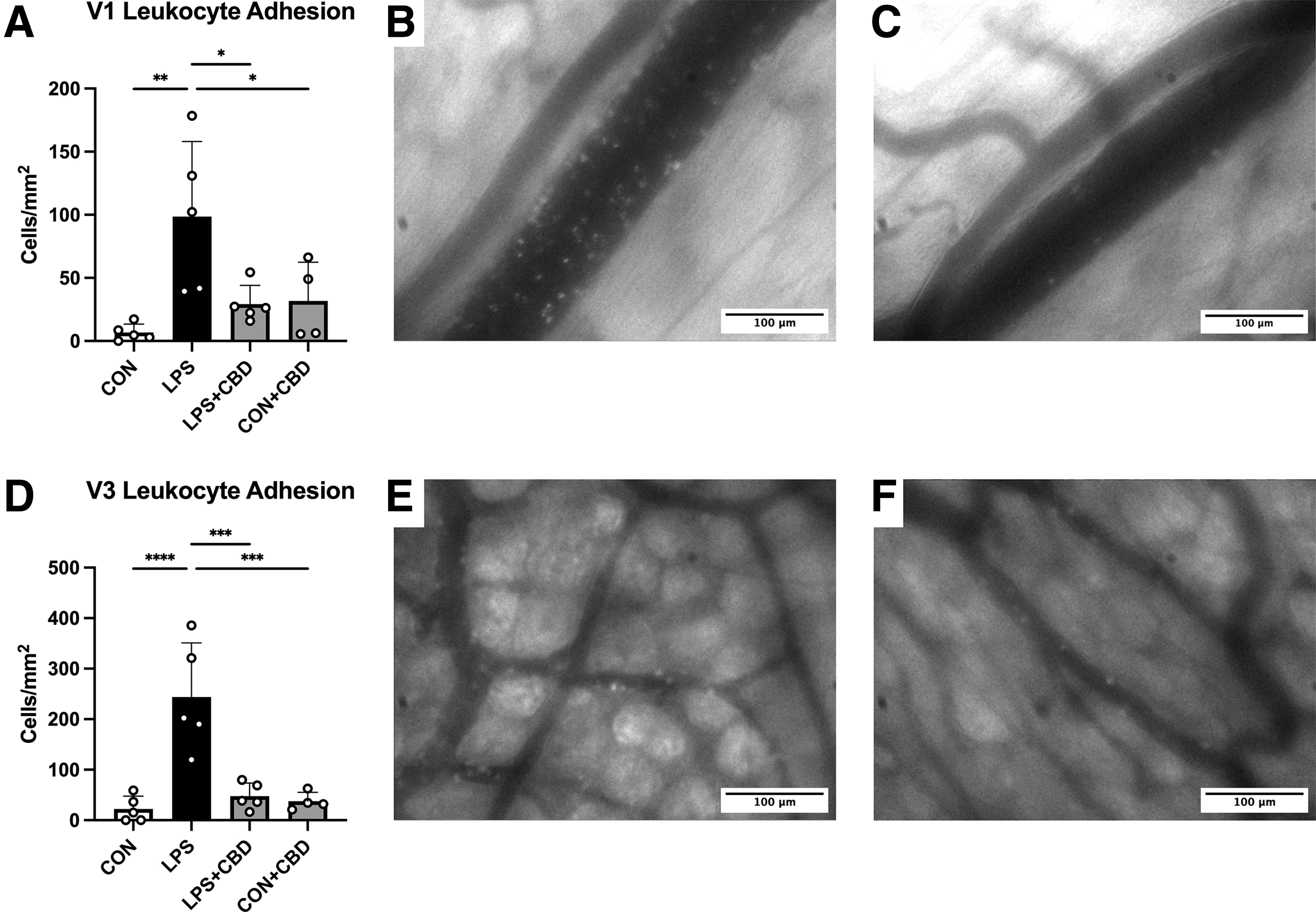
Intravital microscopy of terminal ileal venules.
We then used IVM to assess leukocyte adhesion in the pulmonary microcirculation. LPS challenge increased leukocyte adhesion in pulmonary capillary ROI (Fig. 5). This effect was not significantly reduced by CBD administration. Also of note, neither LPS challenge nor CBD administration significantly impacted leukocyte adhesion in pulmonary arterioles or venules (not shown). For practical reasons, the CBD-treated control group was not included in the pulmonary IVM study.

Intravital microscopy of pulmonary capillaries.
Discussion
We evaluated the effects of CBD treatment on local and systemic inflammation in an experimental model of ALI. The presented data demonstrate a reduction in systemic inflammation and pulmonary chemokine release, as well as an unexpected increase in alveolar neutrophil infiltration, in CBD-treated mice.
Systemic spread of inflammatory mediators represents the initiating step when local inflammation turns into a critical whole-body medical condition. In the case of an infectious origin of inflammation, this condition is defined as sepsis and the systemic cytokine release is often called “cytokine storm.” 26 Significant systemic cytokine release is also seen in other inflammatory conditions, such as a consequence of chimeric antigen receptor T-cell therapy, and is referred to as “cytokine release syndrome.” 27 Regardless of the underlying disorder, systemic cytokine release is a potentially life-threatening condition and an important contributor to morbidity and mortality in critical illness. Therefore, our findings in an experimental model of lung inflammation (i.e., pneumonia) are potentially important for the clinical outcome of ALI.
We observed that LPS challenge significantly elevated plasma and pulmonary tissue levels of proinflammatory cytokines IL-6 and TNF-α, as well as chemokines CXCL1 and CXCL2. CBD treatment appeared to reduce these changes, except for pulmonary tissue cytokine levels. A possible explanation for this exception may be a relatively low dose of CBD. For example, results from a study by Ribeiro et al. are consistent with a CBD dose-dependent reduction in IL-6 and TNF-α levels, but not macrophage inflammatory protein 2 (MIP-2; CXCL2) levels, in bronchoalveolar lavage fluid (BALF). 28 This was achieved at higher doses (20 and 80 mg/kg), whereas a 10 mg/kg dosage (our study) may not have been sufficient to reduce levels of IL-6 and TNF-α.
By contrast, lower doses of CBD have been shown to reduce inflammatory cytokine levels in lung tissue. Khodadadi et al. observed downregulation of pulmonary IL-6 and TNF-α, as well as pulmonary immune cell infiltration, with three daily treatments of 5 mg/kg CBD in an 8-day model of polyinosinic:polycytidylic acid (poly(I:C))-induced ALI. 29 While the reason for these inconsistencies with our results is unclear, it may be related to differences in experimental models. Toll-like receptor (TLR)4-selective agonists such as LPS have been shown to produce comparatively stronger stimulation of cytokine release versus poly(I:C) (TLR3 agonist). 30 In addition, the 8-day time line used by the authors differs substantially from the hyperacute time line used in our experiments. In line with this, plasma IL-6 levels in the poly(I:C) model of Khodadadi et al. were apparently lower than in our model, although levels were reduced by CBD treatment in both cases.
The histological findings herein may suggest an intact local immune response. Surprisingly, CBD treatment increased ALI score in LPS-challenged mice in a modest but statistically significant manner. This finding appeared to be driven by enhanced alveolar neutrophilia coinciding with decreased protein deposition and septal thickening (Supplementary Fig. S1), which represent 40%, 14%, and 4% of the composite ALI score, respectively. 22 This increase in neutrophil infiltration contradicts observed reductions in pulmonary CXCL1 and CXCL2 following CBD treatment, although other chemokines may also play a role here.
The enhanced neutrophilia in our model also contrasts with many studies reporting on the anti-inflammatory effects of CBD. Some of these reports include protection against poly(I:C)-induced lung histopathology 31 and reductions in LPS-induced pulmonary leukocyte recruitment. 28 Nonetheless, a similar finding to ours has been previously reported. Karmaus et al. observed that CBD modestly increased the number of inflammatory cells in BALF at 6 and 24 h post-LPS challenge. 32 The differences among these results may be related to differences in disease models, experimental time lines, CBD dosages, and/or routes of administration.
We used intestinal IVM to study the systemic immune response and observed a significant reduction of LPS-induced leukocyte activation following CBD treatment. Leukocyte adhesion in the gut microvasculature following intranasal LPS challenge was comparable with studies using i.p. or intravenous LPS injections.33,34 This is surprising and might indicate a significant absorption of LPS through the lungs into the systemic circulation. Alternatively, release of locally produced cytokines from the lungs may be the primary contributor to the peripheral immune response.
Another potential option is the direct activation of monocytes and neutrophils in the pulmonary microcirculation during acute inflammation and spreading of those primed immune cells into the systemic circulation. If activated neutrophils and monocytes come into contact with activated (cytokine-induced) endothelial cells in the peripheral (micro-) circulation, they possess the capacity to adhere, transmigrate, and contribute to remote organ damage—a typical event in sepsis causing multiorgan failure. 35 Therefore, observed reductions in leukocyte adhesion within the intestinal microcirculation might be important for the outcome in ALI-induced systemic inflammation.
IVM of the pulmonary microcirculation revealed no major changes in leukocyte adhesion within the pulmonary microcirculation following CBD treatment. This may be consistent with the histological observation that CBD treatment did not reduce neutrophil infiltration (based on ALI score), with the previously mentioned caveat that ALI score was increased by CBD treatment. However, LPS-induced increases in leukocyte adhesion were only seen in lung capillaries but not in arterioles or venules. This may suggest a relatively mild lung injury, which can be explained by the LPS dose. In addition, the shallow penetration depth associated with the observation area may have limited overlap with airspaces exposed to LPS. 10
It is not exactly known at which dose or time point an anti-inflammatory therapy impacts the physiologic immune response, but we do see a potential risk for antibacterial/antiviral defense with high-dose anti-inflammatory therapies. A higher CBD dose might have had stronger effects in our model—not only on the systemic inflammation but also on the local immune response as shown in the studies by the Khodadadi and Ribeiro et al.28,29 Data from glucocorticoid trials in COVID-19 patients with ALI suggest a higher rate of secondary bacterial infections, clearly showing the limits of immune-modulatory therapies. 36 Therefore, future studies should examine the effects of anti-inflammatory therapies such as CBD in the context of infection.
Limitations of our studies include the experimental time line, sex and age of the experimental animals, and methodological challenges. We treated the animals with a single dose of CBD at the beginning of the experiments. Successful later treatments with multiple time points are required to confirm our findings. Age and sex play an immutable role in inflammation in both humans and animals, and thus, further studies should include females and different age groups. 37 Finally, as mentioned previously, fluorescence IVM is only capable to study superficial tissue layers. Multiphoton microscopy has also been deployed in microcirculatory research and might offer “deeper” insights into the immune response within solid organs such as the lung.
Conclusions
The present study aimed to explore the therapeutic potential of CBD in ALI. The data herein suggest that CBD treatment could reduce systemic inflammation associated with the condition. However, the observed effects on inflammation within the lung compartment were varied and further studies are required to mechanistically investigate CBD's impacts on neutrophil recruitment and proinflammatory mediator production within the lungs. Finally, care should be taken when extrapolating data from laboratory animals to humans.
Footnotes
Acknowledgments
The authors would like to thank Bithika Ray and Tanya Myers for technical support. S.H. (PhD student) would like to acknowledge funding through a Student Research Scholarship from the Anesthesia Research Fund, Department of Anesthesia, Pain Management and Perioperative Medicine, and the I3V Wave, Dalhousie University, Halifax, Nova Scotia, Canada.
Authors' Contributions
S.H.: Methodology, formal analysis, investigation, writing (original draft), writing (review and editing), and visualization; S.F.: formal analysis and writing (review and editing); P.T.: investigation; M.C.: methodology; M.K.: funding acquisition; J.Z.: conceptualization, methodology, resources, supervision, funding acquisition, and article editing; C.L.: conceptualization, methodology, resources, writing (original draft), writing (review and editing), supervision, project administration, and funding acquisition.
Author Disclosure Statement
The authors have no conflicts of interest to disclose.
Funding Information
Financial support for this work was provided, in part, through an unrestricted grant from Tetra Bio-Pharma, Inc., Canada, and through the Legacy Research Fund (2019) from the Lung Association of Nova Scotia, Canada.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.